Пищеварение и обмен веществ | Tervisliku toitumise informatsioon
Съеденная пища должна перевариться, чтобы содержащиеся в ней питательные вещества всосались в кровь. Пищеварение осуществляет пищеварительная система человека, или пищеварительный аппарат. Пищеварительный аппарат состоит из ротовой полости, глотки, пищевода, желудка, тонкой кишки (в т.ч. двенадцатиперстной кишки, тощей кишки, подвздошной кишки) и толстой кишки. Также пищеварению способствуют поджелудочная железа (панкреас) и печень.
Желудочно-кишечный тракт, или пищеварительный канал, – трубчатый. Для обеспечения достаточно быстрой скорости всасывания всасывающая поверхность имеет разветвленную структуру. Особенно разветвленной является тонкая кишка. Между разветвлениями имеются пищеварительные железы, которые направляют пищеварительные соки в желудочно-кишечный тракт.
Внутренняя поверхность желудочно-кишечного тракта покрыта слизью, особенно много слизи в районе желудка и ниже.
Наличие слизи необходимо по трем причинам:
- защищает от вредных факторов
- способствует продвижению перевариваемой массы
- в области кишечника в слизи содержится целый ряд исключительно необходимых пищеварительных ферментов и большая часть полезных микроорганизмов
Поскольку пищеварение и всасывание питательных веществ – это взаимосвязанные процессы, в клетках слизистой оболочки желудочно-кишечного тракта очень хорошее кровоснабжение. В желудочно-кишечном тракте перевариваемая масса движется дальше при помощи ритмичных сокращений слизистой оболочки желудка и кишечника, этот процесс и называется перистальтикой.
В желудочно-кишечном тракте перевариваемая масса движется дальше при помощи ритмичных сокращений слизистой оболочки желудка и кишечника, этот процесс и называется перистальтикой.
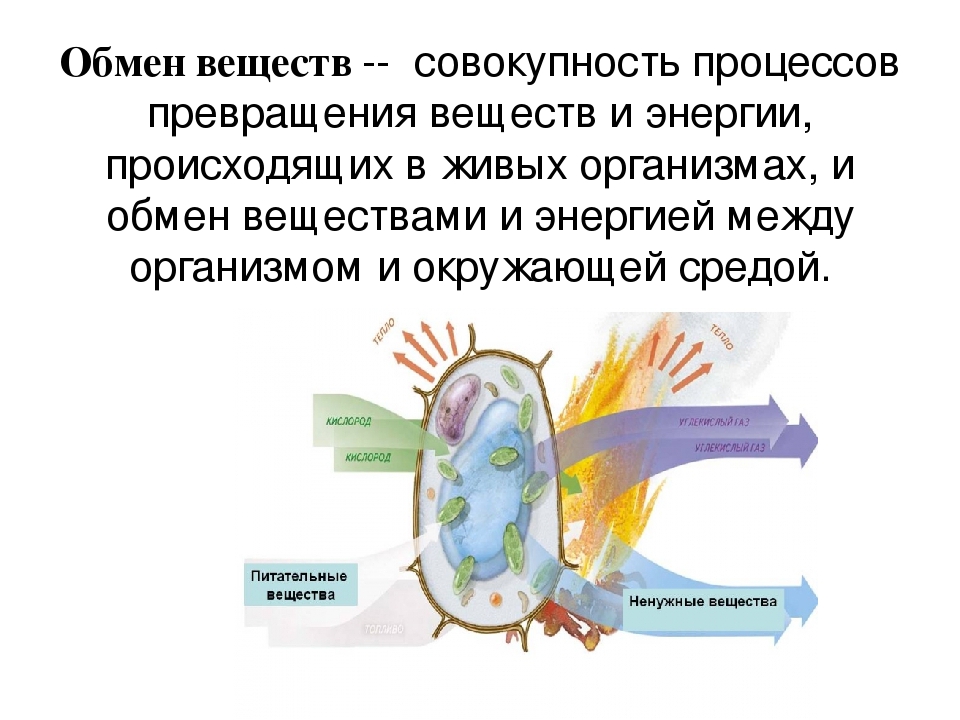
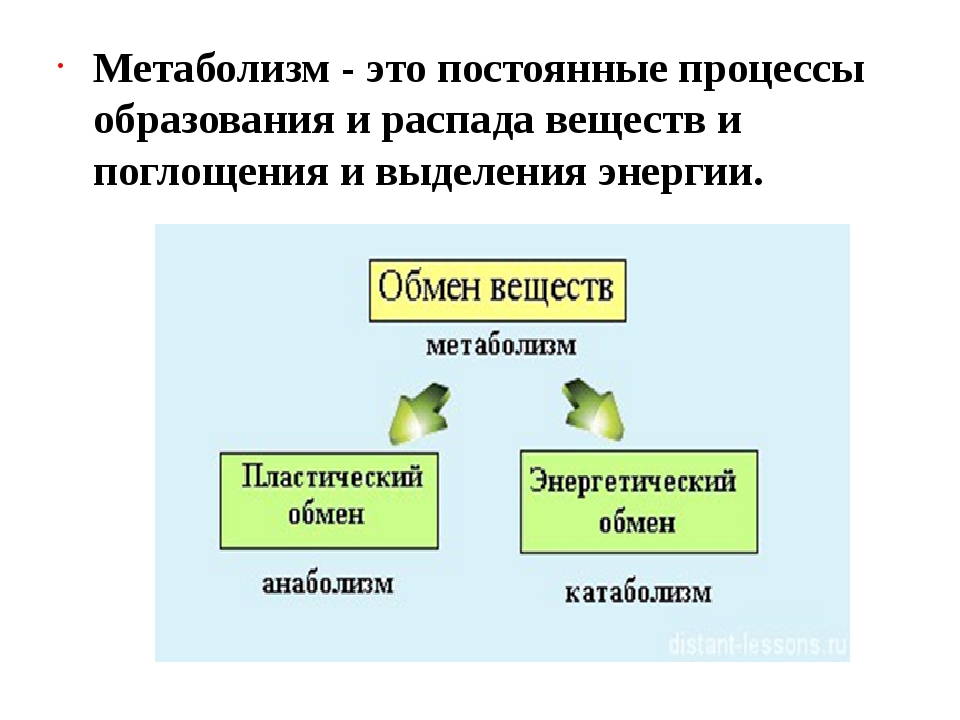
Обмен веществ, или метаболизм, – это совокупность всех (ферментных) реакций, которые происходят в клетке.
Обмен веществ является основой жизнедеятельности организма. Обмен веществ в организме человека – это крайне сложный процесс, в котором принимает участие около 30000 белков, 4000 из которых являются ферментами. Условно обмен веществ можно разделить на катаболизм и анаболизм (процессы расщепления и синтеза).
Основные функции обмена веществ:
- расщепление питательных веществ, их всасывание (переваривание) и использование,
- посредством синтеза биомолекул тела, которые являются строительным материалом,
- для производства энергии,
- вывод из организма конечных продуктов обмена веществ, обезвреживание и вывод из организма чужеродных соединений.
Основные процессы обмена веществ одинаковы у всех людей! Поскольку скорость работы (активность) различных ферментов у разных людей не всегда абсолютно одинакова, скорость обмена веществ также может различаться.
Страницы о пищеварении и обмене веществ были подготовлены совместно с Михкелем Зильмером, профессором медицинской биохимии Тартуского университета.
Обмен веществ и энергии — Сонин 6 класс (ответы)
104. Дайте определение
Обмен веществ — это получение организмом из внешней среды нужных веществ и удаление из организма во внешнюю среду ненужных
105. Каково значение обмена веществ для живого организма?
Без него невозможен рост и обновление клеток любого организма
106. Какие процессы происходят в хлоропластах и митохондриях клетки?
В хлоропластах — синтез органических веществ из неорганических на свету, при использовании воды и углекислого газа. Побочный продукт — кислород
В митохондриях происходит распад органических веществ и синтез энергии
107. Заполните схему «Обмен веществ у животных»
Заполните схему «Обмен веществ у животных»
Вещества, поступающие в организм Процессы, происходящие при обмене веществ Вещества, образовавшиеся в результате обмена веществ кислород, белки, жиры, углеводы, минеральные соли процессы жизнедеятельности вода, углекислый газ, продукты распада
108. Заполните схему «Обмен веществ у растений»
Вещества, поступающие в организм Процессы, происходящие при обмене веществ Вещества, образовавшиеся в результате обмена веществ кислород, углекислый газ, свет, вода с растворенными минеральными веществами дыхание, фотосинтез углекислый газ, кислород, пары воды
109. Дайте определения
Теплокровные животные — это животные, температура тела которых зависит от температуры окружающей среды
Холоднокровные животные — это животные, температура тела которых зависит от температуры окружающей среды
110. Приведите примеры животных, которые относятся к этим группам
Приведите примеры животных, которые относятся к этим группам
Теплокровные: лев, человек, волк
Холоднокровные: окунь, лягушка, черепаха
Обмен веществ и энергии
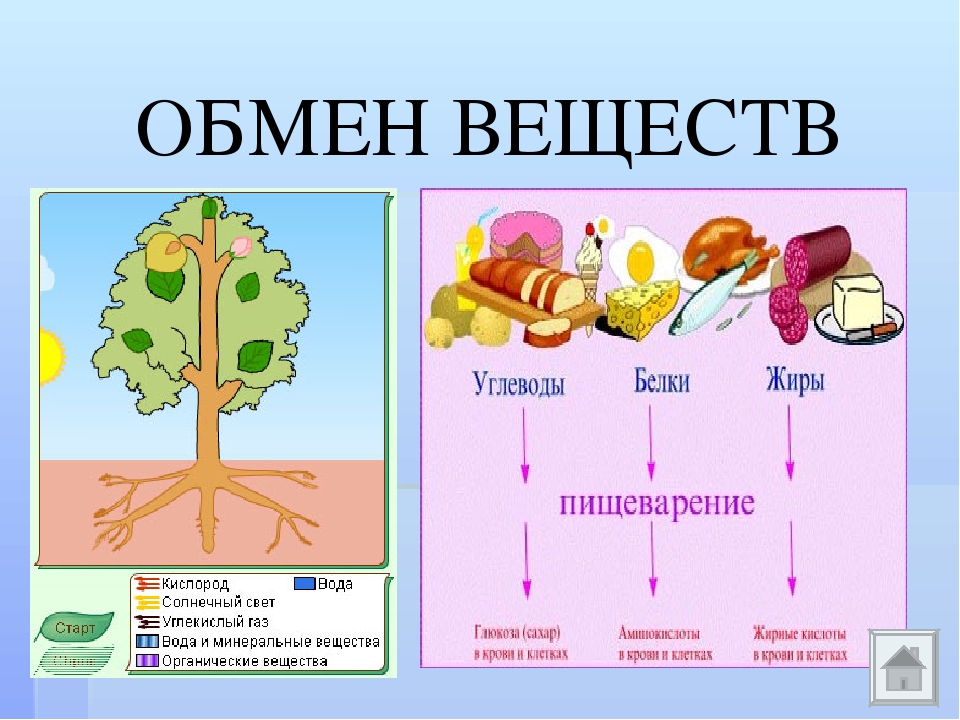
Обмен веществ — это совокупность протекающих в живых организмах химических превращений, обеспечивающих их рост, развитие, процессы жизнедеятельности, воспроизведение потомства, активное взаимодействие с окружающей средой. Во всех живых организмах, от самых примитивных до самых сложных, каким является человек, основа жизни — это обмен веществ и энергии. Благодаря ему каждый организм не только поддерживает своё существование, но развивается и растет. Обмен веществ определяет цикличность жизни: рождение, рост и развитие, старение и смерть.
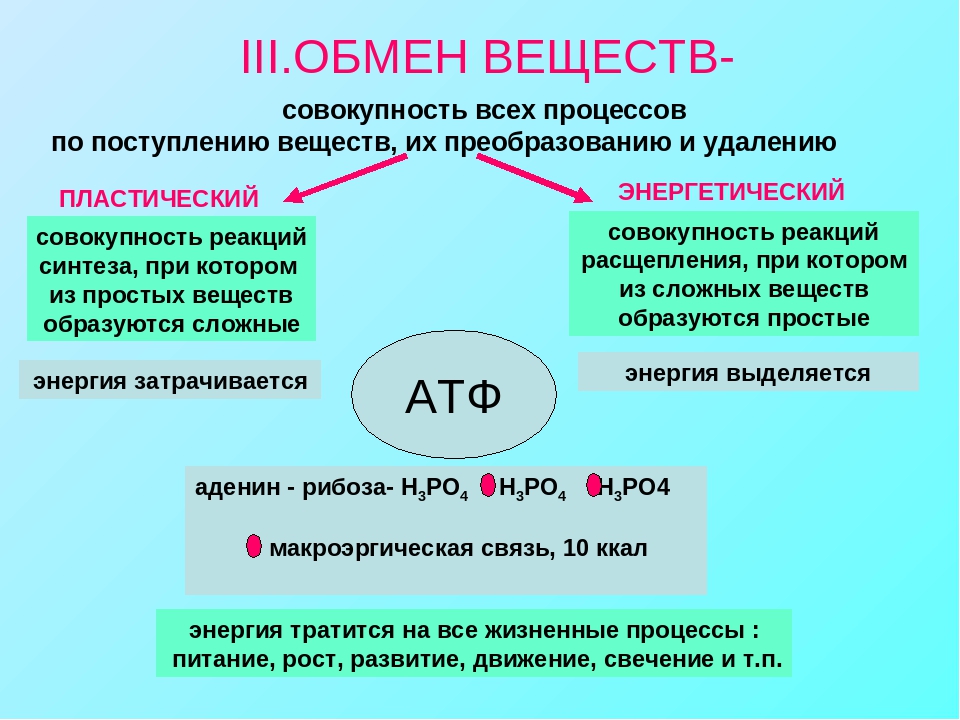
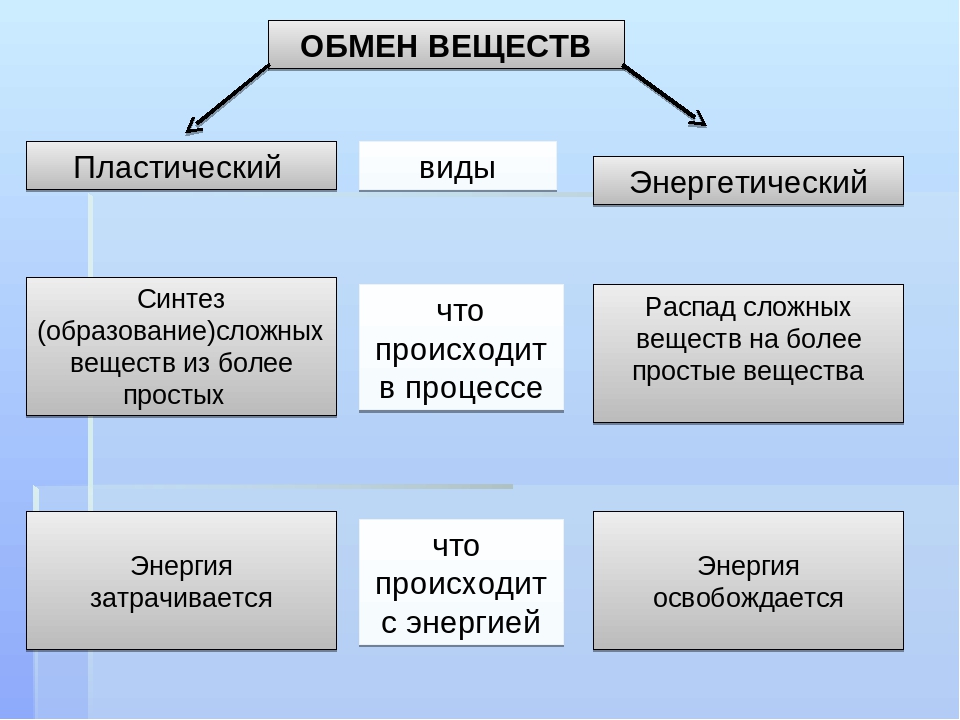
Пластический и энергетический обмен
Под пластическим обменом
 Под энергетическим обменом понимают такие превращения энергии, в ходе которых в результате биологического окисления выделяется энергия, необходимая для жизнедеятельности клеток, тканей и всего организма в целом. Результатом биологического окисления является образование углекислого газа, аммиака, соединений фосфора, натрия, хлора, которые выводятся из организма. Эта заключительная стадия обмена веществ. Она осуществляется кровью, легкими, потовыми железами, органами мочевыделения.
Под энергетическим обменом понимают такие превращения энергии, в ходе которых в результате биологического окисления выделяется энергия, необходимая для жизнедеятельности клеток, тканей и всего организма в целом. Результатом биологического окисления является образование углекислого газа, аммиака, соединений фосфора, натрия, хлора, которые выводятся из организма. Эта заключительная стадия обмена веществ. Она осуществляется кровью, легкими, потовыми железами, органами мочевыделения.Обмен белков
Пищевые белки в ходе подготовительной стадии обмена расщепляются сначала в желудке пепсином, а затем в двенадцатиперстной кишке ферментом поджелудочной железы трипепсином до аминокислот. Аминокислоты через кровеносные капилляры ворсинок поступают в печень. Здесь избыточные аминокислоты теряют свой азот и превращают в жиры и углеводы. В клетках из аминокислот строятся белки тела. Белки входят в состав ядер, цитоплазмы и мембран клеток. Они являются ферментами, входят в состав антител.
Обмен жиров
В органах пищеварения во время подготовительной фазы обмена жиры распадаются на глицерин и жирные кислоты. В эпителии кишечника синтезируется жир, характерный для организма, и через лимфатическую систему направляется в жировое депо и клетки, где он используется как запасное вещество и строительный материал. Жиры выполняют в организме много функций. Они входят в состав клеточных мембран, в них растворяются некоторые витамины. Из жиров образуются некоторые гормоны и биологически активные вещества. В организме человека выполняют защитную роль.
Обмен углеводов
Сложные углеводы начинают распадаться в ротовой полости под действием ферментов слюны — амилазы. В двенадцатиперстной кишке под действием ферментов, выделяемых поджелудочной железой, они расщепляются до глюкозы и других простых углеводов. В тонкой кишке продукты распада всасываются кишечными ворсинками в кровь и направляются в печень. Здесь излишки сахаров задерживаются и превращаются в гликоген и другие соединения, а оставшаяся часть глюкозы в необходимом количестве направляется в кровь и распределяется между клетками тела. В организме глюкоза прежде всего является источником энергии.
Здесь излишки сахаров задерживаются и превращаются в гликоген и другие соединения, а оставшаяся часть глюкозы в необходимом количестве направляется в кровь и распределяется между клетками тела. В организме глюкоза прежде всего является источником энергии.
Обмен воды в организме
Вода — универсальный растворитель. Все жизненные процессы, все биохимические реакции происходят в водной среде. Внутренняя среда человека содержит до 90% воды. Вода в организме либо химически связана с другими соединениями, либо содержит в себе растворенные минеральные соли и органические вещества. Пищеварительные соки содержат воду. Транспорт питательных веществ и кислорода осуществляется в жидкой среде. Продукты распада тоже выносятся водой. Таким образом, в организме поддерживается определенный баланс между поступающей и выделяемой водой.
Обмен минеральных солей
Ни вода, ни минеральные соли не являются источниками энергии, но они необходимы для осуществления важных функций организма. Минеральные соли содержатся в клеточных ядрах и цитоплазме, в жидкостях, образующих внутреннюю среду, в пищеварительных соках и других биологических жидкостях.
Минеральные соли содержатся в клеточных ядрах и цитоплазме, в жидкостях, образующих внутреннюю среду, в пищеварительных соках и других биологических жидкостях.
Другие заметки по биологии
Технологическая карта урока «Обмен веществ и энергии»
Тема «Обмен веществ и энергии» биология 6 класс (УМК Захаров, Сонин).
Цель урока: В ходе самостоятельной работы с текстом учебника, с помощью инструктивных листов, опорных рисунков, консультаций учителя и ранее приобретённых знаний изучить особенности обмена веществ у растений и животных, доказать, что обмен веществ и энергии – основное свойство живых организмов.
Ход урока:
1. Определение темы и цели урока. Мотивация и форма урока.
2. Инструктаж.
3. Самостоятельная работа. Консультация учителя.
Самостоятельная работа. Консультация учителя.
4. Закрепление и обсуждение полученных знаний.
5. Первичное применение знаний.
Оборудование: Учебник Сонин Н.И. Биология. Живой организм. 6 класс. Москва. Дрофа.2013, опорные конспекты-рисунки мультимедийная презентация (далее ММП) «Обмен веществ и энергии у растений», ММП «Обмен веществ и энергии у животных» .
Определение темы и задач урока через актуализацию опорных знаний. (Учащиеся отвечают на вопросы, учитель кратко записывает их ответы на доске). Примерные ответы учащихся выделены курсивом.
1.Вспомните и перечислите признаки, характерные для живых организмов.
(Питание, дыхание, выделение, обмен веществ, размножение, раздражимость, рост и развитие, движение).
2. Какие из них мы уже изучили?
Какие из них мы уже изучили?
(Питание, дыхание, транспорт веществ, выделение).
3.Попробуйте сформулировать понятие каждого из них.
Питание – это процесс получения органических веществ и заключённой в них энергии.
Дыхание – это получение энергии из пищи с участием кислорода. И т.д.
4. Питание, дыхание, выделение – всё это стороны одного самого главного признака живых существ, какого?
Самый главный признак живых организмов – это обмен веществ.
5. Постарайтесь вспомнить его понятие.
Обмен веществ – это поступление пищи и кислорода из среды, их использование для получения новых веществ и энергии, выделение ненужных веществ в среду.
Вот мы и определили тему нашего урока. Тема урока – Обмен веществ. А теперь попробуйте определить задачи, которые нам предстоит решить.
Тема урока – Обмен веществ. А теперь попробуйте определить задачи, которые нам предстоит решить.
Какие вещества поступают в растения и животным из среды? Для чего они используются? Какие вещества организмы выделяют в среду? Как происходит обмен веществ у растений и животных? И т.д.
Учитель: Итак, сегодня мы начинаем изучать тему «Обмен веществ у живых организмов». Изучать вы её будете используя учебники. В ходе урока нам вместе предстоит составить опорный конспект по теме. Работа с рисунками на ст. 78-79 учебника.
Вопросы для обсуждения темы «Обмен веществ и энергии у растений»:
1. Какие вещества растение поглощает из окружающей среды? (Вода, минеральные соли, углекислый газ, кислород)
2.
3. Во что превращаются сахара? (Крахмал, клетчатка, белки и жиры). Учитель поясняет, что в процессе фотосинтеза образуются углеводы, а все остальные органические вещества образуются из них.
4. На что они идут (расходуются)? (Идут на построение новых клеток, расходуются в процессе дыхания, откладываются в семенах, плодах). Учитель продолжает подводить учеников к значимости процесса ОВ. Строятся новые клетки, следовательно, обмен веществ обеспечивает … ?. Рост. Расходуются в процессе дыхания, следовательно, обмен веществ обеспечивает растения …? энергией, а значит, все процессы жизнедеятельности растений. Откладываются в семенах, следовательно, …? запасаются для будущих «деток».
5. Что выделяют растения в окружающую среду? (Вода, углекислый газ, кислород).
Закрепление – работай «в паре», работай с классом.
Сделайте выводы: Что такое обмен веществ? Почему это основное свойство растений, как живых организмов?
Учащиеся совместно с учителем формулируют выводы: Обмен веществ – это основное свойство живых организмов, потому что:
1. Обеспечивает рост растений.
2. Является источником энергии – жизни.
3. Позволяет заботиться о «потомстве».
Первичное применение знаний– работай индивидуально. Составить и воспроизвести рассказ по рис. на ст. 78.
Вопросы для обсуждения темы «Обмен веществ и энергии у животных»:
1. Какие вещества животное поглощает из окружающей среды? (Белки, жиры, углеводы, вода и минеральные соли, кислород) Учитель дополняет: Животные, в отличие от растений, потребляют готовые ОВ.
Какие вещества животное поглощает из окружающей среды? (Белки, жиры, углеводы, вода и минеральные соли, кислород) Учитель дополняет: Животные, в отличие от растений, потребляют готовые ОВ.
2. Что выделяют животные в окружающую среду?(Вода, углекислый газ, мочевина).
3. Что происходит с веществами (белками, жирами, углеводами) в теле животных? Какая система органов принимает в этом участие? (Превращения (пищеварение) – переводятся в более простые соединения, растворимые в воде, и уже в таком виде усваиваются клеткой). Укажите систему органов. (Пищеварительная система).
4. Какие два противоположных процесса происходят в клетке с поступившими в неё веществами? (Из простых веществ образуются другие сложные вещества, т.е. они превращаются в вещества своего тела. Распад сложных веществ на простые. При этом выделяется энергия)
5. На что расходуется энергия, которая выделяется при распаде веществ? (Образование новых веществ, работа внутренних органов – поддержание процессов жизнедеятельности, поддержание температуры тела).
На что расходуется энергия, которая выделяется при распаде веществ? (Образование новых веществ, работа внутренних органов – поддержание процессов жизнедеятельности, поддержание температуры тела).
Подумайте, для чего необходимы организму животного новые вещества (Белки, жиры, углеводы)? (Рост и развитие)
- Прочитайте текст на стр. 78 – 81, устно заполните таблицу
«Особенности обмена веществ у животных»
|
Животные |
Скорость обмена веществ |
Количество энергии |
Температура тела |
Такие животные называются… |
|
|
|
|
|
|
|
|
|
|
|
|
Закрепление
1. Сделайте выводы: Что такое обмен веществ? Почему это основное свойство растений и животных, как живых организмов?
Сделайте выводы: Что такое обмен веществ? Почему это основное свойство растений и животных, как живых организмов?
Обмен веществ – это основное свойство живых организмов, потому что:
1. Обеспечивает рост и развитие.
2. Является источником энергии и строительного материала для живых организмов .
3. Позволяет заботиться о «потомстве».
2. Сделайте выводы: Чем обмен веществ у животных отличается от обмена веществ у растений? Почему ОВ основное свойство животных, как живых организмов?
Рефлексия:
Какие вопросы вызвали затруднение? Что вам было не понятно? Что запомнилось больше всего? На какой вопрос вы хотели бы ответить на следующем уроке?
Д.З. §11 — пересказ, отвечать на вопросы ст. 82.
82.
Индивидуальные рабочие карточки для обучающихся (учитель раздаёт каждому обучающемуся для удобства работы во время устных ответов, обсуждения, повторения; после урока обучающиеся забирают их домой, чтобы использовать для повторения д.з.)
Вопросы для обсуждения темы «Обмен веществ и энергии у растений»: рисунок ст.78
1. Какие вещества растение поглощает из окружающей среды? 2. Что образуется у растений в процессе фотосинтеза из поступивших неорганических веществ? 3. Во что превращаются сахара?
Во что превращаются сахара?
4. На что они идут (расходуются)? 5. Что выделяют растения в окружающую среду?
Сделайте выводы: Что такое обмен веществ? Почему это основное свойство растений, как живых организмов?
Вопросы для обсуждения темы «Обмен веществ и энергии у животных»: рисунок ст.79
1. Какие вещества животное поглощает из окружающей среды?
2. Что выделяют животные в окружающую среду?
3. Что происходит с веществами (белками, жирами, углеводами) в теле животных? Какая система органов принимает в этом участие?
4. Какие два противоположных процесса происходят в клетке с поступившими в неё веществами? 5. На что расходуется энергия, которая выделяется при распаде веществ?
Какие два противоположных процесса происходят в клетке с поступившими в неё веществами? 5. На что расходуется энергия, которая выделяется при распаде веществ?
Подумайте, для чего необходимы организму животного новые вещества (Белки, жиры, углеводы)? (Рост и развитие)
- Прочитайте текст на стр. 78 – 81, устно заполните таблицу
«Особенности обмена веществ у животных»
|
Животные |
Скорость обмена веществ |
Количество энергии |
Температура тела |
Такие животные называются… |
Российские биохимики разработали сверхточный биосенсор для изучения обмена веществ в организме
Перекись водорода H2О2 — одна из важнейших молекул в организме живых существ.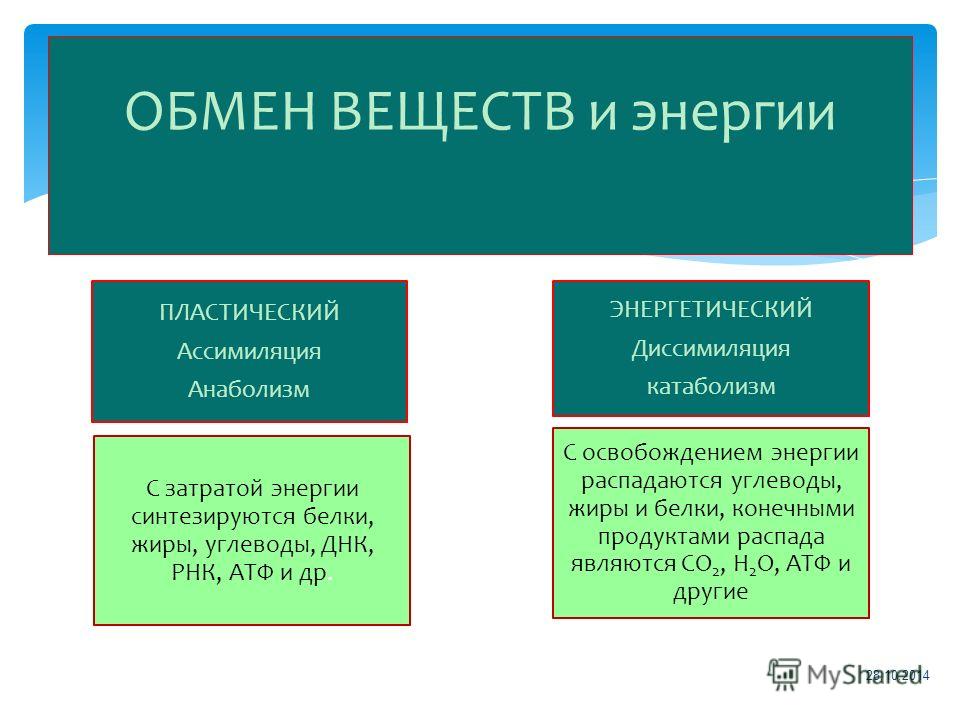 Она образуется в процессе метаболизма, а также выделяется специализированными ферментными системами. В организме это соединение регулирует работу различных биохимических процессов, но при слишком высоком ее содержании может произойти так называемый окислительный стресс — избыток активных форм кислорода в организме. Он участвует в развитии многих заболеваний, таких как воспалительные, онкологические и нейродегенеративные. Регуляторную функцию молекулы выполняют, находясь в чрезвычайно низких концентрациях, из-за высокой скорости реакции они живут максимум несколько секунд. Эти особенности перекиси водорода сильно затрудняют изучение ее влияния на метаболизм и возникновение патологий.
Она образуется в процессе метаболизма, а также выделяется специализированными ферментными системами. В организме это соединение регулирует работу различных биохимических процессов, но при слишком высоком ее содержании может произойти так называемый окислительный стресс — избыток активных форм кислорода в организме. Он участвует в развитии многих заболеваний, таких как воспалительные, онкологические и нейродегенеративные. Регуляторную функцию молекулы выполняют, находясь в чрезвычайно низких концентрациях, из-за высокой скорости реакции они живут максимум несколько секунд. Эти особенности перекиси водорода сильно затрудняют изучение ее влияния на метаболизм и возникновение патологий.
Разработка российских ученых — плод многолетней работы над биосенсорами, позволяющими наблюдать за пероксидом водорода в клетке с помощью оптики, например микроскопа. Первое поколение отечественных зондов получило мировое признание и расширило знания ученых в области метаболизма, но имело существенный недостаток — они не могли обнаружить сверхнизкие концентрации H2О2, с помощью которых реализуется большинство ее функций. Также результаты наблюдения могли быть неточными из-за того, что работа сенсора сильно зависела от кислотности среды.
Также результаты наблюдения могли быть неточными из-за того, что работа сенсора сильно зависела от кислотности среды.
Обычно биосенсоры — это приборы, которые находят химические соединения с помощью веществ, реагирующих на эти соединения. Например, биосенсоры используются для измерения количества сахара или алкоголя в крови. Но биосенсоры, о которых идет речь в исследовании, являются белковыми молекулами. В основе такого зонда ученые использовали зеленый флуоресцентный белок GFP, способный поглощать свет и через несколько наносекунд испускать его обратно с немного увеличенной длиной волны, то есть «светиться». Такой белок сшивается с другим белком, OxyR, природным сенсором пероксида водорода. В результате «свечение» (флуоресценция) GFP меняет свои свойства при взаимодействии зонда с H2О2. OxyR есть почти у всех бактерий. Поскольку исследователи не знали заранее, какой из белков окажется наиболее чувствительным, то выбрали для эксперимента 11 OxyR из разных неродственных друг другу видов микроорганизмов. В итоге сверхчувствительный сенсор получился при сшивании флуоресцентного белка GFP с белком OxyR, содержащимся в бактерии менингококк Neisseria meningitidis, обитающей в носоглотке человека и вызывающей менингит и назофарингит. OxyR этой бактерии реагирует на окисление пероксидом водорода, но при этом нечувствителен к другим активным формам кислорода.
В итоге сверхчувствительный сенсор получился при сшивании флуоресцентного белка GFP с белком OxyR, содержащимся в бактерии менингококк Neisseria meningitidis, обитающей в носоглотке человека и вызывающей менингит и назофарингит. OxyR этой бактерии реагирует на окисление пероксидом водорода, но при этом нечувствителен к другим активным формам кислорода.
«По-видимому, менингококк в процессе эволюции выработал сверхчувствительный OxyR, строго избирательный к пероксиду водорода. Это нужно для того, чтобы заранее чувствовать активацию нейтрофилов и макрофагов, иммунных клеток организма хозяина, использующих пероксид водорода, чтобы убить бактерию. В ответ менингококк способен усиливать свои антиоксидантные системы, защищающие его от окислительного стресса», — рассказывает Всеволод Белоусов, руководитель проекта по гранту РНФ, доктор биологических наук, заведующий отделом метаболизма и редокс-биологии ИБХ имени М. М. Шемякина и Ю.
А. Овчинникова РАН, и.о. директора Федерального центра мозга и нейротехнологий МЗ РФ.
 А. Овчинникова РАН, и.о. директора Федерального центра мозга и нейротехнологий МЗ РФ.
А. Овчинникова РАН, и.о. директора Федерального центра мозга и нейротехнологий МЗ РФ.
По словам ученого, новый зонд HyPer7 — настоящий прорыв в сфере биосенсоров: «Мы получили уникальный индикатор с высокой яркостью и коротким временем отклика, реагирующий на сверхнизкие концентрации перекиси водорода, при этом не зависящий от кислотной среды. HyPer7 оказался в 30 раз чувствительнее и в 80 раз быстрее предыдущих версий».
С помощью сенсора авторам работы уже удалось изучить детали перемещения молекул H2О2 внутри клетки, исследовать роль градиентов пероксида водорода в движении клеток и при повреждении тканей. Разработка биохимиков позволит визуализировать перекись водорода в клетках, тканях и органах, и исследовать роль этой молекулы в нормальных биохимических процессах. Также будет изучена роль перекиси водорода в возникновении и развитии ишемических, нейродегенеративных, воспалительных и онкологических заболеваний, которые тесно связаны с окислительным стрессом. В исследовании российским ученым помогали коллеги из Бельгии, Франции, Германии и США.
В исследовании российским ученым помогали коллеги из Бельгии, Франции, Германии и США.
Обмен веществ и энергии
Основные понятия
Определение 1
Обмен веществ и энергии, или метаболизм – физиологические процессы обеспечения организма необходимыми для его нормального функционирования соединениями, их превращение, получение энергии и выведения во внешнюю среду ненужных соединений произошедших реакций.
В узком смысле, метаболизм – это пути превращений определенного соединения или соединений в организме.
Метаболизм состоит из двух процессов:
- Пластический обмен, анаболизм, ассимиляция, или синтез. Это поступление в организм через пищеварительную систему воды, белков, жиров, углеводов, минеральных солей, витаминов, через дыхательную систему, кожу — кислорода для построения мембран, клеточных структур и их обновления. Анаболические реакции – это реакции, участвующие в синтезе новых молекул, протекают с поглощением энергии.
- Энергетический обмен, катаболизм, диссимиляция, или распад. Это процессы выведения из организма отработанных продуктов, осуществляется через органы пищеварительного тракта, легкие, почки, кожу. Катаболические реакции – это реакции распада, протекающие с выделением энергии. Во время процессов энергетического обмена часть энергии рассеивается в виде тепла, а часть запасается в определенных органических веществах в виде макроэргических связей. Универсальным химическим аккумулятором энергии является АТФ – аденозинтрифосфорная кислота.
Все реакции анаболизма и катаболизма протекают с помощью энзимов (ферментов) – биологических катализаторов.
В процессе обмена веществ постоянно образуются, обновляются, расщепляются клеточные структуры, появляются и разрушаются разнообразные химические соединения. Все это сопровождается превращениями энергии: потенциальная энергия веществ, освобождаемая при расщеплении, переходит в кинетическую энергию, представленную, главным образом тепловой и механической энергиями, частично – электрической энергией.
Поступление в организм различных веществ из внешней среды необходимо для:
- Возмещения энергозатрат.
- Удовлетворения потребностей роста
- Сохранения массы тела.
При этом количество питательных веществ, их соотношение и свойства должны согласовываться с условиями жизни и общим состоянием организма.
Все реакции пластического и энергетического обмена протекают совместно, переходя друг в друга в организме в течение всей жизни. В раннем возрасте преобладают реакции анаболизма, когда наблюдается интенсивный рост и развитие организма. По мере старения в организме начинают преобладать процессы катаболизма, синтез новых веществ постепенно угнетается.
Виды обмена веществ
Основными веществами, поступающими в организм человека, являются вода, минеральные соли, органические вещества: белки, витамины, углеводы и жиры. Для каждого вещества характерен свой путь метаболизма.
Существуют следующие виды обмена веществ:
- обмен воды и минеральных солей;
- обмен белков;
- обмен жиров;
- обмен углеводов.

Замечание 1
Большинство витаминов входят в состав ферментов, поэтому они выполняют в основном функцию катализаторов биохимических процессов.
Регуляция обмена веществ
Под регуляцией обмена веществ рассматривается регуляция почти всех функций организма: пищеварения, кровообращения, дыхания, выделения и др.
Основную роль в регуляции обмена веществ играет эндокринная система. Гормоны оказывают воздействие на скорость протекания биохимических процессов непосредственно в клетке. При совокупном их воздействии на отдельные клетки происходит изменение в функционировании организма в целом. К примеру,
- гормон гипофиза — соматотропный гормон проявляет выраженное анаболическое действие, он повышает синтез пластических веществ, ускоряет рост;
- катехоламины надпочечников усиливают энергообразование через окислительные процессы;
- тироксин и трийодтиронин – гормоны щитовидной железы – активируют разрушение углеводов и жиров, стимулируют образование белка из аминокислот.

В регуляции обмена веществ принимает участие нервная система – гипоталамус, который включает центры жажды, голода и насыщения, терморегуляции. Регуляция осуществляется через вегетативную нервную систему.
Замечание 2
Гипоталамус и гипофиз координируют функционирование почти всех желез внутренней секреции.
Тест: Обмен веществ. — Биология 8 класс
Обмен веществ.
Один вариант ответа из нескольких предложенных. Для получения «5» необходимо правильно ответить не менее чем на 90% вопросов,
Биология 8 класс | ID: 1156 | Дата: 25.1.2014
«;} else {document.getElementById(«torf1″).innerHTML=»»;}; if (answ.charAt(1)==»1″) {document.getElementById(«torf2″).innerHTML=»»;} else {document.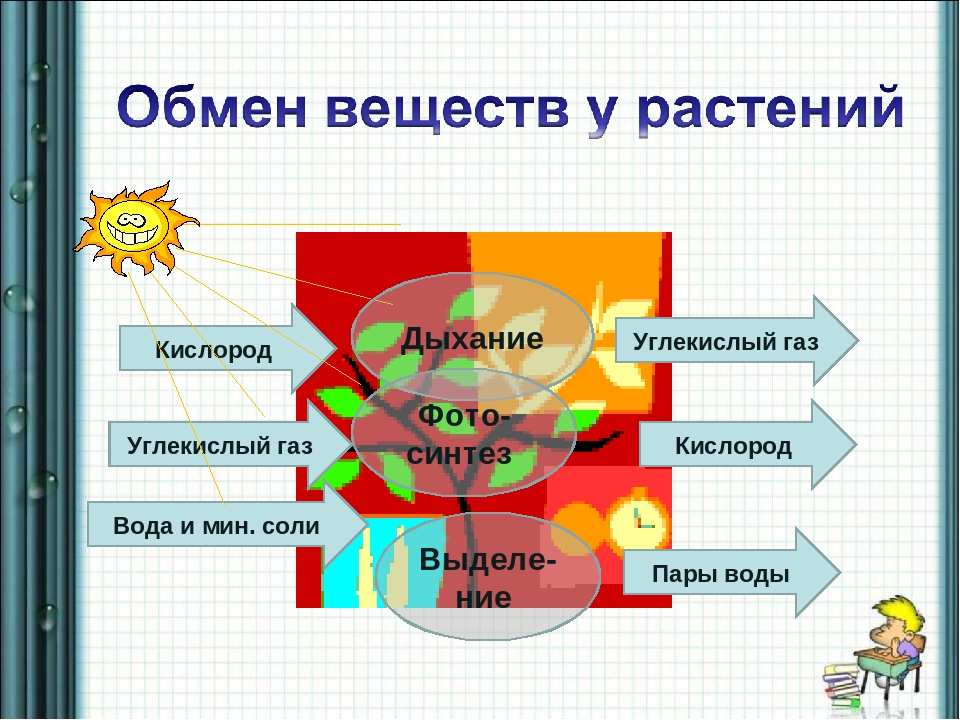 getElementById(«torf2″).innerHTML=»»;};
if (answ.charAt(2)==»1″) {document.getElementById(«torf3″).innerHTML=»»;} else {document.getElementById(«torf3″).innerHTML=»»;};
if (answ.charAt(3)==»1″) {document.getElementById(«torf4″).innerHTML=»»;} else {document.getElementById(«torf4″).innerHTML=»»;};
if (answ.charAt(4)==»1″) {document.getElementById(«torf5″).innerHTML=»»;} else {document.getElementById(«torf5″).innerHTML=»»;};
if (answ.charAt(5)==»1″) {document.getElementById(«torf6″).innerHTML=»»;} else {document.getElementById(«torf6″).innerHTML=»»;};
if (answ.charAt(6)==»1″) {document.getElementById(«torf7″).innerHTML=»»;} else {document.getElementById(«torf7″).innerHTML=»»;};
if (answ.charAt(7)==»1″) {document.getElementById(«torf8″).innerHTML=»»;} else {document.getElementById(«torf8″).innerHTML=»»;};
if (answ.charAt(8)==»1″) {document.getElementById(«torf9″).innerHTML=»»;} else {document.getElementById(«torf9″).innerHTML=»»;};
if (answ.charAt(9)==»1″) {document.getElementById(«torf10»).
getElementById(«torf2″).innerHTML=»»;};
if (answ.charAt(2)==»1″) {document.getElementById(«torf3″).innerHTML=»»;} else {document.getElementById(«torf3″).innerHTML=»»;};
if (answ.charAt(3)==»1″) {document.getElementById(«torf4″).innerHTML=»»;} else {document.getElementById(«torf4″).innerHTML=»»;};
if (answ.charAt(4)==»1″) {document.getElementById(«torf5″).innerHTML=»»;} else {document.getElementById(«torf5″).innerHTML=»»;};
if (answ.charAt(5)==»1″) {document.getElementById(«torf6″).innerHTML=»»;} else {document.getElementById(«torf6″).innerHTML=»»;};
if (answ.charAt(6)==»1″) {document.getElementById(«torf7″).innerHTML=»»;} else {document.getElementById(«torf7″).innerHTML=»»;};
if (answ.charAt(7)==»1″) {document.getElementById(«torf8″).innerHTML=»»;} else {document.getElementById(«torf8″).innerHTML=»»;};
if (answ.charAt(8)==»1″) {document.getElementById(«torf9″).innerHTML=»»;} else {document.getElementById(«torf9″).innerHTML=»»;};
if (answ.charAt(9)==»1″) {document.getElementById(«torf10»). innerHTML=»»;} else {document.getElementById(«torf10″).innerHTML=»»;};
if (answ.charAt(10)==»1″) {document.getElementById(«torf11″).innerHTML=»»;} else {document.getElementById(«torf11″).innerHTML=»»;};
if (answ.charAt(11)==»1″) {document.getElementById(«torf12″).innerHTML=»»;} else {document.getElementById(«torf12″).innerHTML=»»;};
if (answ.charAt(12)==»1″) {document.getElementById(«torf13″).innerHTML=»»;} else {document.getElementById(«torf13″).innerHTML=»»;};
if (answ.charAt(13)==»1″) {document.getElementById(«torf14″).innerHTML=»»;} else {document.getElementById(«torf14″).innerHTML=»»;};
if (answ.charAt(14)==»1″) {document.getElementById(«torf15″).innerHTML=»»;} else {document.getElementById(«torf15″).innerHTML=»»;};
}
}
innerHTML=»»;} else {document.getElementById(«torf10″).innerHTML=»»;};
if (answ.charAt(10)==»1″) {document.getElementById(«torf11″).innerHTML=»»;} else {document.getElementById(«torf11″).innerHTML=»»;};
if (answ.charAt(11)==»1″) {document.getElementById(«torf12″).innerHTML=»»;} else {document.getElementById(«torf12″).innerHTML=»»;};
if (answ.charAt(12)==»1″) {document.getElementById(«torf13″).innerHTML=»»;} else {document.getElementById(«torf13″).innerHTML=»»;};
if (answ.charAt(13)==»1″) {document.getElementById(«torf14″).innerHTML=»»;} else {document.getElementById(«torf14″).innerHTML=»»;};
if (answ.charAt(14)==»1″) {document.getElementById(«torf15″).innerHTML=»»;} else {document.getElementById(«torf15″).innerHTML=»»;};
}
}Получение сертификата
о прохождении теста
Энергия и обмен веществ | Безграничная биология
Роль энергии и метаболизма
Всем организмам требуется энергия для выполнения задач; метаболизм — это набор химических реакций, высвобождающих энергию для клеточных процессов.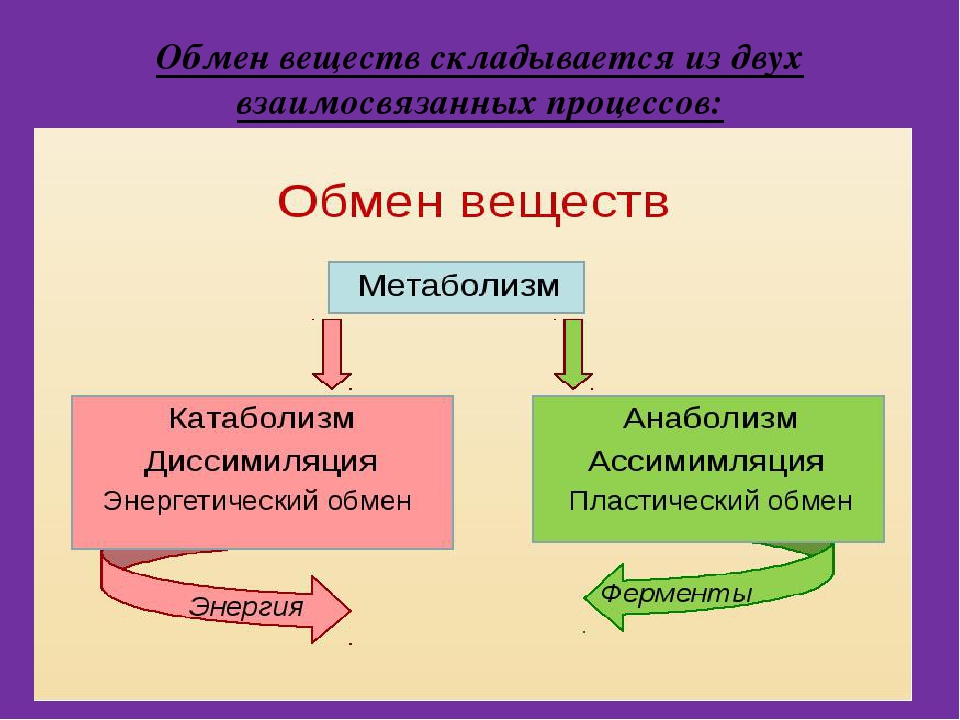
Цели обучения
Объясните важность обмена веществ
Основные выводы
Ключевые моменты
- Все живые организмы нуждаются в энергии для роста и воспроизводства, поддержания своей структуры и реагирования на окружающую среду; метаболизм — это набор процессов, делающих энергию доступной для клеточных процессов.
- Метаболизм — это комбинация химических реакций, которые являются спонтанными и высвобождают энергию, и химических реакций, которые не являются спонтанными и требуют энергии для протекания.
- Живые организмы должны получать энергию через пищу, питательные вещества или солнечный свет, чтобы осуществлять клеточные процессы.
- Транспортировка, синтез и распад питательных веществ и молекул в клетке требуют использования энергии.
Ключевые термины
- метаболизм : полный набор химических реакций, происходящих в живых клетках
- биоэнергетика : исследование превращений энергии, происходящих в живых организмах
- энергия : работоспособность
Энергия и обмен веществ
Всем живым организмам нужна энергия для роста и размножения, поддержания своей структуры и реакции на окружающую среду.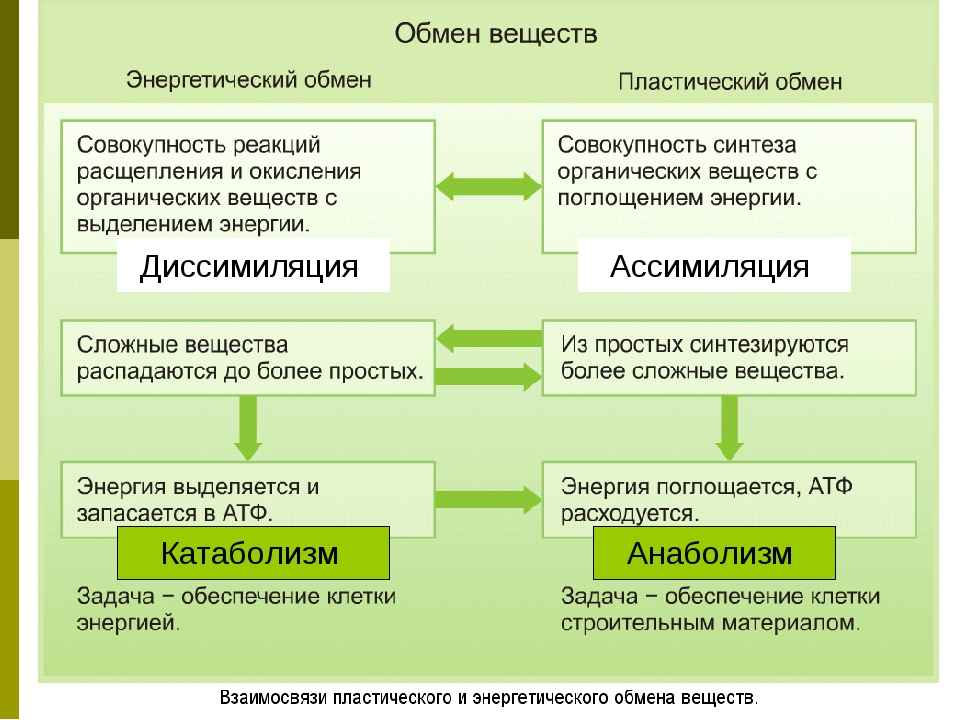 Метаболизм — это набор поддерживающих жизнь химических процессов, которые позволяют организмам преобразовывать химическую энергию, хранящуюся в молекулах, в энергию, которая может использоваться для клеточных процессов. Животные потребляют пищу, чтобы восполнить энергию; их метаболизм расщепляет углеводы, липиды, белки и нуклеиновые кислоты, чтобы обеспечить химическую энергию для этих процессов. В процессе фотосинтеза растения преобразуют световую энергию солнца в химическую энергию, хранящуюся в молекулах.
Метаболизм — это набор поддерживающих жизнь химических процессов, которые позволяют организмам преобразовывать химическую энергию, хранящуюся в молекулах, в энергию, которая может использоваться для клеточных процессов. Животные потребляют пищу, чтобы восполнить энергию; их метаболизм расщепляет углеводы, липиды, белки и нуклеиновые кислоты, чтобы обеспечить химическую энергию для этих процессов. В процессе фотосинтеза растения преобразуют световую энергию солнца в химическую энергию, хранящуюся в молекулах.
Биоэнергетика и химические реакции
Ученые используют термин «биоэнергетика» для обсуждения концепции потока энергии через живые системы, такие как клетки.Клеточные процессы, такие как построение и разрушение сложных молекул, происходят в результате пошаговых химических реакций. Некоторые из этих химических реакций являются спонтанными и высвобождают энергию, тогда как другие требуют энергии для протекания. Все химические реакции, происходящие внутри клеток, включая те, которые используют энергию, и те, которые высвобождают энергию, являются метаболизмом клетки.
Большая часть энергии прямо или косвенно исходит от Солнца. : Большинство форм жизни на Земле получают энергию от Солнца.Растения используют фотосинтез для улавливания солнечного света, а травоядные животные поедают эти растения для получения энергии. Плотоядные животные поедают травоядных, а разлагатели переваривают вещества растений и животных.
Клеточный метаболизм
Каждое задание, выполняемое живыми организмами, требует энергии. Энергия необходима для выполнения тяжелой работы и упражнений, но люди также расходуют много энергии во время размышлений и даже во время сна. Для каждого действия, требующего энергии, происходит множество химических реакций, обеспечивающих химическую энергию системам тела, включая мышцы, нервы, сердце, легкие и мозг.
Живые клетки каждого организма постоянно используют энергию для выживания и роста. Клетки расщепляют сложные углеводы на простые сахара, которые клетка может использовать для получения энергии. Мышечные клетки могут потреблять энергию для построения длинных мышечных белков из небольших молекул аминокислот. Молекулы можно модифицировать и переносить по клетке или можно распространять по всему организму. Так же, как энергия требуется как для строительства, так и для сноса здания, энергия требуется как для синтеза, так и для разрушения молекул.
Мышечные клетки могут потреблять энергию для построения длинных мышечных белков из небольших молекул аминокислот. Молекулы можно модифицировать и переносить по клетке или можно распространять по всему организму. Так же, как энергия требуется как для строительства, так и для сноса здания, энергия требуется как для синтеза, так и для разрушения молекул.
Многие клеточные процессы требуют постоянного снабжения энергией, обеспечиваемой метаболизмом клетки. Сигнальные молекулы, такие как гормоны и нейротрансмиттеры, должны быть синтезированы, а затем транспортироваться между клетками. Патогенные бактерии и вирусы попадают в организм и разрушаются клетками. Клетки также должны экспортировать отходы и токсины, чтобы оставаться здоровыми, и многие клетки должны плавать или перемещать окружающие материалы за счет биения клеточных придатков, таких как реснички и жгутики.
Питание дает энергию для таких действий, как полет. : Колибри нуждается в энергии, чтобы поддерживать длительные периоды полета. Колибри получает энергию, принимая пищу и превращая питательные вещества в энергию с помощью ряда биохимических реакций. Летательные мышцы птиц чрезвычайно эффективны в производстве энергии.
Колибри получает энергию, принимая пищу и превращая питательные вещества в энергию с помощью ряда биохимических реакций. Летательные мышцы птиц чрезвычайно эффективны в производстве энергии.
Типы энергии
Различные типы энергии включают кинетическую, потенциальную и химическую энергию.
Цели обучения
Различия между видами энергии
Основные выводы
Ключевые моменты
- Все организмы используют разные формы энергии для поддержания биологических процессов, которые позволяют им расти и выживать.
- Кинетическая энергия — это энергия, связанная с движущимися объектами.
- Потенциальная энергия — это тип энергии, связанный со способностью объекта выполнять работу.
- Химическая энергия — это тип энергии, высвобождающейся при разрыве химических связей, который может быть использован для метаболических процессов.
Ключевые термины
- химическая энергия : чистая потенциальная энергия, выделяемая или поглощаемая в ходе химической реакции.
- потенциальная энергия : энергия, которой обладает объект из-за его положения (в гравитационном или электрическом поле) или его состояния (в виде растянутой или сжатой пружины, в качестве химического реагента или из-за наличия массы покоя).
- кинетическая энергия : энергия, которой обладает объект из-за его движения, равная половине массы тела, умноженной на квадрат его скорости.

Энергия — это свойство объектов, которое может быть передано другим объектам или преобразовано в другие формы, но не может быть создано или уничтожено. Организмы используют энергию для выживания, роста, реакции на раздражители, воспроизводства и для всех типов биологических процессов. Потенциальная энергия, хранящаяся в молекулах, может быть преобразована в химическую энергию, которая в конечном итоге может быть преобразована в кинетическую энергию, позволяющую организму двигаться.В конце концов, большая часть энергии, используемой организмами, преобразуется в тепло и рассеивается.
Кинетическая энергия
Энергия, связанная с движущимися объектами, называется кинетической энергией. Например, когда самолет находится в полете, он очень быстро движется по воздуху, выполняя работу по изменению своего окружения. Реактивные двигатели преобразуют потенциальную энергию топлива в кинетическую энергию движения. Крушащий шар может нанести большой урон даже при медленном движении.Однако все еще разрушающийся шар не может выполнять никакой работы и, следовательно, не имеет кинетической энергии. Ускоряющаяся пуля, идущий человек, быстрое движение молекул в воздухе, выделяющих тепло, и электромагнитное излучение, такое как солнечный свет, — все они обладают кинетической энергией.
Потенциальная энергия
Что, если тот же самый неподвижный шар для разрушения поднять на два этажа над автомобилем с краном? Если подвешенный шар для разрушения не движется, связана ли с ним энергия? Да, разрушающий шар обладает энергией, потому что разрушающий шар может выполнять свою работу.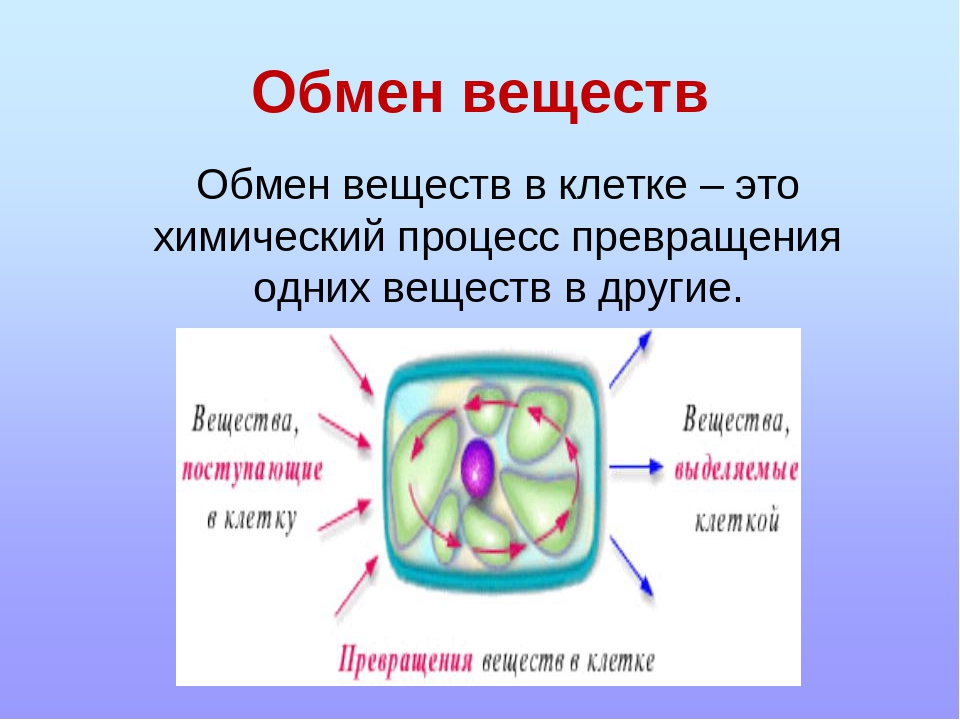 Эта форма энергии называется потенциальной энергией, потому что объект может выполнять работу в данном состоянии.
Эта форма энергии называется потенциальной энергией, потому что объект может выполнять работу в данном состоянии.
Объекты переносят свою энергию между потенциальным и кинетическим состояниями. Поскольку разрушающий шар неподвижно висит, он имеет кинетическую энергию [latex] \ text {0%} [/ latex] и [latex] \ text {100%} [/ latex]. Как только мяч выпущен, его кинетическая энергия увеличивается по мере того, как мяч набирает скорость. В то же время мяч теряет потенциальную энергию при приближении к земле. Другие примеры потенциальной энергии включают энергию воды, удерживаемой за плотиной, или человека, который собирается прыгнуть с парашютом из самолета.
Зависимость потенциальной энергии от кинетической энергии : Вода за плотиной имеет потенциальную энергию. Движущаяся вода, например, в водопаде или в быстро текущей реке, обладает кинетической энергией.
Химическая энергия
Потенциальная энергия связана не только с местонахождением материи, но и со структурой материи. Пружина на земле обладает потенциальной энергией, если она сжимается, как и натянутая резинка. Тот же принцип применим к молекулам. На химическом уровне связи, которые удерживают атомы молекул вместе, обладают потенциальной энергией.Этот тип потенциальной энергии называется химической энергией, и, как и вся потенциальная энергия, ее можно использовать для выполнения работы.
Пружина на земле обладает потенциальной энергией, если она сжимается, как и натянутая резинка. Тот же принцип применим к молекулам. На химическом уровне связи, которые удерживают атомы молекул вместе, обладают потенциальной энергией.Этот тип потенциальной энергии называется химической энергией, и, как и вся потенциальная энергия, ее можно использовать для выполнения работы.
Например, химическая энергия содержится в молекулах бензина, которые используются в автомобилях. Когда газ воспламеняется в двигателе, связи в его молекулах разрываются, и выделяемая энергия используется для приведения в движение поршней. Потенциальная энергия, хранящаяся в химических связях, может быть использована для выполнения работы для биологических процессов. Различные метаболические процессы разрушают органические молекулы, чтобы высвободить энергию для роста и выживания организма.
Химическая энергия : Молекулы бензина (октановое число, указанная химическая формула) содержат химическую энергию.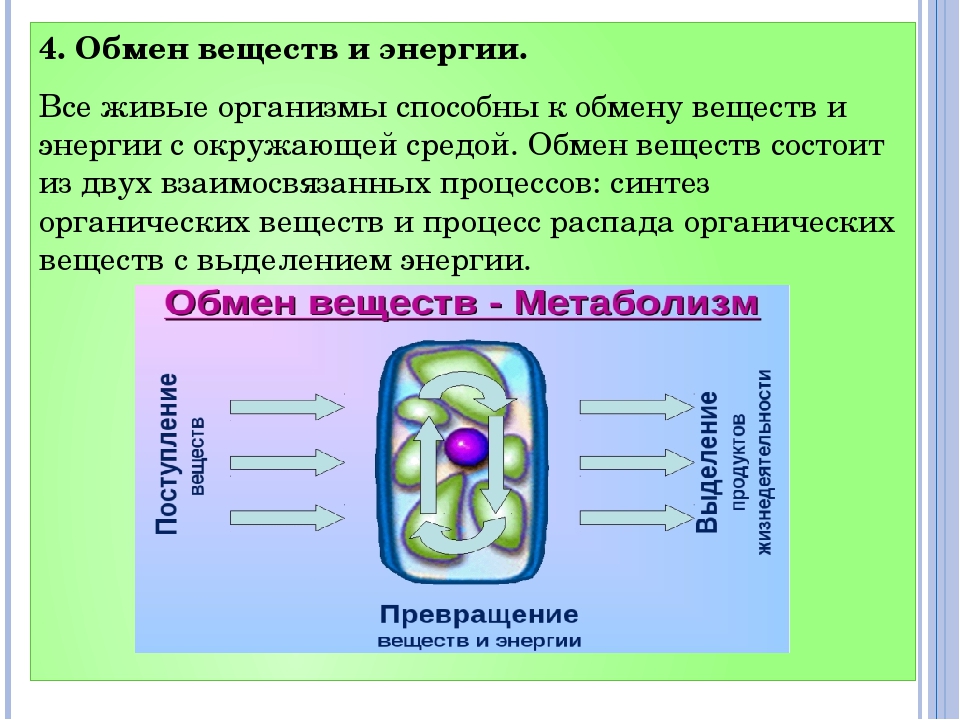 Эта энергия преобразуется в кинетическую энергию, которая позволяет автомобилю мчаться по гоночной трассе.
Эта энергия преобразуется в кинетическую энергию, которая позволяет автомобилю мчаться по гоночной трассе.
Метаболические пути
Анаболический путь требует энергии и строит молекулы, в то время как катаболический путь производит энергию и разрушает молекулы.
Цели обучения
Опишите два основных типа метаболических путей
Основные выводы
Ключевые моменты
- Метаболический путь — это серия химических реакций в клетке, которые создают и разрушают молекулы для клеточных процессов.
- Анаболические пути синтезируют молекулы и требуют энергии.
- Катаболические пути расщепляют молекулы и производят энергию.
- Поскольку почти все метаболические реакции протекают не спонтанно, белки, называемые ферментами, помогают облегчить эти химические реакции.
Ключевые термины
- катаболизм : деструктивный метаболизм, обычно включающий выделение энергии и расщепление материалов
- фермент : глобулярный белок, катализирующий биологическую химическую реакцию
- анаболизм : конструктивный метаболизм тела в отличие от катаболизма
Метаболические пути
Процессы производства и расщепления углеводных молекул иллюстрируют два типа метаболических путей. Метаболический путь — это последовательный ряд взаимосвязанных биохимических реакций, которые преобразуют молекулу или молекулы субстрата через ряд промежуточных продуктов метаболизма, в конечном итоге приводя к конечному продукту или продуктам. Например, один из метаболических путей углеводов расщепляет большие молекулы на глюкозу. Другой метаболический путь может превращать глюкозу в большие молекулы углеводов для хранения. Первый из этих процессов требует энергии и называется анаболическим. Второй процесс производит энергию и называется катаболическим.Следовательно, метаболизм состоит из этих двух противоположных путей:
Метаболический путь — это последовательный ряд взаимосвязанных биохимических реакций, которые преобразуют молекулу или молекулы субстрата через ряд промежуточных продуктов метаболизма, в конечном итоге приводя к конечному продукту или продуктам. Например, один из метаболических путей углеводов расщепляет большие молекулы на глюкозу. Другой метаболический путь может превращать глюкозу в большие молекулы углеводов для хранения. Первый из этих процессов требует энергии и называется анаболическим. Второй процесс производит энергию и называется катаболическим.Следовательно, метаболизм состоит из этих двух противоположных путей:
- Анаболизм (строительные молекулы)
- Катаболизм (разрушение молекул)
Анаболические и катаболические пути : Анаболические пути — это те пути, которые требуют энергии для синтеза более крупных молекул. Катаболические пути — это те, которые генерируют энергию за счет разрушения более крупных молекул. Оба типа путей необходимы для поддержания энергетического баланса клетки.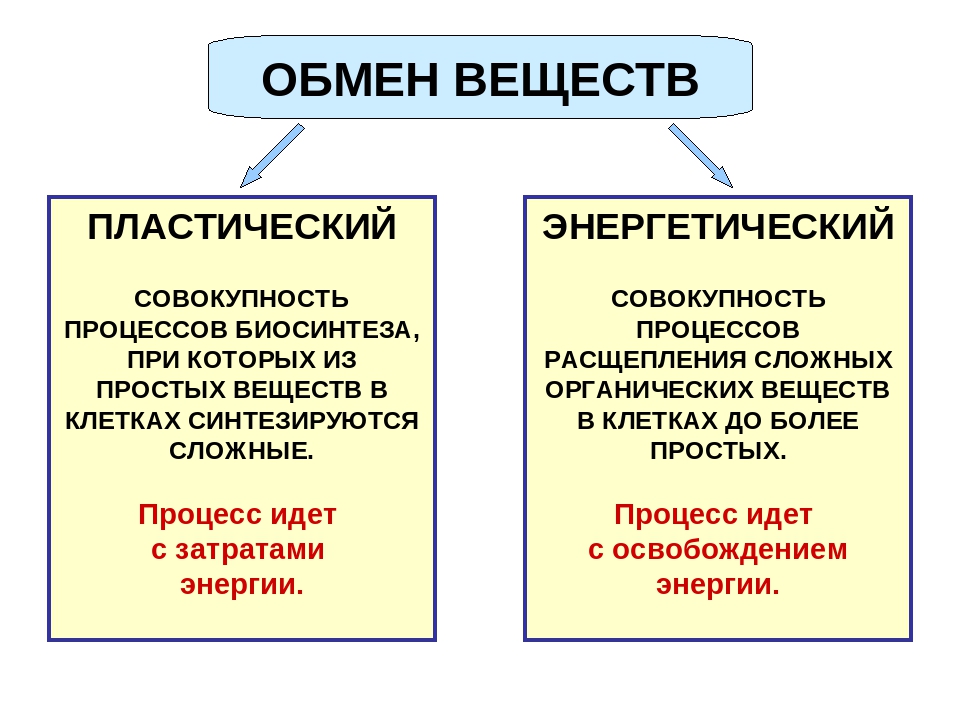
Анаболические пути
Анаболические пути требуют ввода энергии для синтеза сложных молекул из более простых.Одним из примеров анаболического пути является синтез сахара из CO 2 . Другие примеры включают синтез больших белков из строительных блоков аминокислот и синтез новых цепей ДНК из строительных блоков нуклеиновых кислот. Эти процессы имеют решающее значение для жизни клетки, происходят постоянно и требуют энергии, обеспечиваемой АТФ и другими высокоэнергетическими молекулами, такими как НАДН (никотинамидадениндинуклеотид) и НАДФН.
Катаболические пути
Катаболические пути включают разложение сложных молекул на более простые, высвобождая химическую энергию, хранящуюся в связях этих молекул.Некоторые катаболические пути могут улавливать эту энергию для производства АТФ, молекулы, используемой для питания всех клеточных процессов. Другие запасающие энергию молекулы, такие как липиды, также расщепляются с помощью аналогичных катаболических реакций с выделением энергии и образованием АТФ.
Важность ферментов
Химические реакции метаболических путей редко происходят спонтанно. Каждая стадия реакции облегчается или катализируется белком, называемым ферментом. Ферменты важны для катализирования всех типов биологических реакций: тех, которые требуют энергии, а также тех, которые выделяют энергию.
Метаболизм углеводов
Организмы расщепляют углеводы для производства энергии для клеточных процессов, а фотосинтезирующие растения производят углеводы.
Цели обучения
Анализируйте важность углеводного обмена для производства энергии
Основные выводы
Ключевые моменты
- Распад глюкозы, которую живые организмы используют для производства энергии, описывается уравнением: [латекс] {\ text {C}} _ {6} {\ text {H}} _ {12} {\ text {O}} _ {6} +6 {\ text {O}} _ {2} \ rightarrow 6 {\ text {CO}} _ {2} +6 {\ text {H}} _ {2} \ text {O} + \ text {энергия} [/ латекс].
- Процесс фотосинтеза, который растения используют для синтеза глюкозы, описывается уравнением: [латекс] 6 \ text {CO} _ {2} +6 {\ text {H}} _ {2} \ text {O} + \ text { энергия} \ rightarrow {\ text {C}} _ {6} {\ text {H}} _ {12} {\ text {O}} _ {6} +6 \ text {O} _ {2} [/ латекс].
- Потребляемая глюкоза используется для производства энергии в виде АТФ, который используется для выполнения работы и химических реакций в клетке.
- Во время фотосинтеза растения превращают световую энергию в химическую энергию, которая используется для создания молекул глюкозы.
Ключевые термины
- аденозинтрифосфат : многофункциональный нуклеозидтрифосфат, используемый в клетках в качестве кофермента, часто называемый «молекулярной единицей энергетической валюты» при внутриклеточном переносе энергии
- глюкоза : простой моносахарид (сахар) с молекулярной формулой C6h22O6; это основной источник энергии для клеточного метаболизма
Метаболизм углеводов
Углеводы — одна из основных форм энергии для животных и растений.Растения вырабатывают углеводы, используя световую энергию солнца (в процессе фотосинтеза), в то время как животные едят растения или других животных, чтобы получить углеводы.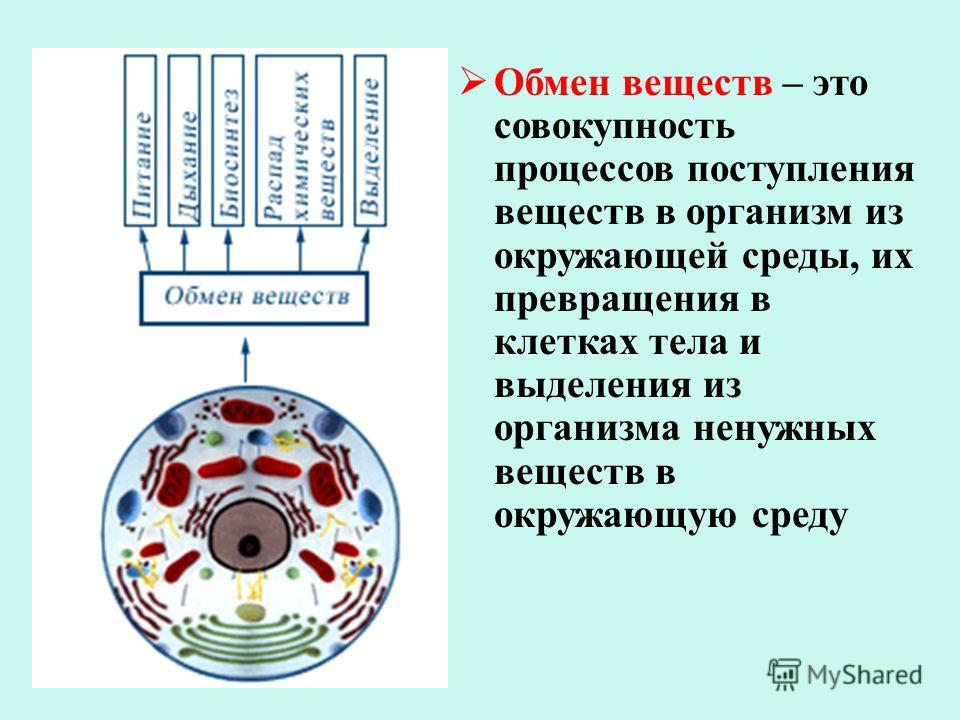 Растения хранят углеводы в длинных полисахаридных цепях, называемых крахмалом, а животные хранят углеводы в виде молекулы гликогена. Эти большие полисахариды содержат много химических связей и, следовательно, хранят много химической энергии. Когда эти молекулы расщепляются во время метаболизма, энергия химических связей высвобождается и может использоваться для клеточных процессов.
Растения хранят углеводы в длинных полисахаридных цепях, называемых крахмалом, а животные хранят углеводы в виде молекулы гликогена. Эти большие полисахариды содержат много химических связей и, следовательно, хранят много химической энергии. Когда эти молекулы расщепляются во время метаболизма, энергия химических связей высвобождается и может использоваться для клеточных процессов.
Все живые существа используют углеводы как форму энергии. : Растения, такие как дуб и желудь, используют энергию солнечного света для производства сахара и других органических молекул. И растения, и животные (например, эта белка) используют клеточное дыхание для получения энергии из органических молекул, изначально производимых растениями
Производство энергии из углеводов (клеточное дыхание)
Метаболизм любого моносахарида (простого сахара) может производить энергию для использования клеткой.Избыточные углеводы хранятся в виде крахмала в растениях и в виде гликогена у животных, готовые к метаболизму, если потребность организма в энергии внезапно возрастет. Когда эта потребность в энергии увеличивается, углеводы расщепляются на составляющие моносахариды, которые затем распределяются по всем живым клеткам организма. Глюкоза (C 6 H 12 O 6 ) является распространенным примером моносахаридов, используемых для производства энергии.
Когда эта потребность в энергии увеличивается, углеводы расщепляются на составляющие моносахариды, которые затем распределяются по всем живым клеткам организма. Глюкоза (C 6 H 12 O 6 ) является распространенным примером моносахаридов, используемых для производства энергии.
Внутри клетки каждая молекула сахара расщепляется в ходе сложной серии химических реакций.Поскольку химическая энергия высвобождается из связей в моносахариде, она используется для синтеза высокоэнергетических молекул аденозинтрифосфата (АТФ). АТФ — это основная энергетическая валюта всех клеток. Точно так же, как доллар используется в качестве валюты для покупки товаров, клетки используют молекулы АТФ для немедленной работы и проведения химических реакций.
Распад глюкозы во время метаболизма — клеточное дыхание — можно описать уравнением:
[латекс] {C} _ {6} {H} _ {12} {O} _ {6} +6 {O} _ {2} \ rightarrow 6 {CO} _ {2} +6 {H} _ {2} О + энергия [/ латекс]
Производство углеводов (фотосинтез)
Растения и некоторые другие виды организмов производят углеводы в процессе фотосинтеза.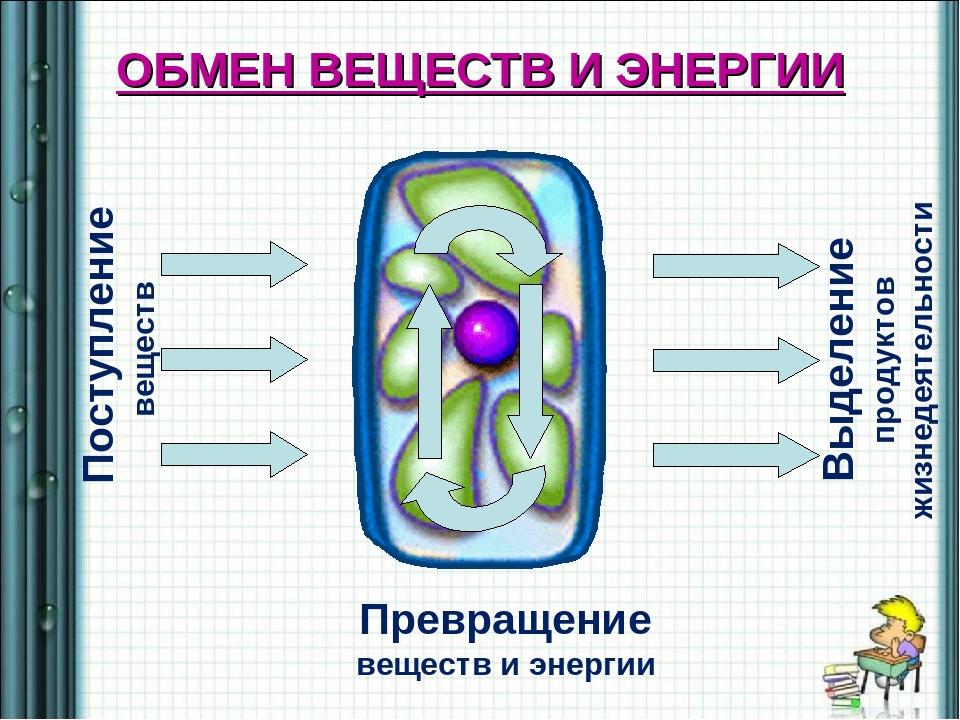 Во время фотосинтеза растения превращают световую энергию в химическую энергию, превращая молекулы углекислого газа (CO 2 ) в молекулы сахара, такие как глюкоза. Поскольку этот процесс включает в себя создание связей для синтеза большой молекулы, для его продолжения требуется ввод энергии (света). Синтез глюкозы путем фотосинтеза описывается этим уравнением (обратите внимание, что оно является обратным предыдущему уравнению):
Во время фотосинтеза растения превращают световую энергию в химическую энергию, превращая молекулы углекислого газа (CO 2 ) в молекулы сахара, такие как глюкоза. Поскольку этот процесс включает в себя создание связей для синтеза большой молекулы, для его продолжения требуется ввод энергии (света). Синтез глюкозы путем фотосинтеза описывается этим уравнением (обратите внимание, что оно является обратным предыдущему уравнению):
[латекс] 6CO_ {2} +6 {H} _ {2} O + энергия \ rightarrow {C} _ {6} {H} _ {12} {O} _ {6} + 6O_ {2} [/ латекс]
В рамках химических процессов растений молекулы глюкозы могут объединяться и превращаться в другие типы сахаров.В растениях глюкоза хранится в форме крахмала, который может быть снова расщеплен на глюкозу посредством клеточного дыхания для обеспечения АТФ.
6.1A: роль энергии и метаболизма
- Последнее обновление
- Сохранить как PDF
- Ключевые моменты
- Ключевые термины
- Энергия и метаболизм
- Биоэнергетика и химические реакции
- Клеточный метаболизм
Всем организмам требуется энергия для выполнения задач; метаболизм — это набор химических реакций, высвобождающих энергию для клеточных процессов.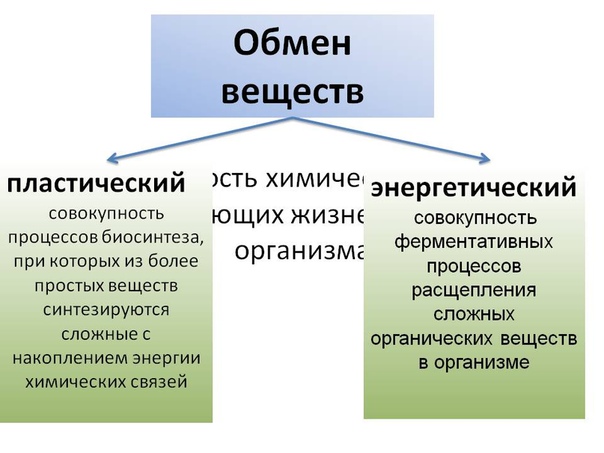
Задачи обучения
- Объяснить важность метаболизма
Ключевые моменты
- Все живые организмы нуждаются в энергии для роста и воспроизводства, поддержания своей структуры и реагирования на окружающую среду; метаболизм — это набор процессов, делающих энергию доступной для клеточных процессов.
- Метаболизм — это комбинация химических реакций, которые являются спонтанными и высвобождают энергию, и химических реакций, которые не являются спонтанными и требуют энергии для протекания.
- Живые организмы должны получать энергию через пищу, питательные вещества или солнечный свет, чтобы осуществлять клеточные процессы.
- Транспортировка, синтез и распад питательных веществ и молекул в клетке требуют использования энергии.
Ключевые термины
- метаболизм : полный набор химических реакций, происходящих в живых клетках
- биоэнергетика : исследование превращений энергии, происходящих в живых организмах
- энергия : работоспособность
Энергия и обмен веществ
Всем живым организмам нужна энергия для роста и размножения, поддержания своей структуры и реакции на окружающую среду. Метаболизм — это набор поддерживающих жизнь химических процессов, которые позволяют организмам преобразовывать химическую энергию, хранящуюся в молекулах, в энергию, которая может использоваться для клеточных процессов. Животные потребляют пищу, чтобы восполнить энергию; их метаболизм расщепляет углеводы, липиды, белки и нуклеиновые кислоты, чтобы обеспечить химическую энергию для этих процессов. В процессе фотосинтеза растения преобразуют световую энергию солнца в химическую энергию, хранящуюся в молекулах.
Метаболизм — это набор поддерживающих жизнь химических процессов, которые позволяют организмам преобразовывать химическую энергию, хранящуюся в молекулах, в энергию, которая может использоваться для клеточных процессов. Животные потребляют пищу, чтобы восполнить энергию; их метаболизм расщепляет углеводы, липиды, белки и нуклеиновые кислоты, чтобы обеспечить химическую энергию для этих процессов. В процессе фотосинтеза растения преобразуют световую энергию солнца в химическую энергию, хранящуюся в молекулах.
Биоэнергетика и химические реакции
Ученые используют термин «биоэнергетика» для обсуждения концепции потока энергии через живые системы, такие как клетки.Клеточные процессы, такие как построение и разрушение сложных молекул, происходят в результате пошаговых химических реакций. Некоторые из этих химических реакций являются спонтанными и высвобождают энергию, тогда как другие требуют энергии для протекания. Все химические реакции, происходящие внутри клеток, включая те, которые используют энергию, и те, которые высвобождают энергию, являются метаболизмом клетки.
Клеточный метаболизм
Каждое задание, выполняемое живыми организмами, требует энергии. Энергия необходима для выполнения тяжелой работы и упражнений, но люди также расходуют много энергии во время размышлений и даже во время сна. Для каждого действия, требующего энергии, происходит множество химических реакций, обеспечивающих химическую энергию системам тела, включая мышцы, нервы, сердце, легкие и мозг.
Живые клетки каждого организма постоянно используют энергию для выживания и роста. Клетки расщепляют сложные углеводы на простые сахара, которые клетка может использовать для получения энергии. Мышечные клетки могут потреблять энергию для построения длинных мышечных белков из небольших молекул аминокислот. Молекулы можно модифицировать и переносить по клетке или можно распространять по всему организму. Так же, как энергия требуется как для строительства, так и для сноса здания, энергия требуется как для синтеза, так и для разрушения молекул.
Мышечные клетки могут потреблять энергию для построения длинных мышечных белков из небольших молекул аминокислот. Молекулы можно модифицировать и переносить по клетке или можно распространять по всему организму. Так же, как энергия требуется как для строительства, так и для сноса здания, энергия требуется как для синтеза, так и для разрушения молекул.
Многие клеточные процессы требуют постоянного снабжения энергией, обеспечиваемой метаболизмом клетки. Сигнальные молекулы, такие как гормоны и нейротрансмиттеры, должны быть синтезированы, а затем транспортироваться между клетками. Патогенные бактерии и вирусы попадают в организм и разрушаются клетками. Клетки также должны экспортировать отходы и токсины, чтобы оставаться здоровыми, и многие клетки должны плавать или перемещать окружающие материалы за счет биения клеточных придатков, таких как реснички и жгутики.
Рисунок \ (\ PageIndex {1} \): Еда дает энергию для таких действий, как полет. : Колибри нужна энергия для поддержания продолжительных периодов полета. Колибри получает энергию, принимая пищу и превращая питательные вещества в энергию с помощью ряда биохимических реакций. Летательные мышцы птиц чрезвычайно эффективны в производстве энергии.
Колибри получает энергию, принимая пищу и превращая питательные вещества в энергию с помощью ряда биохимических реакций. Летательные мышцы птиц чрезвычайно эффективны в производстве энергии.Метаболизм, клеточный — Биологическая энциклопедия
Автор фотографии: Luk Cox
Клеточный метаболизм — это сумма всех биохимических реакций.
происходит внутри клетки.Он включает в себя все реакции, связанные с
разлагая молекулы пищи, синтезируя макромолекулы необходимы клетке, а также в производстве небольших молекул-предшественников, таких как
немного аминокислоты , для сотовых нужд. Он также включает все реакции с участием электронов.
перенос (окислительно-восстановительные или окислительно-восстановительные реакции). Обмен веществ занимает
место в последовательности биохимических реакций, называемых путями.
Метаболические пути
Метаболические пути могут быть простыми линейными последовательностями нескольких реакций или
они могут быть сильно разветвленными, при этом реакции сходятся или расходятся
от центральной главной дороги. Они могут быть циклическими, с предшественником
ранняя реакция регенерируется в конце пути (например, Цикл Кребса из аэробный дыхание, или цикл фотосинтеза Кальвина).Некоторые пути служат
несколько целей. Например, цикл Кребса наиболее известен своей ролью
в окисляющих сахарах и других органический молекулы для обеспечения аденозинтрифосфата ( АТФ ) для клетки, но он также используется в качестве источника молекул-предшественников для
клеточный биосинтез. Ясно, что эволюция неоднократно использовала существующие
метаболические пути для обеспечения новых функций.
Все биохимические реакции проходят катализированный к белки называется ферменты ; для большинства реакций есть один фермент, который катализирует только
реакция.Ферменты могут точно регулироваться клеткой, обеспечивая
высокая степень контроля клеточного метаболизма. Активность ферментов
часто чувствительны к количеству определенных молекул в клетке. За
например, ферменты, участвующие в производстве АТФ, часто ингибируются АТФ;
когда в клетке достаточно этого метаболит , следовательно, пути, которые его производят, отключены, тем самым
предотвращение бесполезных реакций.В качестве альтернативы те же самые ферменты могут быть
сильно активируется предшественником АТФ, аденозиндифосфатом ( ADP ), уровень которого повышается, когда клетка выполняет работу и нуждается в
быстрое образование АТФ. Этот образец регуляции молекулами, которые
предшественники или продукты пути распространены в клеточном
метаболизм.
Этот образец регуляции молекулами, которые
предшественники или продукты пути распространены в клеточном
метаболизм.
Анаболизм и катаболизм
Метаболизм делится на две большие категории.Катаболизм, или деградация молекул, как правило, включает удаление электронов из молекул (окисление) и обычно сопровождается высвобождением энергия. Анаболизм или синтез сложных молекул обычно включает: обогащает молекулы электронами (восстановление) и обычно требует клетка расходует энергию в виде АТФ. Реакции, дающие энергию, такие как большинство катаболических реакций, называются экзергоническими, а те, что требуют ввода энергии, например, большинство анаболический реакции, называются эндергоническими.
Основная функция анаболических путей — синтез четырех
классы макромолекул, необходимые клетке: полисахариды , липиды , нуклеиновые кислоты и белки. Хотя эти четыре категории
химически различны, все они синтезируются одним и тем же общим типом
реакция, конденсационный синтез отдельных малых субъединиц (мономеров)
в макромолекулы (полимеры).В реакции конденсации водород
атом удален из одного мономер , а гидроксил группа из другой, образуя воду. Между двумя
мономеры, из которых была удалена вода:
Хотя эти четыре категории
химически различны, все они синтезируются одним и тем же общим типом
реакция, конденсационный синтез отдельных малых субъединиц (мономеров)
в макромолекулы (полимеры).В реакции конденсации водород
атом удален из одного мономер , а гидроксил группа из другой, образуя воду. Между двумя
мономеры, из которых была удалена вода:
—А — ОН + Н — А— → Н 2 О + —А — А—
Например, нуклеиновые кислоты, такие как ДНК и РНК, синтезируются из их мономеры, нуклеотиды , путем конденсационного синтеза.Полисахариды и белки производятся в аналогичным образом из их мономеров, сахаров и аминокислот, соответственно. Липиды, четвертый класс макромолекул, несколько разные. В отличие от других макромолекул, которые
Сканирующая электронная микрофотография эпителиальной клетки кишечника. показаны две лизомы, содержащие пищеварительные ферменты, разрушающие
поврежденные молекулы. Одна категория метаболизма — катаболизм,
деградация молекул.
показаны две лизомы, содержащие пищеварительные ферменты, разрушающие
поврежденные молекулы. Одна категория метаболизма — катаболизм,
деградация молекул.
Обратный конденсационный синтез гидролиз , в котором молекула воды присоединена к связи между двумя мономерами, его разрушение и разделение мономеров.Один из водородов из воды присоединяется к одному из мономеров, а оставшийся гидроксил к другому прикреплен:
ЧАС 2 О + -А-А- → -А-ОН + H — A—
Например, нуклеиновые кислоты разлагаются до мономеров, когда вода
вставлен между отдельными нуклеотидными мономерами, разрывая связь
что присоединяется к ним.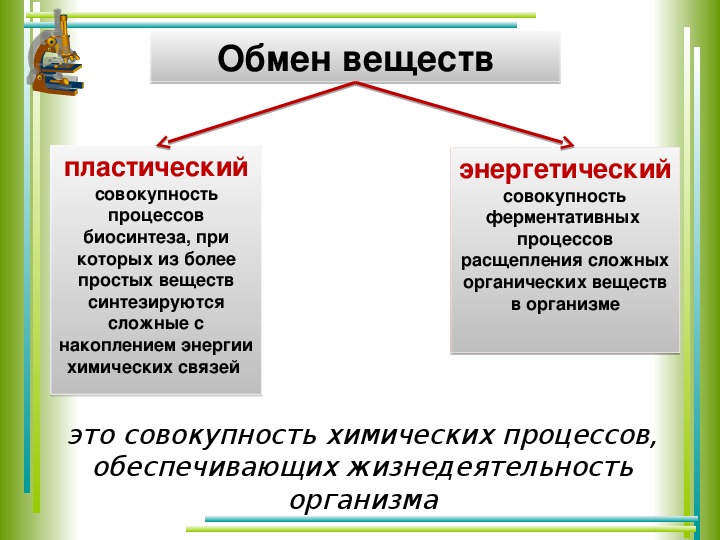 Реакции гидролиза — это разновидность катаболической реакции,
хотя они обычно не производят напрямую АТФ; они, однако,
продуцируют мономеры, которые часто далее катаболизируются с образованием АТФ.
Реакции гидролиза — это разновидность катаболической реакции,
хотя они обычно не производят напрямую АТФ; они, однако,
продуцируют мономеры, которые часто далее катаболизируются с образованием АТФ.
Оборот
Обмен веществ — это динамический процесс. Клетка постоянно деградирует и синтезирующие молекулы. В целом катаболические пути обеспечивают энергия в виде АТФ, которая используется для управления анаболическими процессами. Это необходимо, поскольку для протекания эндергонических реакций требуется вклад энергии, который они получают от АТФ. Это достигается соединение эндергонической реакции с гидролизом АТФ до АДФ и неорганический фосфат, экзэргоническая реакция.Пока количество энергии меньше, чем количество, высвобождаемое при гидролизе АТФ, связанная реакции будут продолжаться.
Динамический характер метаболизма приводит к постоянной деградации и
восстановление большинства ячеистых материалов. Например, белки существуют в
ячейку в течение относительно короткого времени, от минут до недель, причем большинство
белки, средняя продолжительность жизни которых составляет несколько дней. Структурные белки
обычно служат дольше, чем ферменты, но они тоже в конечном итоге разлагаются
и синтезировал заново.Таким же образом переворачиваются и другие ячеистые материалы.
аналогичным образом. Этот постоянный оборот ячеистых материалов сохраняет
клетка в хорошем состоянии. Молекулы, которые могли быть повреждены в результате
например, будучи частично окисленным, рано или поздно разрушится и
заменены.
Например, белки существуют в
ячейку в течение относительно короткого времени, от минут до недель, причем большинство
белки, средняя продолжительность жизни которых составляет несколько дней. Структурные белки
обычно служат дольше, чем ферменты, но они тоже в конечном итоге разлагаются
и синтезировал заново.Таким же образом переворачиваются и другие ячеистые материалы.
аналогичным образом. Этот постоянный оборот ячеистых материалов сохраняет
клетка в хорошем состоянии. Молекулы, которые могли быть повреждены в результате
например, будучи частично окисленным, рано или поздно разрушится и
заменены.
Клеточный метаболизм — это самый фундаментальный уровень, на котором динамический
свойства жизни начинают проявляться. Сложные взаимодействия разнообразных
пути, их регулирование и их организация демонстрируют
изысканная утонченность биохимии жизни.Все происходящие процессы
внутри отдельных организмов можно проследить пути клеточного
метаболизм.
Библиография
Хинкл П.С. и Р.Э. Маккарти. «Как клетки производят АТФ». Scientific American 238 (март 1978 г.): 104–123.
Страйер, Люберт. Биохимия. Нью-Йорк: В. Х. Фриман и компания, 1995.
Другие статьи, которые могут вам понравиться:
Метаболизм во времени и пространстве — изучение границ биологии развития
Введение
Истоки биологии развития настолько прочно укоренились в исследованиях метаболизма, что к 1931 году книга Джозефа Нидхема Химическая эмбриология превысила 2000 страниц (Needham, 1931 ).Однако это исследование было в значительной степени забыто, когда биологи развития обратились к более геноцентрическому подходу. Многие из вопросов вековой давности, поднятых Нидхэмом, занимали видное место на Симпозиуме EMBO / EMBL «Метаболизм во времени и пространстве», который возвещал о возвращении этой давно забытой основы биологии развития.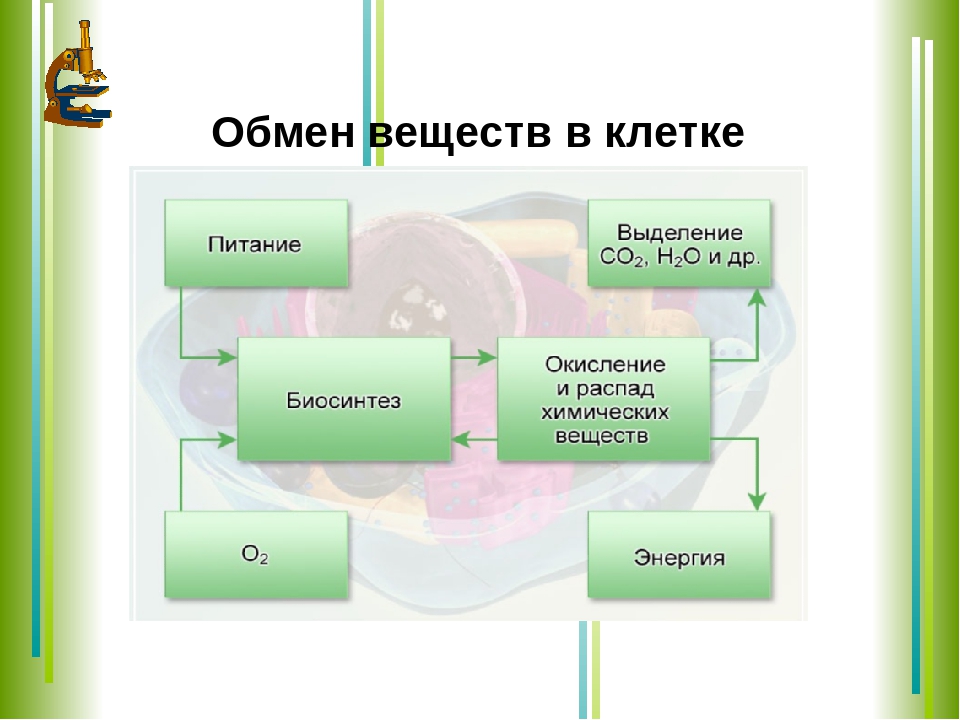 В течение трех дней в Гейдельберге (Германия) встречались самые разные ученые, чтобы обсудить исследования метаболизма в организмах, от бактерий до людей; независимо от системы, возникло несколько основных тем.Метаболизм должен адаптироваться к энергетическим потребностям пролиферации клеток, роста и развития тканей, и в результате скорость и направление метаболического потока претерпят драматические изменения в течение жизни любого организма. Однако метаболизм не просто реагирует на сигналы развития; скорее, отдельные метаболиты могут резко влиять на рост, дифференциацию и время событий жизненного цикла, таких как начало полового созревания. Наконец, организмы используют метаболические сенсорные механизмы, которые контролируют решения клеточной судьбы и развитие в зависимости от наличия питательных веществ.Эти общие темы способствовали чувству единства среди участников и подчеркнули важность этого типа междисциплинарных встреч.
В течение трех дней в Гейдельберге (Германия) встречались самые разные ученые, чтобы обсудить исследования метаболизма в организмах, от бактерий до людей; независимо от системы, возникло несколько основных тем.Метаболизм должен адаптироваться к энергетическим потребностям пролиферации клеток, роста и развития тканей, и в результате скорость и направление метаболического потока претерпят драматические изменения в течение жизни любого организма. Однако метаболизм не просто реагирует на сигналы развития; скорее, отдельные метаболиты могут резко влиять на рост, дифференциацию и время событий жизненного цикла, таких как начало полового созревания. Наконец, организмы используют метаболические сенсорные механизмы, которые контролируют решения клеточной судьбы и развитие в зависимости от наличия питательных веществ.Эти общие темы способствовали чувству единства среди участников и подчеркнули важность этого типа междисциплинарных встреч.
Механизмы определения питательных веществ
Рост и развитие — это ресурсоемкие процессы, которые зависят от доступа к адекватным запасам питательных веществ.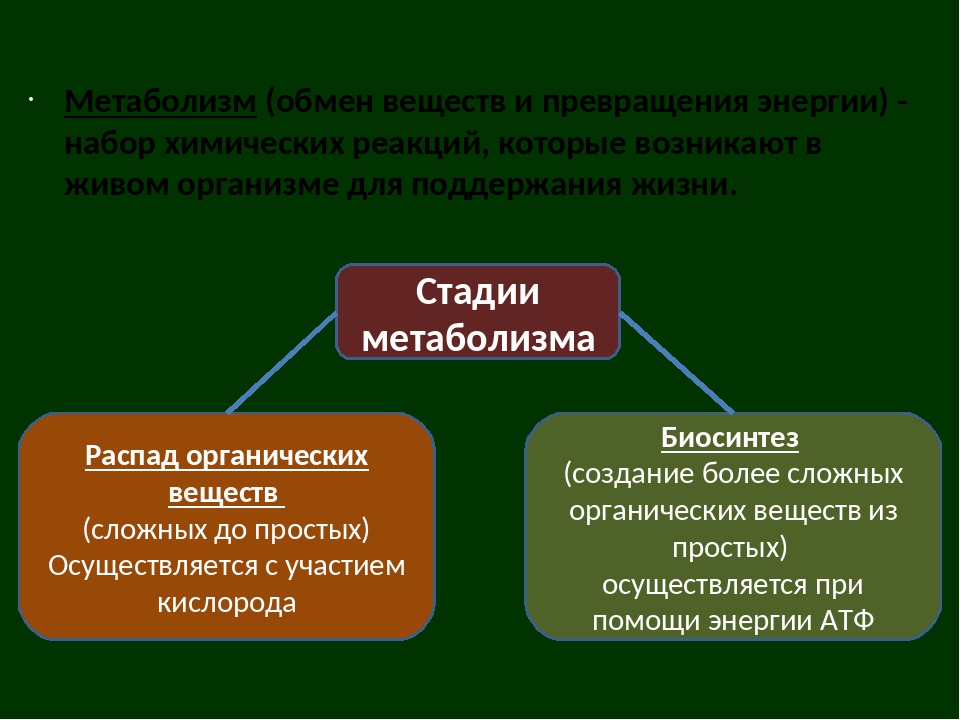 В результате решения о развитии часто контролируются датчиками питательных веществ, которые гарантируют, что растущие организмы имеют достаточные метаболические запасы. Эта взаимосвязь между питанием и ростом проиллюстрирована на примере бактерий, где деление клеток тесно связано с метаболическим потоком.В связи с этим Уве Зауэр (ETH Zurich, Швейцария) описал, как его лаборатория использовала метаболомику в реальном времени для определения того, как голодные по углероду Escherichia coli реагируют на импульсы глюкозы (Link et al., 2015). Изучая изменения в метаболическом потоке с 10-секундными интервалами, лаборатория Зауэра не только наблюдала, что глюкоза быстро метаболизируется и направляется в биосинтетические пути, но также раскрыла механизм, с помощью которого клетки, лишенные углерода, решают делиться под воздействием глюкозы.
В результате решения о развитии часто контролируются датчиками питательных веществ, которые гарантируют, что растущие организмы имеют достаточные метаболические запасы. Эта взаимосвязь между питанием и ростом проиллюстрирована на примере бактерий, где деление клеток тесно связано с метаболическим потоком.В связи с этим Уве Зауэр (ETH Zurich, Швейцария) описал, как его лаборатория использовала метаболомику в реальном времени для определения того, как голодные по углероду Escherichia coli реагируют на импульсы глюкозы (Link et al., 2015). Изучая изменения в метаболическом потоке с 10-секундными интервалами, лаборатория Зауэра не только наблюдала, что глюкоза быстро метаболизируется и направляется в биосинтетические пути, но также раскрыла механизм, с помощью которого клетки, лишенные углерода, решают делиться под воздействием глюкозы.
Точно так же, как размножение бактерий зависит от способности точно измерять гликолитический поток, рост эукариот также зависит от способности контролировать доступность питательных веществ. Классический пример этой метаболической необходимости был предоставлен Элисон Смит (Центр Джона Иннеса, Норвич, Великобритания), которая объяснила, как растения накапливают фотосинтетический крахмал в течение дня и используют его ночью. Смит и его коллеги определили, что Arabidopsis устанавливает скорость ночного использования крахмала на основе содержания крахмала в листьях и циркадных часов, которые предсказывают продолжительность ночи, тем самым гарантируя, что запасы крахмала сохранятся до рассвета (Scialdone et al., 2013). Их модель также показывает, что растения способны количественно определять крахмал, и, работая над выяснением этого механизма, лаборатория Смита обнаружила белок Arabidopsis , ESV1, который влияет на скорость оборота крахмала, физически взаимодействуя со структурой и контролируя ее. гранул крахмала (Feike et al., 2016).
Классический пример этой метаболической необходимости был предоставлен Элисон Смит (Центр Джона Иннеса, Норвич, Великобритания), которая объяснила, как растения накапливают фотосинтетический крахмал в течение дня и используют его ночью. Смит и его коллеги определили, что Arabidopsis устанавливает скорость ночного использования крахмала на основе содержания крахмала в листьях и циркадных часов, которые предсказывают продолжительность ночи, тем самым гарантируя, что запасы крахмала сохранятся до рассвета (Scialdone et al., 2013). Их модель также показывает, что растения способны количественно определять крахмал, и, работая над выяснением этого механизма, лаборатория Смита обнаружила белок Arabidopsis , ESV1, который влияет на скорость оборота крахмала, физически взаимодействуя со структурой и контролируя ее. гранул крахмала (Feike et al., 2016).
Развитие животных также должно реагировать на метаболические сигналы, и ряд лабораторий используют Drosophila в качестве модели для изучения восприятия питательных веществ. Карла Маргулис (Ludwig-Maximilians Universität, Мюнхен, Германия) объяснила, как ее лаборатория использовала ChIP, специфичный для определенного типа клеток, для идентификации транскрипционных мишеней сахарозависимого фактора транскрипции Mondo, который является гомологом Drosophila связывающего углеводный элемент белка. (ChREBP, также известный как MLXIPL). Их анализ показал, что участки связывания Mondo обогащены рядом с гликолитическими и липогенными генами, что позволяет предположить, что Mondo способствует синтезу липидов в ответ на сахар.Кроме того, лаборатория Маргулиса определила, что CrebA, член семейства факторов транскрипции Creb3L, активен у кормленных животных и регулирует гены, участвующие в секреторном аппарате.
Карла Маргулис (Ludwig-Maximilians Universität, Мюнхен, Германия) объяснила, как ее лаборатория использовала ChIP, специфичный для определенного типа клеток, для идентификации транскрипционных мишеней сахарозависимого фактора транскрипции Mondo, который является гомологом Drosophila связывающего углеводный элемент белка. (ChREBP, также известный как MLXIPL). Их анализ показал, что участки связывания Mondo обогащены рядом с гликолитическими и липогенными генами, что позволяет предположить, что Mondo способствует синтезу липидов в ответ на сахар.Кроме того, лаборатория Маргулиса определила, что CrebA, член семейства факторов транскрипции Creb3L, активен у кормленных животных и регулирует гены, участвующие в секреторном аппарате.
Пьер Леопольд (CNRS, Париж, Франция) и Норберт Перримон (Гарвардская медицинская школа, Кембридж, США) расширили обсуждение восприятия питательных веществ, описав сеть сигналов, которые контролируют секрецию инсулиноподобных пептидов Drosophila (Dilps).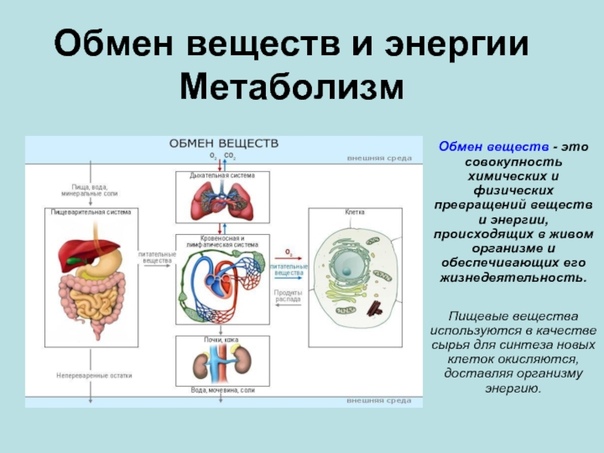 из инсулин-продуцирующих клеток (IPC).Подобно человеческому инсулину, Dilps являются ключевыми регуляторами роста и метаболизма, и, таким образом, муха оказалась мощной моделью для изучения механизмов, контролирующих секрецию инсулина. Любопытно, что IPC не несут прямой ответственности за обнаружение отдельных метаболитов; скорее, механизмы чувствительности к питательным веществам в периферических тканях дистанционно регулируют функцию IPC. Лаборатория Леопольда определила, что белки Eiger (TNFα) и Stunted (F1-F0 АТФ-синтаза) секретируются из жирового тела, чтобы контролировать высвобождение Dilp, хотя и для противоположных целей.Когда личинки попадают на диету с низким содержанием белка, снижение уровня аминокислот в жировом теле приводит к ингибированию Tor. Поскольку TNFα-превращающий фермент ТАСЕ обычно инактивируется Tor, диета с низким содержанием белка приводит к активации ТАСЕ и вызывает высвобождение Эйгера в гемолимфу. Затем Эйгер связывает G-белок-связанный рецептор (GPCR) Гриндельвальда, расположенный на поверхности IPC, что приводит к активации Jnk и ингибированию секреции Dilp (Agrawal et al.
из инсулин-продуцирующих клеток (IPC).Подобно человеческому инсулину, Dilps являются ключевыми регуляторами роста и метаболизма, и, таким образом, муха оказалась мощной моделью для изучения механизмов, контролирующих секрецию инсулина. Любопытно, что IPC не несут прямой ответственности за обнаружение отдельных метаболитов; скорее, механизмы чувствительности к питательным веществам в периферических тканях дистанционно регулируют функцию IPC. Лаборатория Леопольда определила, что белки Eiger (TNFα) и Stunted (F1-F0 АТФ-синтаза) секретируются из жирового тела, чтобы контролировать высвобождение Dilp, хотя и для противоположных целей.Когда личинки попадают на диету с низким содержанием белка, снижение уровня аминокислот в жировом теле приводит к ингибированию Tor. Поскольку TNFα-превращающий фермент ТАСЕ обычно инактивируется Tor, диета с низким содержанием белка приводит к активации ТАСЕ и вызывает высвобождение Эйгера в гемолимфу. Затем Эйгер связывает G-белок-связанный рецептор (GPCR) Гриндельвальда, расположенный на поверхности IPC, что приводит к активации Jnk и ингибированию секреции Dilp (Agrawal et al. , 2016). Напротив, диета с высоким содержанием белка активирует передачу сигналов Tor в жировом теле и вызывает высвобождение Stunted, который передает сигналы IPC через GPCR Methuselah и вызывает секрецию Dilp (Delanoue et al., 2016).
, 2016). Напротив, диета с высоким содержанием белка активирует передачу сигналов Tor в жировом теле и вызывает высвобождение Stunted, который передает сигналы IPC через GPCR Methuselah и вызывает секрецию Dilp (Delanoue et al., 2016).
Дополнительные исследования, проведенные лабораторией Perrimon, обнаружили дополнительные секретируемые факторы, которые контролируют передачу сигналов Dilp и системный метаболизм. Например, полученный из жирового тела белок ImpL2 секретируется в гемолимфу, чтобы изолировать и инактивировать циркулирующий Dilps (Kwon et al., 2015). Хроническая диета с высоким содержанием сахара вызывает высвобождение активина-β из средней кишки, что способствует передаче сигналов адипокинетического гормона в жировом теле и приводит к гипергликемии (Song et al., 2017). Примечательно, что Perrimon также описал свой анализ адипокина Drosophila Unpaired 2 (Upd2), который действует как функциональный ортолог лептина и высвобождается из жирового тела животных, которых кормят, чтобы индуцировать секрецию Dilp из IPC (Rajan and Perrimon, 2012).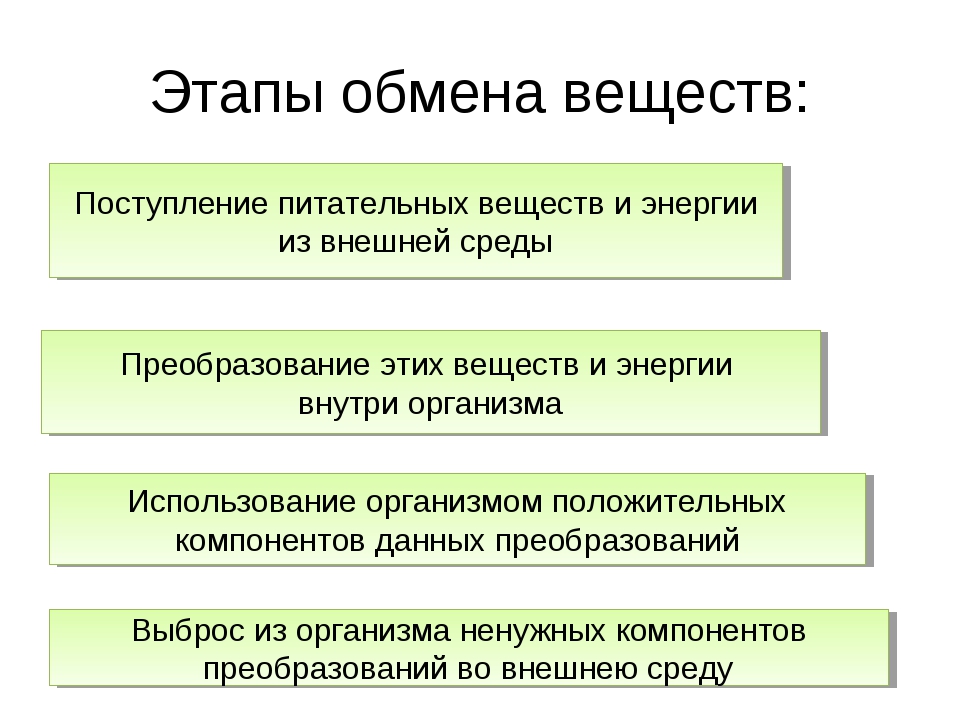 Перримон и его коллеги продемонстрировали, что регулируемый питательными веществами нетрадиционный путь секреции контролирует выпуск Upd2. Они также демонстрируют, что ключевой белок, участвующий в этом секреторном аппарате, фосфорилируется в ответ на голодание, что служит для ингибирования секреции Upd2. Поскольку этот механизм также способен регулировать секрецию человеческого лептина из жирового тела Drosophila , результаты лаборатории Perrimon предполагают, что консервативный механизм контролирует секрецию адипокина.
Перримон и его коллеги продемонстрировали, что регулируемый питательными веществами нетрадиционный путь секреции контролирует выпуск Upd2. Они также демонстрируют, что ключевой белок, участвующий в этом секреторном аппарате, фосфорилируется в ответ на голодание, что служит для ингибирования секреции Upd2. Поскольку этот механизм также способен регулировать секрецию человеческого лептина из жирового тела Drosophila , результаты лаборатории Perrimon предполагают, что консервативный механизм контролирует секрецию адипокина.
Наконец, Андреа Бранд (The Gurdon Institute, Кембридж, Великобритания) описала, как покоящиеся нервные стволовые клетки (NSC) реактивируются с помощью передачи сигналов инсулина во время развития личинок Drosophila . Предыдущие исследования в лаборатории Бранда показали, что кормление личинок индуцирует секрецию глиальных клеток гематоэнцефалического барьера (ГЭБ) Dilp6 (Ilp6), который запускает личиночные НСК для повторного входа в клеточный цикл (Chell and Brand, 2010).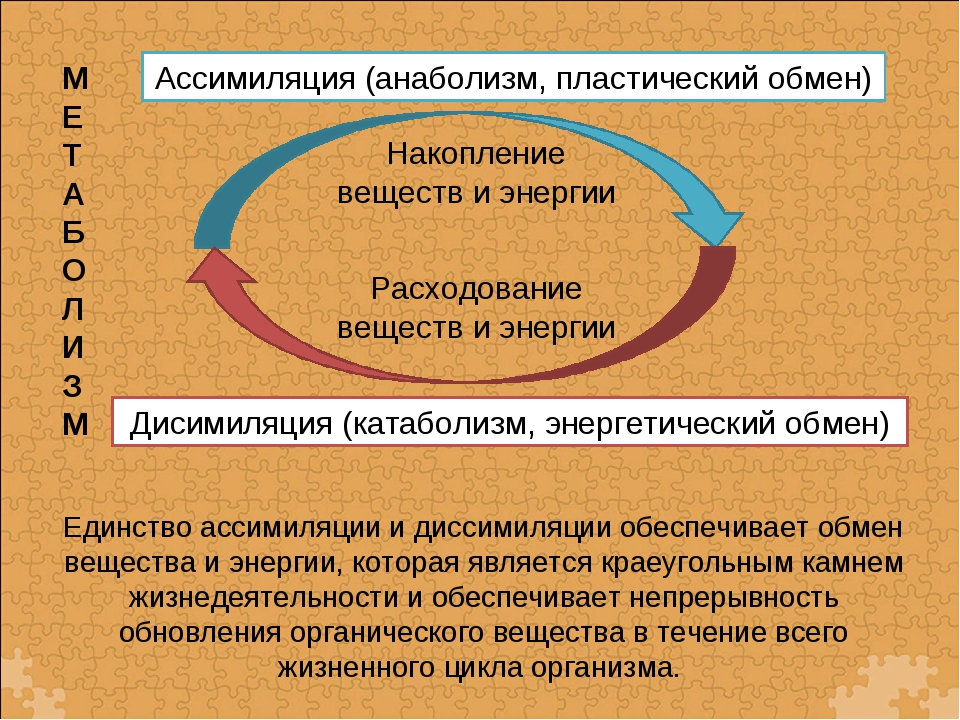 Ее лаборатория теперь определила, что передача сигналов Dilp6 от глии BBB требует белков щелевого соединения Inx1 (Ogre) и Inx2, которые позволяют зависимым от питательных веществ колебаниям кальция синхронизировать активность этих клеток (Spéder and Brand, 2014).Интересно, что секреция инсулина β-клетками поджелудочной железы человека также координируется передачей сигналов кальция и зависит от щелевых контактов, что указывает на сходство между системной передачей сигналов инсулина (от поджелудочной железы человека) и локальной передачей сигналов (в мозге Drosophila ).
Ее лаборатория теперь определила, что передача сигналов Dilp6 от глии BBB требует белков щелевого соединения Inx1 (Ogre) и Inx2, которые позволяют зависимым от питательных веществ колебаниям кальция синхронизировать активность этих клеток (Spéder and Brand, 2014).Интересно, что секреция инсулина β-клетками поджелудочной железы человека также координируется передачей сигналов кальция и зависит от щелевых контактов, что указывает на сходство между системной передачей сигналов инсулина (от поджелудочной железы человека) и локальной передачей сигналов (в мозге Drosophila ).
Пластичность метаболизма и развития
Клеточный метаболизм in vivo должен адаптироваться к изменениям качества и количества питательных веществ. Однако большинство метаболических экспериментов проводится с использованием стандартизированных условий роста, и в результате механизмы, регулирующие метаболическую пластичность, остаются плохо изученными.Чтобы понять, как клетки адаптируются к изменениям в доступности питательных веществ, Вильгельм Палм (Memorial Sloan Kettering, Нью-Йорк, США) изучил, как клетки пролиферируют в отсутствие свободных аминокислот (АК). Как объяснил Палм, когда количество свободных АК ограничивается, клетки используют макропиноцитоз для поглощения внеклеточных белков в качестве альтернативного источника АК. Однако, если свободные АК становятся доступными, активация mTor, индуцированная АК, будет ингибировать лизосомный катаболизм макропиноцитозированных белков. Следовательно, активация mTor обеспечивает метаболический переключатель, который обеспечивает непрерывный рост, несмотря на колебания в доступности АК.
Как объяснил Палм, когда количество свободных АК ограничивается, клетки используют макропиноцитоз для поглощения внеклеточных белков в качестве альтернативного источника АК. Однако, если свободные АК становятся доступными, активация mTor, индуцированная АК, будет ингибировать лизосомный катаболизм макропиноцитозированных белков. Следовательно, активация mTor обеспечивает метаболический переключатель, который обеспечивает непрерывный рост, несмотря на колебания в доступности АК.
Подобно клеточным исследованиям, описанным Палмом, метаболизм животных должен адаптироваться к различным источникам питательных веществ. Мэриан Уолхаут (Медицинская школа Массачусетского университета, Вустер, США) подчеркнула этот факт, описав, как C. elegans может перестраивать все метаболические пути в ответ на диетические сигналы. Лаборатория Уолхаута обнаружила, что личинок C. elegans , выращенных на рационе Comamonas DA1877, демонстрируют ускоренный рост личинок, сниженную плодовитость и сокращенную продолжительность жизни по сравнению с личинками, выращенными на стандартном рационе E.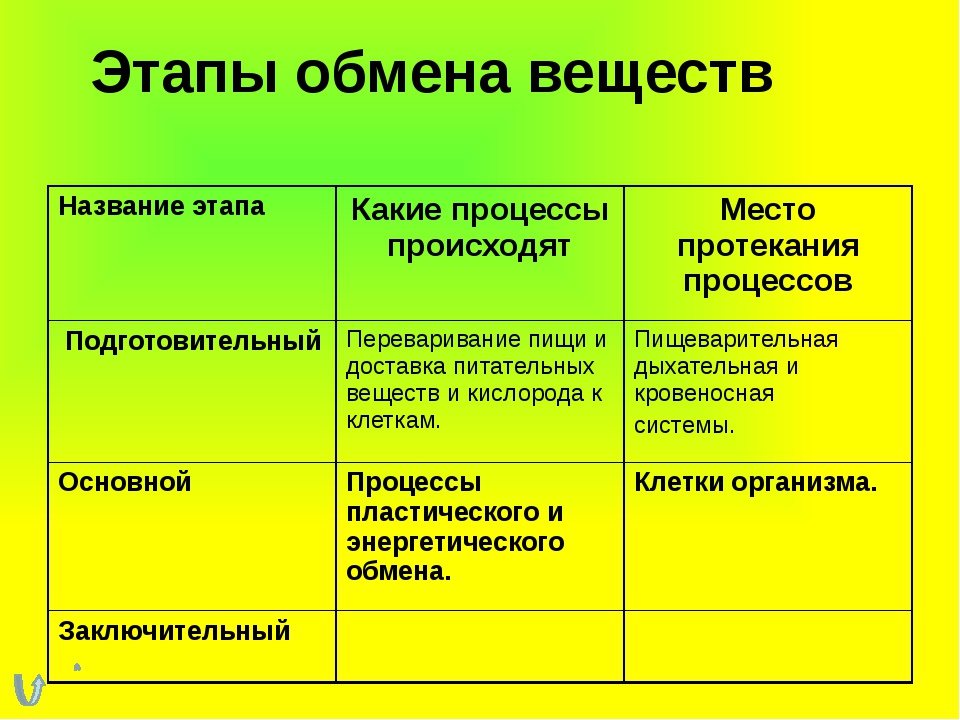 coli OP50 (MacNeil et al., 2013). Используя подход межвидовой системной биологии, они продемонстрировали, что эти зависимые от диеты фенотипы происходят из витамина B12, которого много в DA1877, но ограничено в диете OP50 (Watson et al., 2014). Кроме того, витамин B12 необходим для расщепления токсичного метаболита пропионата, и лаборатория Walhout определила, что личинки, выращенные на диете OP50, должны транскрипционно активировать витаминнезависимый путь разложения этого соединения (Watson et al., 2016). Кристиан Рёдельспергер (Институт биологии развития Макса Планка, Тюбинген, Германия) также подчеркнул пластичность развития нематод, описав, как род Pristionchus идеально подходит для изучения решений, принимаемых в зависимости от диеты в истории жизни. В ходе развития Pristionchus способны как формировать устойчивую к стрессу личинку дауэра, так и развивать хищные крючки в ответ на сигналы окружающей среды. Поскольку оба эти решения в отношении развития контролируются внутренне синтезируемыми феромонами, Родельспергер использует подход системной биологии, чтобы определить, как биосинтез феромонов изменяется в ответ на метаболический состав различных диет.
coli OP50 (MacNeil et al., 2013). Используя подход межвидовой системной биологии, они продемонстрировали, что эти зависимые от диеты фенотипы происходят из витамина B12, которого много в DA1877, но ограничено в диете OP50 (Watson et al., 2014). Кроме того, витамин B12 необходим для расщепления токсичного метаболита пропионата, и лаборатория Walhout определила, что личинки, выращенные на диете OP50, должны транскрипционно активировать витаминнезависимый путь разложения этого соединения (Watson et al., 2016). Кристиан Рёдельспергер (Институт биологии развития Макса Планка, Тюбинген, Германия) также подчеркнул пластичность развития нематод, описав, как род Pristionchus идеально подходит для изучения решений, принимаемых в зависимости от диеты в истории жизни. В ходе развития Pristionchus способны как формировать устойчивую к стрессу личинку дауэра, так и развивать хищные крючки в ответ на сигналы окружающей среды. Поскольку оба эти решения в отношении развития контролируются внутренне синтезируемыми феромонами, Родельспергер использует подход системной биологии, чтобы определить, как биосинтез феромонов изменяется в ответ на метаболический состав различных диет.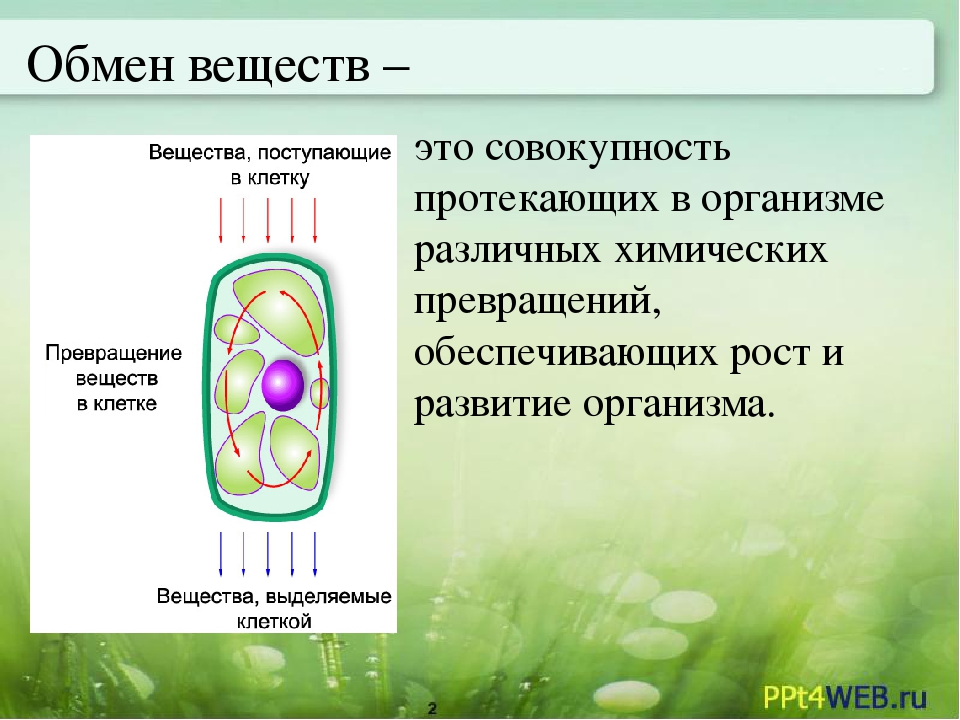
Важность использования природного разнообразия для изучения метаболической пластичности была также подчеркнута Тадаси Уэмура (Университет Киото, Япония), чья лаборатория изучает, как различные виды Drosophila развиваются на диетах с низким или высоким соотношением белков: углеводов. Лаборатория Уэмуры обнаружила, что личинки универсальных видов, таких как Drosophila melanogaster , могут расти на любой диете, тогда как специализированные виды не могут развиваться на среде с высоким содержанием углеводов.Метаболомное сравнение этих видов показало, что универсалы демонстрируют гомеостаз на богатой углеводами диете по сравнению со специалистами, и подразумевает, что Drosophila, гомолог активина , Dawdle, является регулятором этих адаптивных метаболических ответов.
Ирен Мигель-Алиага (MRC LMS, Imperial College, Лондон, Великобритания) расширила обсуждение пластичности, описав, насколько кишечник самок дрозофилы значительно больше, чем у самцов. Поскольку кишечник является ключевым регулятором усвоения питательных веществ, эта особенность, зависящая от пола, существенно влияет на метаболизм и физиологию.Лаборатория Мигеля-Алиаги определила, что различия между мужским и женским кишечником являются результатом неканонического пути определения пола, который функционирует внутри кишечных стволовых клеток (ISC) (Hudry et al., 2016). Более того, ее лаборатория обнаружила, что маскулинизация ISC у взрослых самок уменьшает размер кишечника и предотвращает изменение размера, которое обычно происходит после спаривания, тем самым раскрывая новый механизм, который глубоко влияет на обработку питательных веществ и репродуктивную способность. Их текущая работа теперь расширяет изучение половых различий на другие типы кишечных клеток, характеризуя их с помощью датчиков метаболизма и сплайсинга, специфичных для каждого типа клеток.
Поскольку кишечник является ключевым регулятором усвоения питательных веществ, эта особенность, зависящая от пола, существенно влияет на метаболизм и физиологию.Лаборатория Мигеля-Алиаги определила, что различия между мужским и женским кишечником являются результатом неканонического пути определения пола, который функционирует внутри кишечных стволовых клеток (ISC) (Hudry et al., 2016). Более того, ее лаборатория обнаружила, что маскулинизация ISC у взрослых самок уменьшает размер кишечника и предотвращает изменение размера, которое обычно происходит после спаривания, тем самым раскрывая новый механизм, который глубоко влияет на обработку питательных веществ и репродуктивную способность. Их текущая работа теперь расширяет изучение половых различий на другие типы кишечных клеток, характеризуя их с помощью датчиков метаболизма и сплайсинга, специфичных для каждого типа клеток.
Экологические сигналы регулируют развитие растений и передачу сигналов ауксина
Любой растущий организм должен адаптироваться к стрессу окружающей среды, но растения демонстрируют исключительный уровень пластичности, поскольку, в отличие от животных, они неподвижны и должны адаптироваться к местному дефициту питательных веществ. В связи с этим Оттолин Лейзер (Кембриджский университет, Великобритания) описал, как низкие уровни нитратов в почве подавляют ветвление у Arabidopsis , косвенно регулируя транспорт ауксина. Ауксин вырабатывается растущими листьями на верхушке побега и транспортируется вниз по стеблю с помощью семейства PIN белков оттока ауксинов, которые локализуются на базальной мембране клеток.В нормальных условиях роста новые побеги образуются, когда ауксин перетекает из верхушки почек в основной стебель. Лаборатория Лейзера обнаружила, что гормон стриголактон позволяет растениям настраивать свое ветвление в ответ на ограниченную доступность питательных веществ, индуцируя эндоцитоз PIN1 и PIN7 в стеблях, уменьшая поток ауксина и ингибируя активацию почек (de Jong et al., 2014).
В связи с этим Оттолин Лейзер (Кембриджский университет, Великобритания) описал, как низкие уровни нитратов в почве подавляют ветвление у Arabidopsis , косвенно регулируя транспорт ауксина. Ауксин вырабатывается растущими листьями на верхушке побега и транспортируется вниз по стеблю с помощью семейства PIN белков оттока ауксинов, которые локализуются на базальной мембране клеток.В нормальных условиях роста новые побеги образуются, когда ауксин перетекает из верхушки почек в основной стебель. Лаборатория Лейзера обнаружила, что гормон стриголактон позволяет растениям настраивать свое ветвление в ответ на ограниченную доступность питательных веществ, индуцируя эндоцитоз PIN1 и PIN7 в стеблях, уменьшая поток ауксина и ингибируя активацию почек (de Jong et al., 2014).
Пластичность развития корня также контролируется PIN-зависимым направленным транспортом ауксина, который, как объяснил Бен Шерес (Университет Вагенингена, Нидерланды), контролирует как быстрые трофические реакции, так и долгосрочное поддержание зон развития корня. Используя Arabidopsis , лаборатория Scheres обнаружила, что факторы транскрипции PLETHORA (PLT) являются ключевыми детерминантами зон развития внутри корневой меристемы и экспрессируются только в клетках, которые испытывают длительное воздействие высоких уровней ауксина (Mähönen et al., 2014). В результате экспрессия PLT поддерживает долгосрочную идентичность зон в меристеме корня, даже если локальные концентрации ауксина испытывают краткосрочные колебания, например, во время гравитропного ответа, когда градиент ауксина временно перераспределяется в нижний участок корня. .Эти результаты вместе с исследованием, описанным Лейзером, предоставили элегантный снимок того, как внешние сигналы влияют на развитие растений, модулируя движение отдельной молекулы.
Используя Arabidopsis , лаборатория Scheres обнаружила, что факторы транскрипции PLETHORA (PLT) являются ключевыми детерминантами зон развития внутри корневой меристемы и экспрессируются только в клетках, которые испытывают длительное воздействие высоких уровней ауксина (Mähönen et al., 2014). В результате экспрессия PLT поддерживает долгосрочную идентичность зон в меристеме корня, даже если локальные концентрации ауксина испытывают краткосрочные колебания, например, во время гравитропного ответа, когда градиент ауксина временно перераспределяется в нижний участок корня. .Эти результаты вместе с исследованием, описанным Лейзером, предоставили элегантный снимок того, как внешние сигналы влияют на развитие растений, модулируя движение отдельной молекулы.
Метаболическая регуляция развития и клеточного цикла
Многие из выступавших подчеркивали тот факт, что метаболизм не просто пассивная мишень передачи сигналов фактора роста, а, скорее, непосредственно регулирует развитие.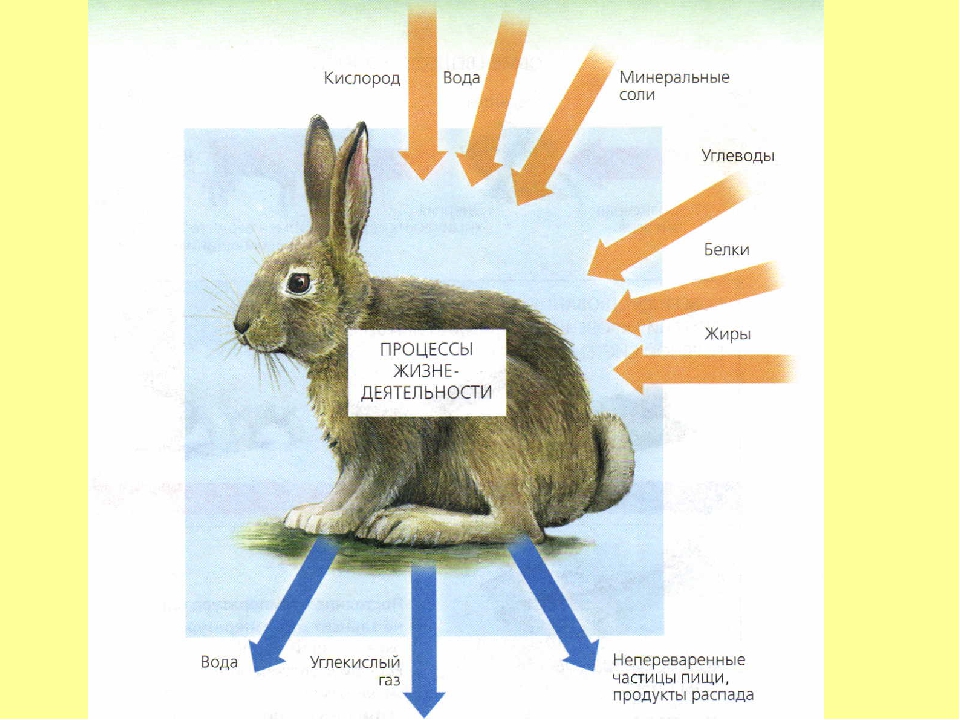 Этот момент был подчеркнут Питером Кармелиетом (VIB-KU Leuven, Бельгия), который описал, как метаболизм эндотелиальных клеток (ЭК) контролирует ангиогенез у млекопитающих.Предыдущая работа лаборатории Carmeliet продемонстрировала, что фермент PFKFB3 способствует прорастанию сосудов, индуцируя высокие уровни гликолиза ЭК (De Bock et al., 2013; Schoors et al., 2014). Тем не менее, несмотря на гликолитическую природу ЭК, лаборатория Кармелиета обнаружила, что CPT1A, который представляет собой фермент, ограничивающий скорость β-окисления жирных кислот, также необходим для образования кровеносных сосудов. Однако роль CPT1A в этом контексте не в продукции АТФ; скорее ЭК используют катаболизм жиров для синтеза de novo нуклеотидов (Schoors et al., 2015). Эти результаты вместе с другими неопубликованными исследованиями лаборатории Carmeliet подчеркивают, что метаболизм является важным регулятором ангиогенеза, и подчеркивают важность изучения того, как метаболические процессы напрямую влияют на рост тканей.
Этот момент был подчеркнут Питером Кармелиетом (VIB-KU Leuven, Бельгия), который описал, как метаболизм эндотелиальных клеток (ЭК) контролирует ангиогенез у млекопитающих.Предыдущая работа лаборатории Carmeliet продемонстрировала, что фермент PFKFB3 способствует прорастанию сосудов, индуцируя высокие уровни гликолиза ЭК (De Bock et al., 2013; Schoors et al., 2014). Тем не менее, несмотря на гликолитическую природу ЭК, лаборатория Кармелиета обнаружила, что CPT1A, который представляет собой фермент, ограничивающий скорость β-окисления жирных кислот, также необходим для образования кровеносных сосудов. Однако роль CPT1A в этом контексте не в продукции АТФ; скорее ЭК используют катаболизм жиров для синтеза de novo нуклеотидов (Schoors et al., 2015). Эти результаты вместе с другими неопубликованными исследованиями лаборатории Carmeliet подчеркивают, что метаболизм является важным регулятором ангиогенеза, и подчеркивают важность изучения того, как метаболические процессы напрямую влияют на рост тканей.
Маттиас Хайнеманн (Университет Гронингена, Нидерланды) дополнительно исследовал метаболические регуляторные функции у почкующихся дрожжей. Изучая метаболизм отдельных клеток, лаборатория Heinemann определила, что NAD (P) H и АТФ колеблются в противоположной фазе, один раз за клеточный цикл (Papagiannakis et al., 2017). Примечательно, что эти колебания продолжались и в клетках с остановленным клеточным циклом, указывая на то, что метаболические колебания автономны. В нормально делящихся клетках S-фаза последовательно начинается в то время, когда уровни NAD (P) H повышаются, и выход из митоза происходит на дне сигнала NAD (P) H, что указывает на связь метаболизма с клеточным циклом. Однако, когда клетки были переведены из среды с низким содержанием глюкозы в среду с высоким содержанием глюкозы, результирующее увеличение уровней NAD (P) H сбрасывает время клеточного цикла, так что выход из митоза задерживается для поддержания синхронизации с NAD (P). Колебания H.Эти результаты показывают, что метаболический поток играет ранее недооцененную роль в контроле клеточного цикла, и предполагают, что метаболические колебания могут серьезно влиять на пролиферацию клеток.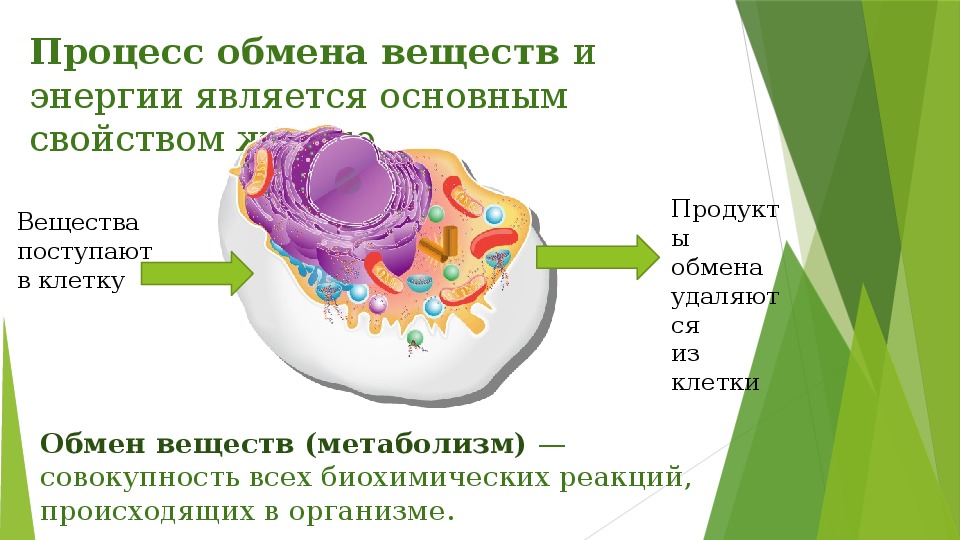
В центре внимания липиды
Липиды являются одними из самых распространенных метаболитов в эукариотической клетке и контролируют различные процессы развития. Анн-Клод Гэвин (EMBL, Гейдельберг, Германия) объяснила, как эти молекулы действуют в отдельных клеточных компартментах, влияя на структуру мембраны, метаболизм и передачу сигналов.Поскольку липиды гидрофобны, механизмы, которые перемещают отдельные виды липидов по цитозолю, также являются ключевыми регуляторами их активности. Путем очистки белков-переносчиков липидов из дрожжей лаборатория Гэвина определила, что оксистерин-связывающие белки Osh6 и Osh7 переносят фосфатидилсерин из эндоплазматического ретикулума в плазматическую мембрану (Maeda et al., 2013). Интересно, что эти семейства белков связаны с рядом заболеваний человека, что указывает на то, что сортировка липидов может глубоко влиять на метаболизм и физиологию.
Важность изучения липидного обмена была далее подчеркнута Алексом Гулдом (Институт Фрэнсиса Крика, Лондон, Великобритания), чья лаборатория изучила, как окислительный стресс влияет на рост личинок Drosophila . Лаборатория Гулда обнаружила, что, хотя гипоксия вызывает остановку клеточного цикла в большинстве тканей личинок, нейробласты продолжают делиться при низком давлении кислорода (Bailey et al., 2015). Это ограничение пролиферации нейробластов зависит от индуцированного гипоксией накопления липидных капель (LD) в глиальных клетках, которые образуются, когда полиненасыщенные жирные кислоты (PUFA) и другие жирные кислоты перераспределяются из плазматической мембраны в глиальные LD.Поскольку окислительный стресс вызывает перекисное окисление ПНЖК, которое впоследствии может повредить белки и другие макромолекулы, их результаты показывают, что образование ЛД защищает ПНЖК и ограничивает повреждение, вызванное активными формами кислорода как в глии, так и в соседних нейробластах.
Лаборатория Гулда обнаружила, что, хотя гипоксия вызывает остановку клеточного цикла в большинстве тканей личинок, нейробласты продолжают делиться при низком давлении кислорода (Bailey et al., 2015). Это ограничение пролиферации нейробластов зависит от индуцированного гипоксией накопления липидных капель (LD) в глиальных клетках, которые образуются, когда полиненасыщенные жирные кислоты (PUFA) и другие жирные кислоты перераспределяются из плазматической мембраны в глиальные LD.Поскольку окислительный стресс вызывает перекисное окисление ПНЖК, которое впоследствии может повредить белки и другие макромолекулы, их результаты показывают, что образование ЛД защищает ПНЖК и ограничивает повреждение, вызванное активными формами кислорода как в глии, так и в соседних нейробластах.
Наконец, Аурелио Телеман (German Cancer Research Center, Гейдельберг, Германия) описал, как стеариновая кислота (C18: 0) действует как сигнальная молекула во время развития Drosophila (Senyilmaz et al. , 2015).В лаборатории Teleman мутировали ген Drosophila Elovl6 ( Baldspot ), который кодирует элонгазу жирных кислот, необходимую для синтеза C18: 0. При характеристике этого мутанта они обнаружили, что животные, лишенные C18: 0, демонстрируют дефекты слияния митохондрий и гиперчувствительны к митотоксинам. Однако эти фенотипы не являются результатом дефекта структуры митохондриальной мембраны. Как в клетках Drosophila , так и в клетках человека C18: 0 регулирует убиквитилирование и активность митофузина, который контролирует слияние митохондрий.В клетках человека C18: 0 необходим для стеаролилирования и инактивации рецептора трансферрина (TFR1 или TFRC). В отсутствие C18: 0 TFR1 активирует JNK, что приводит к убиквитилированию и инактивации митофузина и фрагментации митохондрий. В целом, неожиданные результаты лаборатории Teleman подчеркивают, как кажущийся обычным липид может напрямую влиять на передачу сигналов, рост и метаболизм клеток.
, 2015).В лаборатории Teleman мутировали ген Drosophila Elovl6 ( Baldspot ), который кодирует элонгазу жирных кислот, необходимую для синтеза C18: 0. При характеристике этого мутанта они обнаружили, что животные, лишенные C18: 0, демонстрируют дефекты слияния митохондрий и гиперчувствительны к митотоксинам. Однако эти фенотипы не являются результатом дефекта структуры митохондриальной мембраны. Как в клетках Drosophila , так и в клетках человека C18: 0 регулирует убиквитилирование и активность митофузина, который контролирует слияние митохондрий.В клетках человека C18: 0 необходим для стеаролилирования и инактивации рецептора трансферрина (TFR1 или TFRC). В отсутствие C18: 0 TFR1 активирует JNK, что приводит к убиквитилированию и инактивации митофузина и фрагментации митохондрий. В целом, неожиданные результаты лаборатории Teleman подчеркивают, как кажущийся обычным липид может напрямую влиять на передачу сигналов, рост и метаболизм клеток.
Новые роли эффекта Варбурга
Почти столетие назад Отто Варбург заметил, что опухоли потребляют исключительно высокий уровень глюкозы и имеют тенденцию превращать пируват в лактат независимо от наличия кислорода. Это явление, известное как аэробный гликолиз или эффект Варбурга, идеально подходит для поддержки роста и биосинтеза. Джаред Раттер (Университет штата Юта, Солт-Лейк-Сити, США) подчеркнул важность эффекта Варбурга, описывая, как его лаборатория открыла молекулярную идентичность митохондриального переносчика пирувата (MPC). Этот консервативный белковый комплекс отвечает за транспортировку пирувата в митохондриальный матрикс и представляет собой ключевое метаболическое звено между гликолизом и окислительным фосфорилированием (Bricker et al., 2012). В соответствии с наблюдениями Варбурга лаборатория Раттера обнаружила, что многие виды рака демонстрируют снижение экспрессии MPC1, что приводит к усиленному гликолитическому состоянию и увеличению роста опухоли (Schell et al., 2014). Кроме того, Раттер также описал неопубликованные данные, которые показывают, что MPC способствует пролиферации стволовых клеток в кишечнике, тем самым делая MPC ключевым регулятором развития.
Это явление, известное как аэробный гликолиз или эффект Варбурга, идеально подходит для поддержки роста и биосинтеза. Джаред Раттер (Университет штата Юта, Солт-Лейк-Сити, США) подчеркнул важность эффекта Варбурга, описывая, как его лаборатория открыла молекулярную идентичность митохондриального переносчика пирувата (MPC). Этот консервативный белковый комплекс отвечает за транспортировку пирувата в митохондриальный матрикс и представляет собой ключевое метаболическое звено между гликолизом и окислительным фосфорилированием (Bricker et al., 2012). В соответствии с наблюдениями Варбурга лаборатория Раттера обнаружила, что многие виды рака демонстрируют снижение экспрессии MPC1, что приводит к усиленному гликолитическому состоянию и увеличению роста опухоли (Schell et al., 2014). Кроме того, Раттер также описал неопубликованные данные, которые показывают, что MPC способствует пролиферации стволовых клеток в кишечнике, тем самым делая MPC ключевым регулятором развития.
Результаты лаборатории Раттера были поддержаны в выступлениях Николь Прайор (лаборатория Aulehla, EMBL, Гейдельберг, Германия) и Оливье Пуркье (Гарвардская медицинская школа, Кембридж, США), которые описали градиент гликолиза от заднего к переднему в пресомитической области.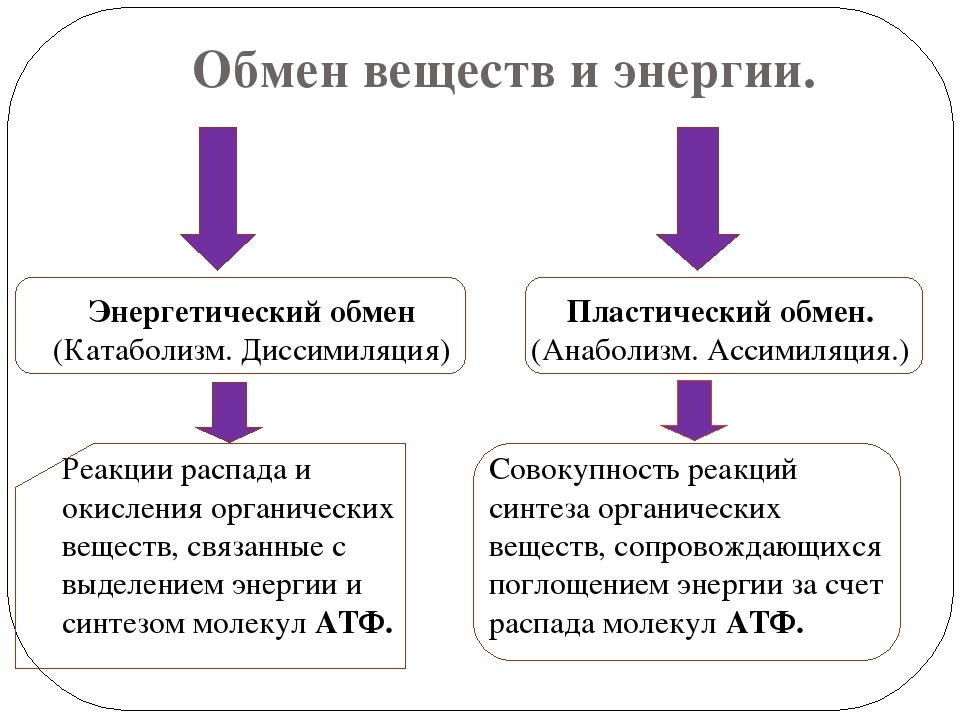 мезодерма (PSM) хвостовой почки позвоночных (Bulusu et al., 2017; Огинума и др., 2017). Используя комбинацию метаболомики, транскриптомики и сенсора FRET пирувата, их лаборатории обнаружили, что задний PSM демонстрирует повышенные уровни гликолитического потока и продукции лактата, предполагая, что он полагается на аэробный гликолиз для поддержания недифференцированного состояния. В соответствии с этой возможностью, обе группы определили, что ингибирование гликолитического потока в PSM приводит к аномалиям развития. Однако цель гликолиза в этой области остается неясной, поскольку неопубликованные данные из лаборатории Aulehla показали, что только минимальное количество глюкозы необходимо для поддержки формирования мезодермального паттерна.Как объяснил Прайор, следовые количества глюкозы, использованные в этом эксперименте, не влияют на накопление гликолитических метаболитов, предполагая, что аэробный гликолиз в PSM имеет неканоническую функцию. Один ключ к пониманию того, как аэробный гликолиз способствует развитию PSM независимо от биосинтеза, принадлежит группе Pourquié, которая продемонстрировала, что градиент передачи сигналов FGF / MAPK, присутствующий в этой ткани, регулирует транскрипцию нескольких гликолитических ферментов.
мезодерма (PSM) хвостовой почки позвоночных (Bulusu et al., 2017; Огинума и др., 2017). Используя комбинацию метаболомики, транскриптомики и сенсора FRET пирувата, их лаборатории обнаружили, что задний PSM демонстрирует повышенные уровни гликолитического потока и продукции лактата, предполагая, что он полагается на аэробный гликолиз для поддержания недифференцированного состояния. В соответствии с этой возможностью, обе группы определили, что ингибирование гликолитического потока в PSM приводит к аномалиям развития. Однако цель гликолиза в этой области остается неясной, поскольку неопубликованные данные из лаборатории Aulehla показали, что только минимальное количество глюкозы необходимо для поддержки формирования мезодермального паттерна.Как объяснил Прайор, следовые количества глюкозы, использованные в этом эксперименте, не влияют на накопление гликолитических метаболитов, предполагая, что аэробный гликолиз в PSM имеет неканоническую функцию. Один ключ к пониманию того, как аэробный гликолиз способствует развитию PSM независимо от биосинтеза, принадлежит группе Pourquié, которая продемонстрировала, что градиент передачи сигналов FGF / MAPK, присутствующий в этой ткани, регулирует транскрипцию нескольких гликолитических ферментов.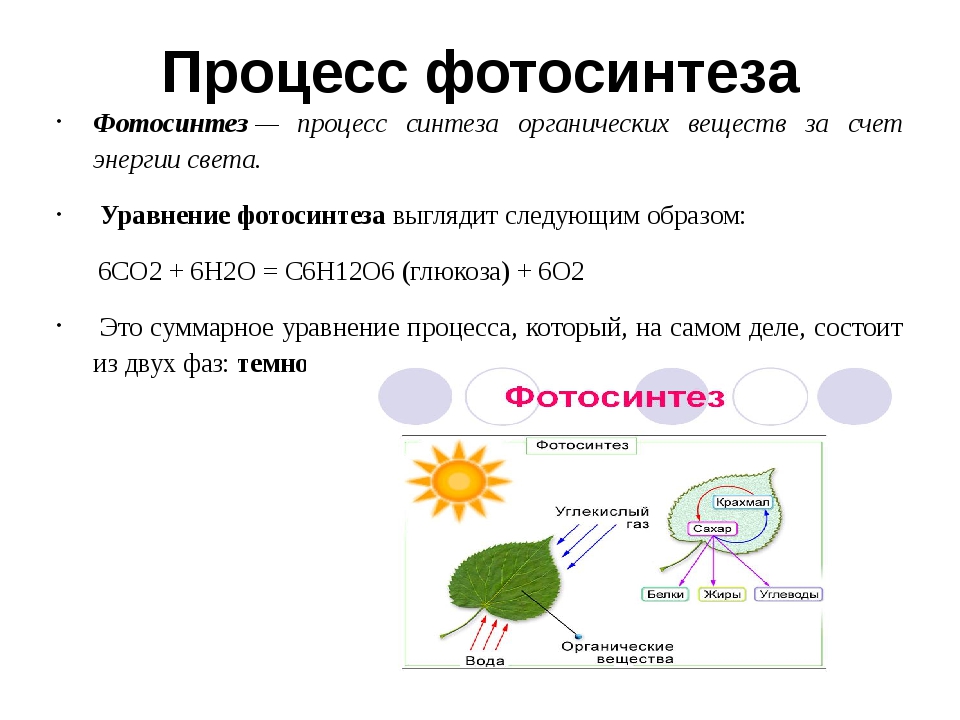 Интересно, что FGF также контролирует передачу сигналов Wnt в PSM, а ингибирование гликолиза в этой ткани фенокопирует потерю передачи сигналов Wnt в хвостовой зачатке.Следовательно, группа Pourquié проверила возможность того, что FGF влияет на передачу сигналов Wnt посредством гликолитического потока, и действительно обнаружила, что мишени Wnt подавляются после ингибирования гликолиза. Вместе эти данные подтверждают модель, в которой передача сигналов FGF устанавливает гликолитический градиент в развивающейся хвостовой почке, который, в свою очередь, необходим для поддержания передачи сигналов Wnt.
Интересно, что FGF также контролирует передачу сигналов Wnt в PSM, а ингибирование гликолиза в этой ткани фенокопирует потерю передачи сигналов Wnt в хвостовой зачатке.Следовательно, группа Pourquié проверила возможность того, что FGF влияет на передачу сигналов Wnt посредством гликолитического потока, и действительно обнаружила, что мишени Wnt подавляются после ингибирования гликолиза. Вместе эти данные подтверждают модель, в которой передача сигналов FGF устанавливает гликолитический градиент в развивающейся хвостовой почке, который, в свою очередь, необходим для поддержания передачи сигналов Wnt.
В дополнение к недавно обнаруженной взаимосвязи между аэробным гликолизом, FGF и передачей сигналов Wnt, Alena Krejci (Университет Южной Богемии, Ческе-Будейовице, Чешская Республика) сообщила, что сигнальный путь Notch также способен способствовать гликолитическому состоянию.Группа Крейчи обнаружила, что многие метаболические гены Drosophila содержат сайты связывания для Notch-специфичного супрессора фактора транскрипции лысого, и что короткий импульс активности Notch в клетках культуры ткани S2N вызывает длительное метаболическое ремоделирование в направлении эффекта Варбурга (Slaninova et al. ., 2016). Более того, экспрессия нескольких генов, участвующих в метаболизме глюкозы, регулируется Notch в крыловом диске, и Notch может использовать этот механизм для стимулирования роста этой развивающейся ткани.Принимая во внимание, что путь Notch активен в других контекстах, в которых клетки или ткани обнаруживают повышенные уровни гликолиза, эти наблюдения предполагают, что Notch может регулировать аэробный гликолиз в различных условиях.
., 2016). Более того, экспрессия нескольких генов, участвующих в метаболизме глюкозы, регулируется Notch в крыловом диске, и Notch может использовать этот механизм для стимулирования роста этой развивающейся ткани.Принимая во внимание, что путь Notch активен в других контекстах, в которых клетки или ткани обнаруживают повышенные уровни гликолиза, эти наблюдения предполагают, что Notch может регулировать аэробный гликолиз в различных условиях.
Роль эффекта Варбурга в развитии была дополнительно исследована Джейсоном Теннессеном (Университет Индианы, Блумингтон, США), чья лаборатория обнаружила, что личинок дрозофилы используют аэробный гликолиз для синтеза предполагаемого онкометаболита L-2-гидроксиглутарата (Li et al. ., 2017). Поскольку это соединение влияет на метилирование гистонов и ДНК, оно может действовать как метаболический сигнал во время развития личинок. При рассмотрении в контексте презентаций Прайора и Пуркье наблюдения лаборатории Теннессена подчеркивают, что аэробный гликолиз потенциально может влиять на развитие независимо от биосинтеза.
Метаболизм и иммунитет
Общая тема, которая возникла на симпозиуме, заключалась в том, что сходные метаболические механизмы контролируют решения о судьбе клеток как во время развития, так и во время иммунного ответа.Например, так же, как эффект Варбурга связан с опухолями и недифференцированными тканями, активированные эффекторные Т-клетки (T E ) полагаются на аэробный гликолиз, тогда как Т-клетки памяти (T M ), как правило, используют окислительный метаболизм. Группа Эрики Пирс (Институт иммунобиологии и эпигенетики Макса Планка, Фрайбург, Германия) определила, что эти различия в метаболизме Т-клеток зависят от структуры митохондрий (Buck et al., 2016). Как описал Пирс, более гликолитические клетки T E имеют точечные митохондрии, тогда как клетки T M демонстрируют митохондрии, которые слиты в сети.Поскольку эти структурные изменения предшествуют метаболическим изменениям, их наблюдения утверждают, что морфология митохондрий контролирует клеточное метаболическое состояние.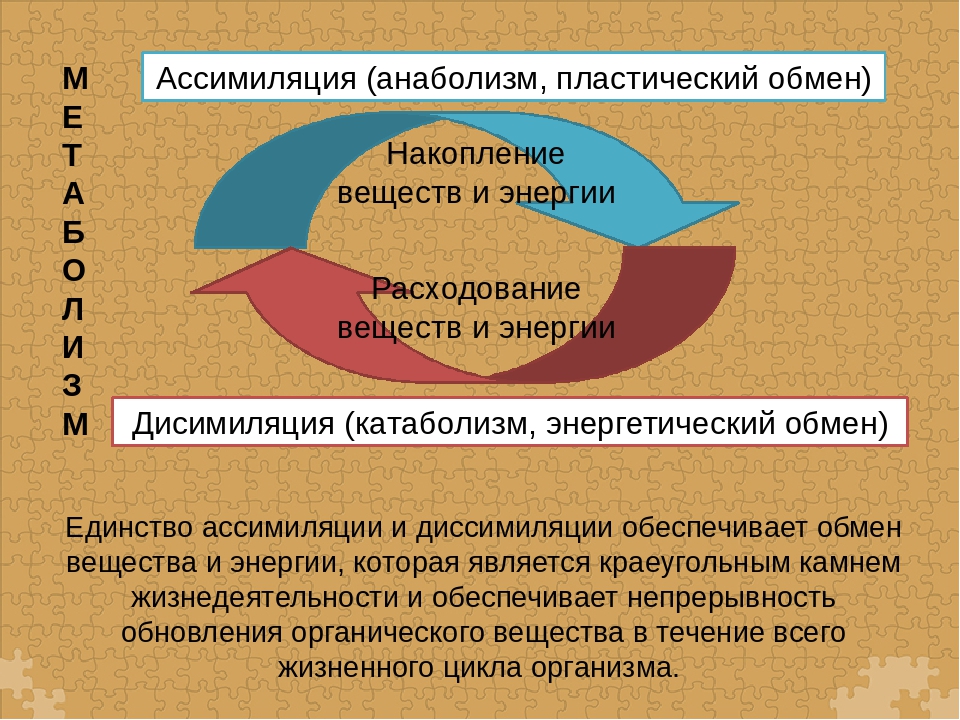 Действительно, лаборатория Пирса продемонстрировала, что белок внутренней митохондриальной мембраны Opa1 способствует слиянию митохондрий в клетках T M и индуцирует судьбу клеток T M даже в условиях культивирования, которые обычно программируют дифференцировку клеток T E . Кроме того, эти наблюдения предполагают, что препараты, способствующие слиянию митохондрий, могут создавать метаболически приспособленные Т-клетки и улучшать адаптивную клеточную иммунотерапию против опухолей.
Действительно, лаборатория Пирса продемонстрировала, что белок внутренней митохондриальной мембраны Opa1 способствует слиянию митохондрий в клетках T M и индуцирует судьбу клеток T M даже в условиях культивирования, которые обычно программируют дифференцировку клеток T E . Кроме того, эти наблюдения предполагают, что препараты, способствующие слиянию митохондрий, могут создавать метаболически приспособленные Т-клетки и улучшать адаптивную клеточную иммунотерапию против опухолей.
Как упоминал Пирс, взаимодействие между метаболизмом и иммунитетом представляет особый интерес в области рака. С целью дальнейшего изучения этой взаимосвязи Лиза Келли (Эдинбургский университет, Великобритания) и ее коллеги используют рыбок данио в качестве модели для выяснения метаболических сигналов, участвующих в привлечении клеток врожденного иммунитета во время раннего развития рака. Их данные свидетельствуют о том, что развивающиеся предопухолевые клетки демонстрируют изменения в митохондриальной динамике и связанных с ними метаболических процессах, которые изменяют дыхательную способность. Келли теперь использует репортерные линии трансгенных рыбок данио в сочетании с визуализацией в реальном времени, чтобы понять, как эти метаболические сигналы задействуют иммунные клетки.
Келли теперь использует репортерные линии трансгенных рыбок данио в сочетании с визуализацией в реальном времени, чтобы понять, как эти метаболические сигналы задействуют иммунные клетки.
Два сотрудника лаборатории Доминика Феррандона (Страсбургский университет, Франция) дополнительно изучили тему иммунных реакций, описав, как кишечник животных реагирует на острое воздействие токсина. Во-первых, Кэтрин Соча описала, как порообразующий бактериальный токсин гемолизин приводит к истончению кишечного эпителия у Drosophila , медоносных пчел и людей (Lee et al., 2016). Неожиданно это истончение является результатом вытеснения апикальной цитоплазмы из энтероцитов (кишечных клеток) в просвет кишечника, тем самым обеспечивая этим клеткам механизм для поддержания барьерной функции против бактериальной инвазии. Соча также объяснил, как эндотелий кишечника восстанавливается через несколько часов после воздействия токсина, и подробно рассказал о механизмах, лежащих в основе этого процесса.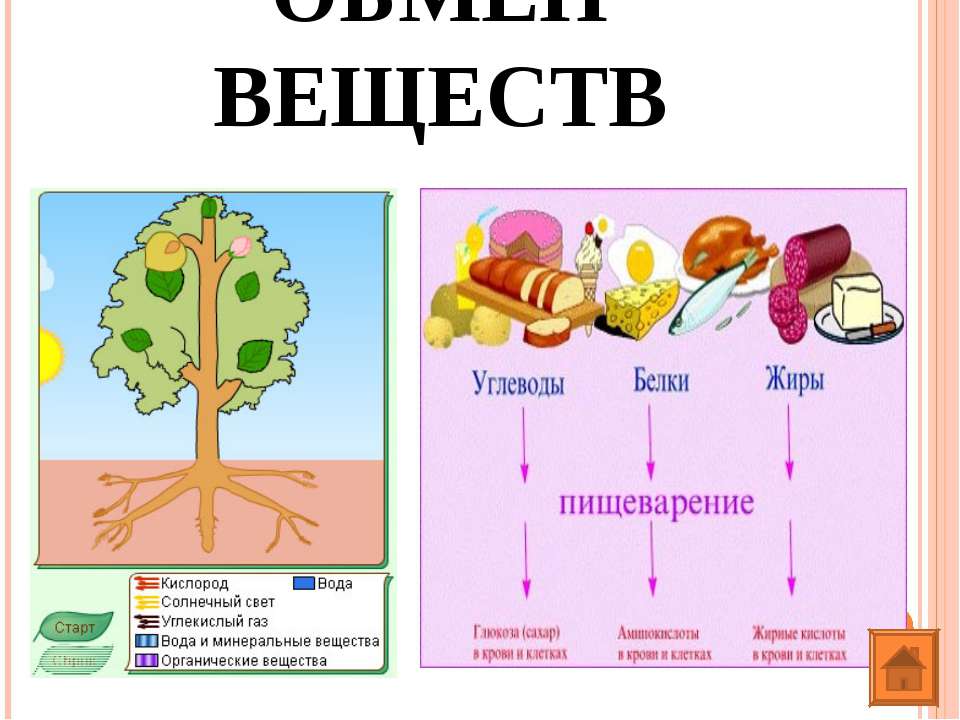 Во-вторых, Адриен Франше представил результаты, показывающие, что эпителий кишечника Drosophila реагирует на воздействие некоторых ксенобиотических веществ путем экструзии ЛД.Его наблюдения предполагают, что экструдированные ЛД защищают организм от пагубного действия ксенобиотиков и дополняют открытие лаборатории Гулда о том, что глиальные ЛД защищают нейробласты от окислительного повреждения.
Во-вторых, Адриен Франше представил результаты, показывающие, что эпителий кишечника Drosophila реагирует на воздействие некоторых ксенобиотических веществ путем экструзии ЛД.Его наблюдения предполагают, что экструдированные ЛД защищают организм от пагубного действия ксенобиотиков и дополняют открытие лаборатории Гулда о том, что глиальные ЛД защищают нейробласты от окислительного повреждения.
Метаболизм при лунном свете
Многие ферменты, участвующие в метаболизме, были впервые описаны почти столетие назад. Тем не менее, несмотря на десятилетия исследований, только недавно начали проявляться функции «лунного света» этих хорошо изученных белков. Неканоническая функция ферментов заметно фигурировала в презентации Матиаса Хенце (EMBL, Гейдельберг, Германия), чья лаборатория недавно продемонстрировала, что многие ферменты, участвующие в метаболизме центрального углерода, служат в качестве РНК-связывающих белков (названных enigmRBP) (Beckmann et al., 2015). Хотя функция большинства enigmRBPs остается плохо изученной, Hentze описал неопубликованные результаты, которые показывают, что белок рецептора аутофагии p62 (SQSTM1) связывает сводные РНК (vtRNAs). Это взаимодействие предотвращает индукцию аутофагии p62 — результат, который раскрывает новый основанный на РНК механизм регуляции аутофагии и предполагает, что сотни enigmRBPs, вероятно, будут выполнять важные и разнообразные функции.
Это взаимодействие предотвращает индукцию аутофагии p62 — результат, который раскрывает новый основанный на РНК механизм регуляции аутофагии и предполагает, что сотни enigmRBPs, вероятно, будут выполнять важные и разнообразные функции.
В то время как Хенце подчеркивал подрабатывающую функцию метаболических ферментов, Авинаш Патель (Институт молекулярной клеточной биологии и генетики им. Макса Планка, Дрезден, Германия) обнаружил, что даже АТФ играет неметаболическую роль (Patel et al., 2017). Патель и его коллеги обнаружили, что в физиологических условиях (2-8 мМ) АТФ способствует растворимости белка и подавляет образование агрегатов белка. Примечательно, что эта гидротропная функция не зависит от более канонической роли АТФ как носителя энергии и предполагает, что возрастное снижение концентрации АТФ может быть причиной усиленной патологической агрегации белков, наблюдаемой при нейродегенеративных заболеваниях. Хотя эти наблюдения интересны сами по себе, более широкая интерпретация результатов Пателя предполагает, что многие хорошо изученные метаболиты, вероятно, обладают еще не открытыми функциями.
Новые технологии и направления будущего
Главный вывод заключался в том, что развитие и метаболизм неразрывно связаны. Это важный момент для биологов, занимающихся развитием, которые часто рассматривают метаболические ферменты либо как гены домашнего хозяйства, либо как нижестоящие мишени каскадов передачи сигналов. Двигаясь вперед, наше сообщество должно предпринять согласованные усилия, чтобы понять, как отдельные метаболиты контролируют клеточную сигнализацию и развитие. Кроме того, мы должны выйти за рамки геномного и протеомного подходов и принять новые метаболомные технологии, чтобы понять, как распределение и количество метаболитов изменяются во время развития.Помня об этих целях, на встрече был отмечен ряд технологических достижений, которые значительно расширят метаболический инструментарий биологов-разработчиков. Например, Кай Джонссон (EPFL, Лозанна, Швейцария) описал флуоресцентные белковые сенсоры, которые позволяют измерять метаболиты в реальном времени, а Теодор Александров (EMBL, Гейдельберг, Германия) объяснил, как новые методы метаболомики могут визуализировать пространственное распределение метаболитов в развивающихся тканях.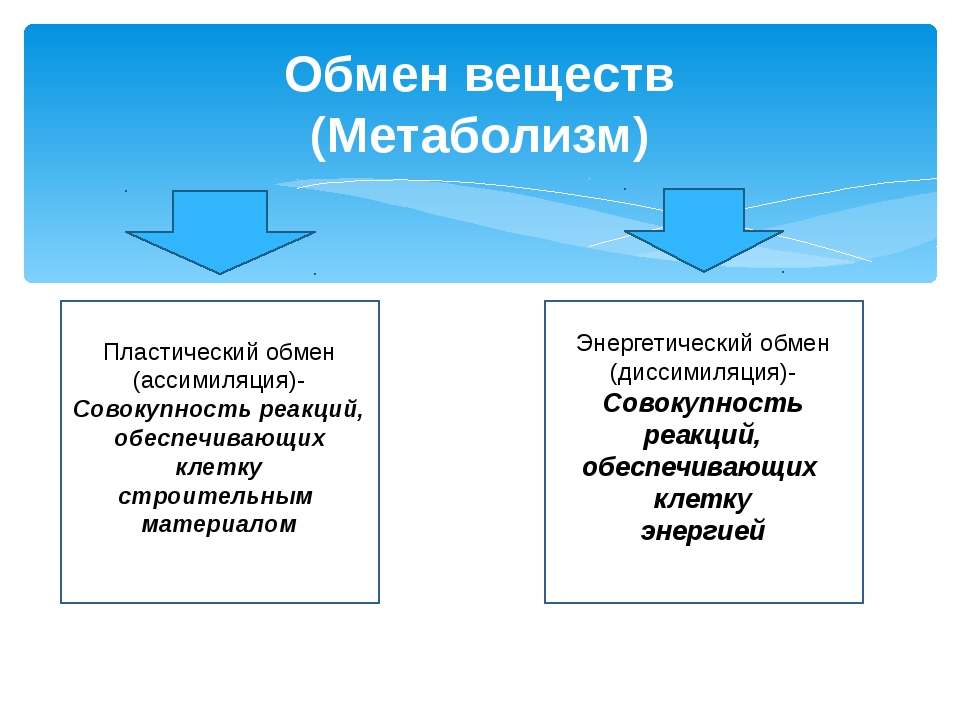 (Палмер и др., 2017). Такие достижения в метаболомике, визуализации метаболитов и моделировании метаболических сетей открывают новые возможности для изучения биохимических сил, определяющих рост и развитие. У этой вновь возникающей области блестящее будущее, и спустя почти 90 лет после публикации трактата Нидхема мы снова исследуем метаболические основы биологии развития.
(Палмер и др., 2017). Такие достижения в метаболомике, визуализации метаболитов и моделировании метаболических сетей открывают новые возможности для изучения биохимических сил, определяющих рост и развитие. У этой вновь возникающей области блестящее будущее, и спустя почти 90 лет после публикации трактата Нидхема мы снова исследуем метаболические основы биологии развития.
Обзор — от метаболизма болезней к точной медицине
В клеточной и молекулярной биологии метаболизм — единственная система, которую можно полностью смоделировать в масштабе генома.Биология метаболических систем предлагает мощные инструменты абстракции для моделирования всех известных метаболических реакций в клетке, что позволяет получить снимок, близкий к наблюдаемому фенотипу. В этом обзоре мы рассматриваем 15 лет моделирования метаболизма человека. Мы показываем, что, хотя за последние пять лет не произошло значительных улучшений в размерах наборов генов и метаболитов в моделях метаболизма человека, их точность быстро растет.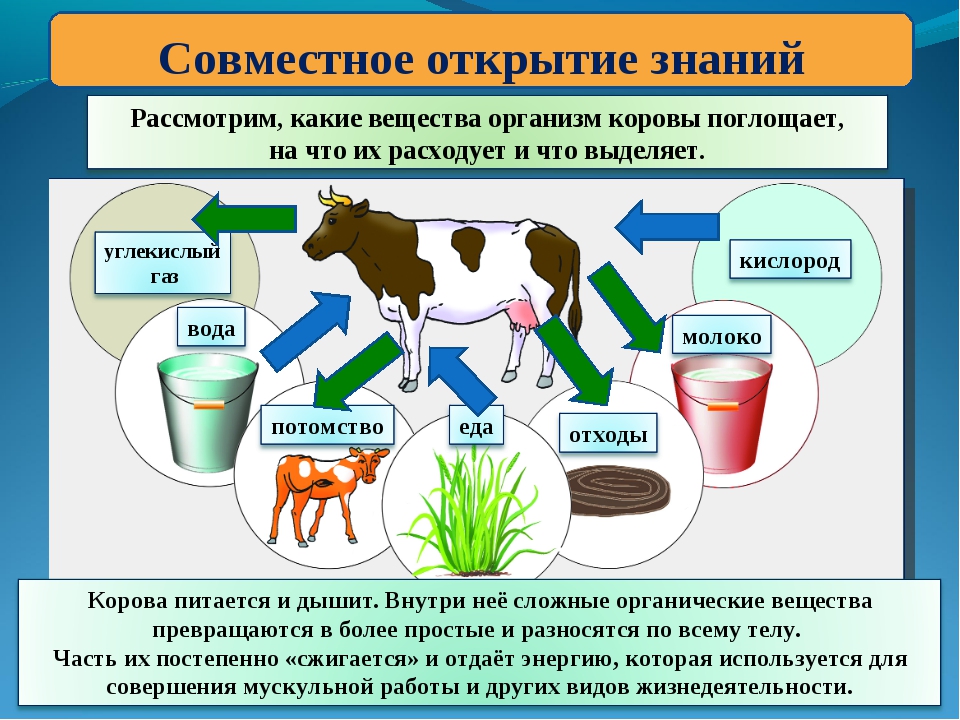 Мы также описываем, как метаболические модели, специфичные для состояния, ткани и пациента, проливают свет на клеточно-специфические изменения, происходящие в метаболической сети, что позволяет прогнозировать биомаркеры метаболизма заболевания.Наконец, мы обсудим текущие проблемы и будущие перспективные направления в этой области исследований, включая машинное / глубокое обучение и точную медицину. В эпоху омики профилирование пациентов и биологических процессов с многомерной точки зрения становится все более распространенным и менее дорогостоящим. Начиная с многомерных данных, собранных у пациентов, и испытаний N-of-1, в которых отдельные пациенты представляют собой разные тематические исследования, методы построения моделей и интеграции данных используются для создания моделей для конкретных пациентов.В сочетании с современными методами машинного обучения это позволит охарактеризовать фенотип заболевания каждого пациента и предложить решения для точной медицины, что приведет к профилактической медицине, сокращению лечения и клиническим испытаниям in silico, .
Мы также описываем, как метаболические модели, специфичные для состояния, ткани и пациента, проливают свет на клеточно-специфические изменения, происходящие в метаболической сети, что позволяет прогнозировать биомаркеры метаболизма заболевания.Наконец, мы обсудим текущие проблемы и будущие перспективные направления в этой области исследований, включая машинное / глубокое обучение и точную медицину. В эпоху омики профилирование пациентов и биологических процессов с многомерной точки зрения становится все более распространенным и менее дорогостоящим. Начиная с многомерных данных, собранных у пациентов, и испытаний N-of-1, в которых отдельные пациенты представляют собой разные тематические исследования, методы построения моделей и интеграции данных используются для создания моделей для конкретных пациентов.В сочетании с современными методами машинного обучения это позволит охарактеризовать фенотип заболевания каждого пациента и предложить решения для точной медицины, что приведет к профилактической медицине, сокращению лечения и клиническим испытаниям in silico, .
1. Введение
С появлением биоинформатики и вычислительной биологии вычислительные и математические методы могут обеспечить точное моделирование биологических процессов. Наиболее широко используемые подходы к анализу данных omics в основном сосредоточены на геномике, транскриптомике и протеомике с помощью дифференциальной экспрессии или анализа коэкспрессии на основе сети.Однако сами по себе гены и их экспрессия не всегда являются надежным индикатором клеточного фенотипа. При характеристике фенотипического результата, полагаясь исключительно на профили экспрессии гена или белка, упускается из виду сильно нелинейное взаимодействие между этими биологическими слоями. Такие подходы часто упускают из виду уровень метаболизма, густую сеть биохимических реакций, происходящих в клетке с целью преобразования питательных веществ в энергию и строительные блоки клетки.
Будучи наиболее хорошо охарактеризованной сетью в биологических системах, а также наиболее близкой к фенотипу, метаболизм, возможно, является лучшим индикатором физиологического состояния клетки [1].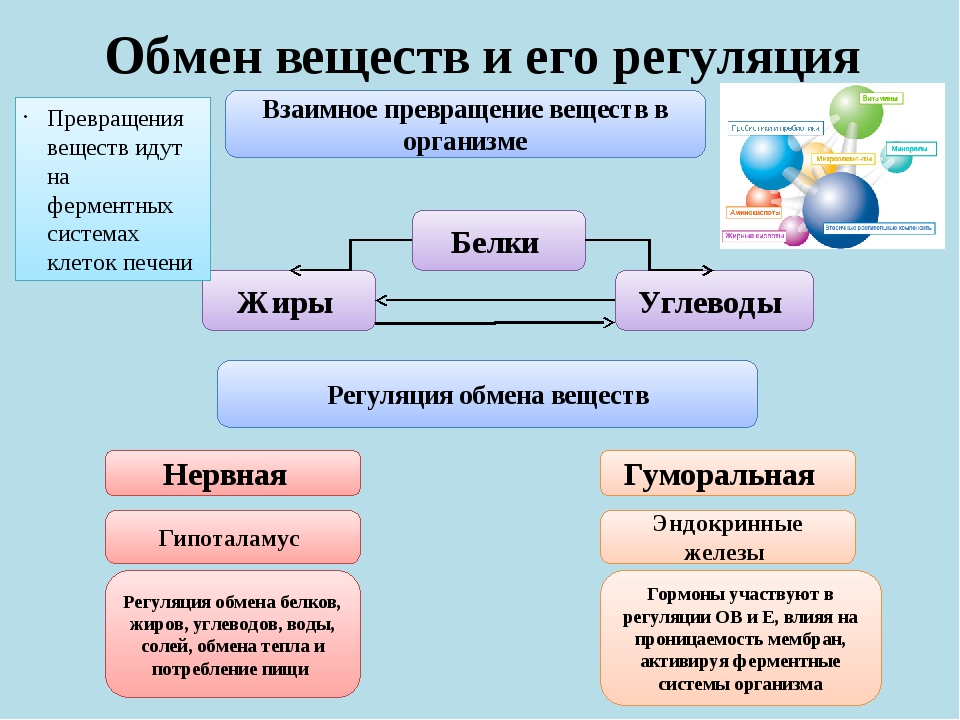 Когда-то считавшийся лишь пассивным результатом состояния клетки, теперь он широко признан как основной фактор, влияющий на клеточное поведение. В частности, он играет ключевую роль в ряде заболеваний, включая диабет, нейродегенеративные заболевания и рак, отличительным признаком которых в настоящее время считается изменение метаболизма [2].
Когда-то считавшийся лишь пассивным результатом состояния клетки, теперь он широко признан как основной фактор, влияющий на клеточное поведение. В частности, он играет ключевую роль в ряде заболеваний, включая диабет, нейродегенеративные заболевания и рак, отличительным признаком которых в настоящее время считается изменение метаболизма [2].
Недавняя доступность высокопроизводительных данных о нескольких уровнях биологической организации (омики) позволяет картировать клеточные процессы на уровнях генов, мРНК, белков и метаболитов (рис. 1).В одном эксперименте эти измерения часто выполняются как на уровне «генотипа» (т.е. относящиеся к генетическим элементам генома), так и на уровне «фенотипа» (форма и функция клетки). Фундаментальным вопросом в системной биологии является определение и понимание взаимосвязи генотип-фенотип [3]. Механистическая связь между генотипом и фенотипом обеспечивается моделями метаболизма в масштабе генома, которые содержат все известные биохимические реакции, происходящие в клетке. Такие модели были созданы с учетом десятилетий исследований в области биохимии и в большинстве случаев способны предсказать клеточный фенотип с высокой точностью.
Такие модели были созданы с учетом десятилетий исследований в области биохимии и в большинстве случаев способны предсказать клеточный фенотип с высокой точностью.
Моделирование на основе ограничений — наиболее широко используемый подход для моделирования поведения метаболизма, часто предполагающий, что клетки должны выполнять заданную задачу (например, производство, рост или пролиферацию АТФ) или оптимизировать производство заданного сложный. Такие модели имеют два основных преимущества: во-первых, они не нуждаются в динамических или кинетических данных, поскольку они основаны на балансе масс в метаболической сети; во-вторых, они подходят для интеграции различных атомных слоев в масштабе генома, чтобы улучшить их прогностические характеристики.В частности, были предложены методы многомерной «вертикальной» интеграции, включающие в себя томные слои (в основном транскриптомику и протеомику) [4–6]. И наоборот, методы «горизонтальной» интеграции сосредоточены на моделировании различных сред, раковых заболеваний или условий роста, исходя из одной и той же модели [7–11]. Такая мультиомная интеграция в моделях в масштабе генома обеспечивает механистическую связь между генотипом и их фенотипическими наблюдаемыми [12-15]. Это ключевая дополнительная особенность, которой обладают такие модели по сравнению с исследованиями общегеномных ассоциаций (GWAS), которые способны связывать вариации генов с фенотипическими признаками, но не обеспечивают механистического объяснения наблюдаемых ассоциаций [16].
Такая мультиомная интеграция в моделях в масштабе генома обеспечивает механистическую связь между генотипом и их фенотипическими наблюдаемыми [12-15]. Это ключевая дополнительная особенность, которой обладают такие модели по сравнению с исследованиями общегеномных ассоциаций (GWAS), которые способны связывать вариации генов с фенотипическими признаками, но не обеспечивают механистического объяснения наблюдаемых ассоциаций [16].
В этом обзоре мы сосредоточены на биологии метаболических систем и, в частности, на моделировании метаболизма человека. Изначально испытанная и подтвержденная с помощью микроорганизмов, биология метаболических систем в настоящее время получает широкое распространение для тканей человека и биомедицинских приложений [17]. Мы предоставляем всесторонний обзор метаболических моделей, разработанных на данный момент для клеток человека, обсуждая их сильные стороны и ограничения. Мы показываем, что основное внимание в сообществе метаболического моделирования сместилось с увеличения размера моделей — «подхода, ориентированного на размер модели», часто за счет надежности прогнозов, — на командные подходы к курированию, когда алгоритмы заполнения пробелов и обширное ручное управление повышает надежность прогнозов без необходимости увеличения (а на самом деле часто уменьшения) количества компонентов в модели. Мы также описываем ключевые истории успеха, текущие проблемы и будущие направления в области моделирования метаболизма человека, включая интеграцию с методами машинного обучения и точной медициной.
Мы также описываем ключевые истории успеха, текущие проблемы и будущие направления в области моделирования метаболизма человека, включая интеграцию с методами машинного обучения и точной медициной.
2. Биология метаболических систем
Понимание роли отдельных компонентов в биологической системе — важный шаг для выяснения или прогнозирования ее поведения. Однако основная тема системной биологии — исследование этих систем с применением комплексного подхода. Фактически, изучение взаимодействий между различными компонентами также дает представление о функционировании и поведении компонентов, взятых независимо [18].
Метаболизм — это самая большая полностью описанная биологическая сеть. Это суррогат фенотипа, способный выявить фенотипические изменения, вызванные единичными или комбинированными омиками или физиологическими факторами. Он также имеет то преимущество, что является единственной системой, которую можно полностью смоделировать в масштабе генома. В результате он играет центральную роль в выяснении связи генотип-фенотип. Системные подходы успешно применялись в течение последнего десятилетия для исследования метаболических сетей, состоящих из набора химических реакций и пула метаболитов.Первые примеры биологии метаболических систем появились в 1999 г. и были сосредоточены на моделировании, соединении и моделировании нескольких клеточных процессов [19]. Полноклеточное моделирование, названное «большой проблемой 21 века», все еще является активной областью исследований [20–24]. В биологии метаболических систем значение моделирования клеточного метаболизма не только в объяснении биологического процесса, но и в прогнозировании. Модель может использоваться для выдвижения гипотез, которые можно проверить, или для выявления неожиданного поведения, которое можно дополнительно исследовать in vitro .
Системные подходы успешно применялись в течение последнего десятилетия для исследования метаболических сетей, состоящих из набора химических реакций и пула метаболитов.Первые примеры биологии метаболических систем появились в 1999 г. и были сосредоточены на моделировании, соединении и моделировании нескольких клеточных процессов [19]. Полноклеточное моделирование, названное «большой проблемой 21 века», все еще является активной областью исследований [20–24]. В биологии метаболических систем значение моделирования клеточного метаболизма не только в объяснении биологического процесса, но и в прогнозировании. Модель может использоваться для выдвижения гипотез, которые можно проверить, или для выявления неожиданного поведения, которое можно дополнительно исследовать in vitro .
Существуют различные методы моделирования метаболической системы: стационарный анализ (например, FBA) включает набор линейных уравнений, а кинетическое моделирование включает обыкновенные дифференциальные уравнения (ODE). Каждая переменная представляет собой изменение концентрации метаболита в динамическом или устойчивом состоянии, где концентрация зависит от скорости реакций, которые производят и потребляют этот метаболит [25]. Кинетические модели не предполагают установившегося состояния и поэтому могут моделировать высокодинамичные механизмы, включая аллостерическую и посттрансляционную регуляцию, концентрации метаболитов и термодинамику.Такие системы на основе ОДУ содержат большое количество уравнений (дифференциальных или алгебраических) и требуют уникальных значений кинетических параметров. Они очень эффективны при прогнозировании поведения небольших систем, где можно собрать достаточно экспериментальных данных для калибровки модели и оценки параметров [26]. Более того, в отличие от стандартных методов, основанных на FBA, кинетика реакции может быть учтена, а концентрации метаболитов могут быть смоделированы явно, и, следовательно, данные внутриклеточной метаболомики могут быть интегрированы напрямую [27].
Каждая переменная представляет собой изменение концентрации метаболита в динамическом или устойчивом состоянии, где концентрация зависит от скорости реакций, которые производят и потребляют этот метаболит [25]. Кинетические модели не предполагают установившегося состояния и поэтому могут моделировать высокодинамичные механизмы, включая аллостерическую и посттрансляционную регуляцию, концентрации метаболитов и термодинамику.Такие системы на основе ОДУ содержат большое количество уравнений (дифференциальных или алгебраических) и требуют уникальных значений кинетических параметров. Они очень эффективны при прогнозировании поведения небольших систем, где можно собрать достаточно экспериментальных данных для калибровки модели и оценки параметров [26]. Более того, в отличие от стандартных методов, основанных на FBA, кинетика реакции может быть учтена, а концентрации метаболитов могут быть смоделированы явно, и, следовательно, данные внутриклеточной метаболомики могут быть интегрированы напрямую [27].
Однако для больших систем использование кинетического моделирования остается сложной задачей. Растущий спрос на анализ в масштабе генома на системном уровне в последнее время привел к широкому использованию моделей устойчивого состояния на основе ограничений и их расширений для нестационарного состояния. Этому также способствовало увеличение доступности мультиомных данных и фенотипической информации, используемых для динамического ограничения модели и, следовательно, часто компенсирующей отсутствие регуляторного и кинетического моделирования.
После ряда успешных попыток построения многомасштабных кинетических моделей и моделей, основанных на ограничениях [28, 29], достижение правильной комбинации (и, следовательно, компромисса) кинетического моделирования и допущений о стационарном состоянии, вероятно, станет следующим шагом для метаболического модельное сообщество. Такие методы, как нестационарный FBA (uFBA) [4], были недавно разработаны для ослабления допущения о стационарном состоянии в моделях в масштабе генома с целью моделирования динамических клеточных состояний, возникающих в результате изменений концентрации внутренних метаболитов. Также был предложен подход к интеграции кинетики реакции с устойчивыми метаболическими сетями, при котором информация об общих метаболитах различных реакций в масштабе генома используется для информирования о взаимодействиях между реакциями и прогнозирования концентраций метаболитов в подходе сетевой кинетики [30]. . В более общем плане, из-за большего диапазона прогнозов, которые кинетические модели могут выполнять по сравнению с подходами стационарного состояния, их расширение в сторону масштаба генома или использование информации, полученной из моделирования модели в масштабе генома, является многообещающим направлением, которое повысит их пространственное и временное разрешение. .
Также был предложен подход к интеграции кинетики реакции с устойчивыми метаболическими сетями, при котором информация об общих метаболитах различных реакций в масштабе генома используется для информирования о взаимодействиях между реакциями и прогнозирования концентраций метаболитов в подходе сетевой кинетики [30]. . В более общем плане, из-за большего диапазона прогнозов, которые кинетические модели могут выполнять по сравнению с подходами стационарного состояния, их расширение в сторону масштаба генома или использование информации, полученной из моделирования модели в масштабе генома, является многообещающим направлением, которое повысит их пространственное и временное разрешение. .
2.1. Модели метаболизма в масштабе генома
Модели метаболизма в масштабе генома содержат все известные метаболические реакции в организме и поэтому могут служить функциональными базами данных клеточно-специфического метаболизма. Модель в масштабе генома строится с использованием следующего процесса. Сначала создается предварительная реконструкция, начиная с генома и включая все кодируемые геномом метаболические реакции. Черновой вариант реконструкции также включает аннотированные данные о ферментах, реакциях и путях из таких баз данных, как KEGG [31], BioCyc [32] и BRENDA [33].Также включены подробные сведения о том, какие гены контролируют каждую реакцию. Затем последовательность шагов ручного курирования улучшает черновую реконструкцию, собирая доказательства, чтобы доказать или опровергнуть наличие реакции в сети организма.
Сначала создается предварительная реконструкция, начиная с генома и включая все кодируемые геномом метаболические реакции. Черновой вариант реконструкции также включает аннотированные данные о ферментах, реакциях и путях из таких баз данных, как KEGG [31], BioCyc [32] и BRENDA [33].Также включены подробные сведения о том, какие гены контролируют каждую реакцию. Затем последовательность шагов ручного курирования улучшает черновую реконструкцию, собирая доказательства, чтобы доказать или опровергнуть наличие реакции в сети организма.
Построение модели в масштабе генома представляет собой отправную точку для анализа баланса потоков (FBA, см. Следующие подразделы). Наконец, модель запускается и проверяется путем сравнения ее прогнозов с существующими экспериментальными результатами, и проводятся новые эксперименты in silico для дальнейшего улучшения и проверки модели.Для получения более подробной информации о том, как построить метаболическую модель из последовательности ДНК организма, читатель может найти полные протоколы [34–36].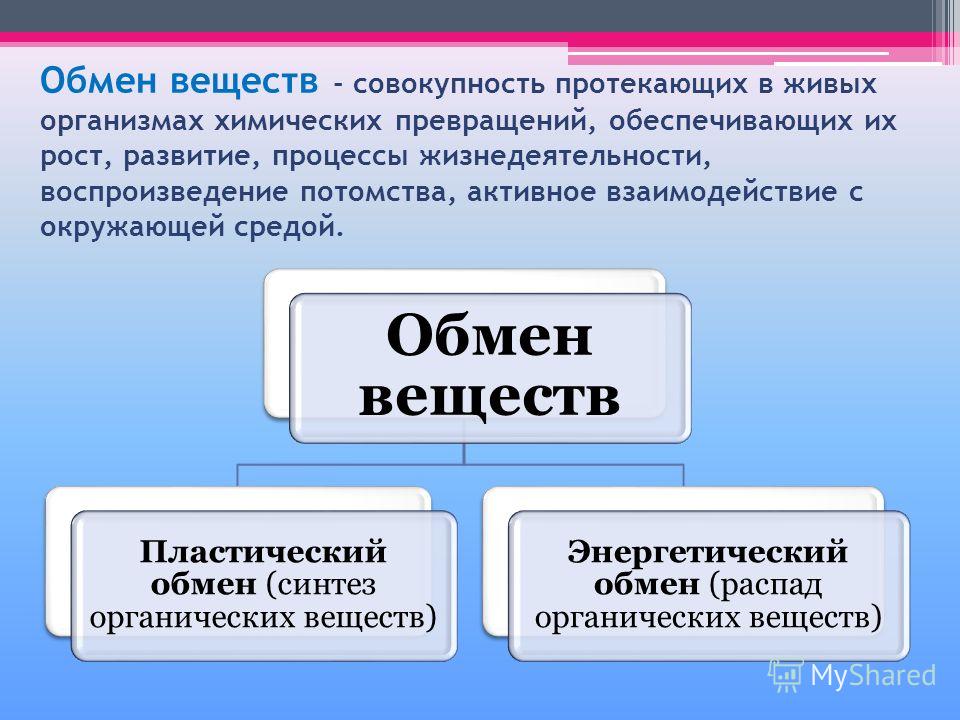
Большинство моделей в масштабе генома аннотированы тщательно подобранными ассоциациями ген-белок-реакция (правила GPR), связывающими гены с ферментами (рис. 2). Такие аннотации открывают путь для наложения многомерных данных на модели, используя их в качестве omic-каркасов (см. Раздел 2.3). Поскольку омические данные могут быть количественно определены в зависимости от состояния, ткани и пациента (например,g., транскриптомные профили, уровни белка и концентрации метаболитов), модели с правилами GPR могут служить базой для создания персонализированных метаболических моделей. Например, персонализированные прогнозы с использованием таких моделей могут привести к точной фенотипической характеристике пациентов (см. Раздел 4).
2.2. Моделирование на основе ограничений и анализ баланса потоков
Анализ баланса потоков (FBA) — это наиболее широко используемый метод на основе ограничений для прогнозирования распределений потоков и сетевых возможностей в моделях в масштабе генома [37]. FBA доказала свою полезность благодаря своей способности обрабатывать большие сети и прогнозировать распределение потоков в масштабе генома. Он требует информации о биохимических реакциях и стехиометрических коэффициентах, но не включает кинетические параметры. Это делает его хорошо подходящим для исследований метаболической инженерии, которые идентифицируют и характеризуют оптимальные нарушения, такие как различные субстраты или генетические вмешательства (например, нокауты), ведущие к обязательной связи между скоростью роста и избыточной продукцией желаемого метаболита [38–42].В целом, FBA — мощный инструмент для прогнозирования поведения клеток в различных метаболических условиях.
FBA доказала свою полезность благодаря своей способности обрабатывать большие сети и прогнозировать распределение потоков в масштабе генома. Он требует информации о биохимических реакциях и стехиометрических коэффициентах, но не включает кинетические параметры. Это делает его хорошо подходящим для исследований метаболической инженерии, которые идентифицируют и характеризуют оптимальные нарушения, такие как различные субстраты или генетические вмешательства (например, нокауты), ведущие к обязательной связи между скоростью роста и избыточной продукцией желаемого метаболита [38–42].В целом, FBA — мощный инструмент для прогнозирования поведения клеток в различных метаболических условиях.
FBA — это метод линейного программирования, который моделирует установившееся состояние в сети химических реакций [43]. FBA-представление модели в масштабе генома построено на основе стехиометрической матрицы, содержащей стехиометрические коэффициенты каждого метаболита (в строках) в каждой реакции сети (в столбцах). Стехиометрический коэффициент метаболита в данной реакции положительный, если метаболит образуется в результате реакции, и отрицательный, если он потребляется.Недоопределенная линейная система уравнений определяется из стехиометрической матрицы, где неизвестные представлены вектором скоростей потока реакции. Дополнительные ограничения включены как нижняя и верхняя границы потоков в (). Они учитывают рост или физиологическое состояние и могут использоваться для включения данных омиков. Одна или несколько клеточных целей (обычно связанных с ростом или энергией, например, биомасса или АТФ) или их линейная комбинация, наконец, выбираются для максимизации или минимизации при вышеупомянутых ограничениях, таким образом решая линейную программу.
Стехиометрический коэффициент метаболита в данной реакции положительный, если метаболит образуется в результате реакции, и отрицательный, если он потребляется.Недоопределенная линейная система уравнений определяется из стехиометрической матрицы, где неизвестные представлены вектором скоростей потока реакции. Дополнительные ограничения включены как нижняя и верхняя границы потоков в (). Они учитывают рост или физиологическое состояние и могут использоваться для включения данных омиков. Одна или несколько клеточных целей (обычно связанных с ростом или энергией, например, биомасса или АТФ) или их линейная комбинация, наконец, выбираются для максимизации или минимизации при вышеупомянутых ограничениях, таким образом решая линейную программу.
FBA, таким образом, основывается на двух основных предположениях: (i) гомеостатическое предположение: организм достиг устойчивого состояния, при котором концентрации метаболитов постоянны, а набор питательных веществ постоянно преобразуется для генерации биомассы. (Ii) (многоуровневая) оптимальность : в каждом состоянии организм стремится максимизировать одну или несколько целей, обычно связанных с ростом, производством биотехнологических соединений (например, ацетатный обмен) и важных энергоносителей (например, АТФ).
(Ii) (многоуровневая) оптимальность : в каждом состоянии организм стремится максимизировать одну или несколько целей, обычно связанных с ростом, производством биотехнологических соединений (например, ацетатный обмен) и важных энергоносителей (например, АТФ).
FBA широко используется в системной биологии для количественной оценки всего устойчивого метаболического состояния клетки и расчета распределения ее потока. Рассмотрены все известные метаболические реакции в данной клетке, и они математически описаны таким образом, чтобы можно было моделировать различные состояния и конфигурации сети химических реакций. Интуитивно понятно, что стационарные ограничения, используемые в FBA, можно рассматривать как законы Кирхгофа, применяемые к любому узлу, представляющему метаболит в сети: поток через каждый метаболит в сети должен быть постоянным, а именно, входной поток должен равняться выходному потоку.Комбинация стационарных ограничений и ограничений пропускной способности на потоки реакции представляет собой систему линейных однородных уравнений и неравенств; таким образом, его пространство решений представляет собой выпуклый многогранный конус, представляющий допустимые распределения потоков.
Предположения FBA и сведение проблемы к линейной программе может вызвать некоторые ограничения. Во-первых, включение или прогнозирование концентраций метаболитов является сложной задачей и требует динамического FBA [44], ослабления устойчивого состояния [4], методов суммирования потоков [45] или термодинамических подходов [46].Во-вторых, надежность распределений потоков во многом зависит от выбранной целевой функции (см. Раздел 2.4), от качества реконструкции и от метода, используемого для получения решения. Регуляризованные методы FBA помогают облегчить эту проблему, как обсуждается далее в этом разделе и в разделе 5. Наконец, FBA не обладает способностью непосредственно моделировать регуляторные эффекты или посттранскрипционную регуляцию уровней экспрессии. Аналогичным образом, изменения, происходящие во время быстрых переходных процессов (например, возмущения в микросреде клетки) не могут быть смоделированы динамически, но могут быть аппроксимированы пошагово до / после моделирования [47]. Хотя правило устойчивого состояния было оспорено, и были предложены вероятностные подходы, чтобы ослабить равенство устойчивого состояния [48], это предположение позволяет использовать линейные системы и линейное программирование, снижая вычислительные требования и позволяя быстро моделировать масштаб генома. модели в различных условиях роста или физиологических условиях.
Хотя правило устойчивого состояния было оспорено, и были предложены вероятностные подходы, чтобы ослабить равенство устойчивого состояния [48], это предположение позволяет использовать линейные системы и линейное программирование, снижая вычислительные требования и позволяя быстро моделировать масштаб генома. модели в различных условиях роста или физиологических условиях.
При решении линейных программ для FBA разные решатели могут давать разные решения из-за различий в численной реализации и существования множества альтернативных оптимальных решений [49–51].Хотя значение целевой функции одинаково во всех оптимальных решениях, другие скорости потока могут варьироваться. Поэтому наличие уникального решения важно, когда полное распределение потока используется для дальнейшего анализа или в качестве функции прогнозных алгоритмов. Чтобы избежать вырожденных решений, предоставляемых стандартными подходами FBA, был предложен экономичный FBA (pFBA) с целью минимизировать общий поток, переносимый метаболической сетью после максимизации основной цели [52].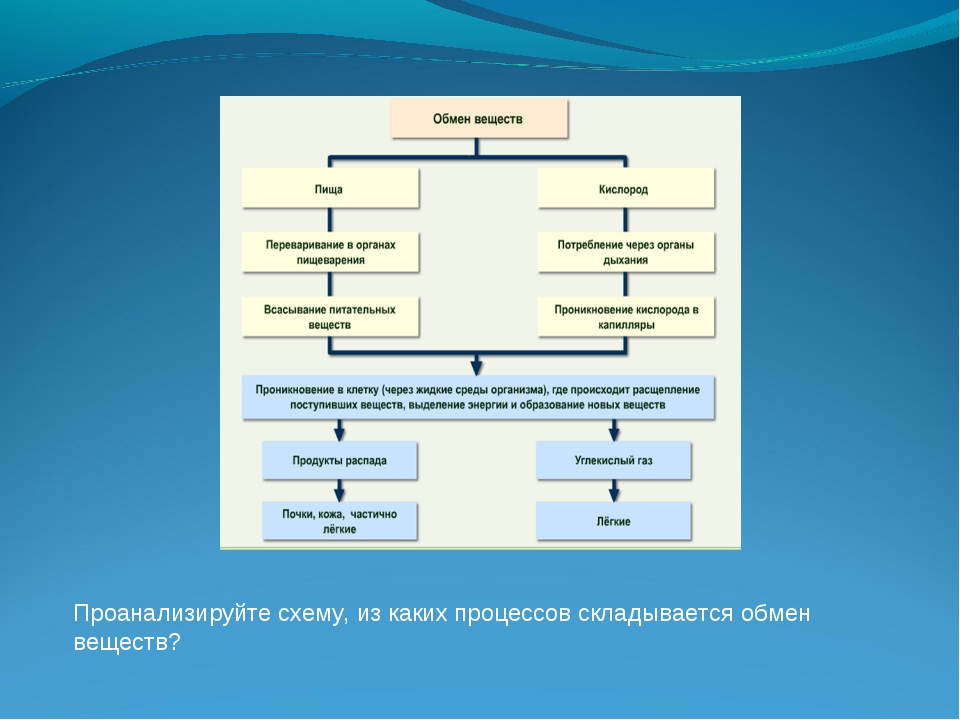 Однако в некоторых случаях pFBA дает менее правдоподобные результаты в метаболизме центрального углерода и в гликолитическом пути по сравнению со стандартными методами FBA [53]. Кроме того, pFBA использует L1-норму (минимизацию суммы абсолютных значений магнитного потока), которая не гарантирует уникального решения, поскольку оно не является строго выпуклым [54]. Для решения таких задач минимизации с гарантией единственности решения вместо этого следует использовать L2-норму (минимизация суммы квадратов значений потока). Дополнительные подходы для облегчения проблемы альтернативных оптимальных решений включают геометрический FBA [55] и выборку [56].
Однако в некоторых случаях pFBA дает менее правдоподобные результаты в метаболизме центрального углерода и в гликолитическом пути по сравнению со стандартными методами FBA [53]. Кроме того, pFBA использует L1-норму (минимизацию суммы абсолютных значений магнитного потока), которая не гарантирует уникального решения, поскольку оно не является строго выпуклым [54]. Для решения таких задач минимизации с гарантией единственности решения вместо этого следует использовать L2-норму (минимизация суммы квадратов значений потока). Дополнительные подходы для облегчения проблемы альтернативных оптимальных решений включают геометрический FBA [55] и выборку [56].
Доступно несколько инструментов для реконструкции метаболической модели, моделирования на основе ограничений, FBA и связанных анализов. К ним относятся COBRA [57], его версия для Python COBRApy [58], RAVEN [59], PathwayTools [60] и FAME [61]. Полный список можно найти в соответствующих обзорах и сравнительных статьях [62, 63].
2.
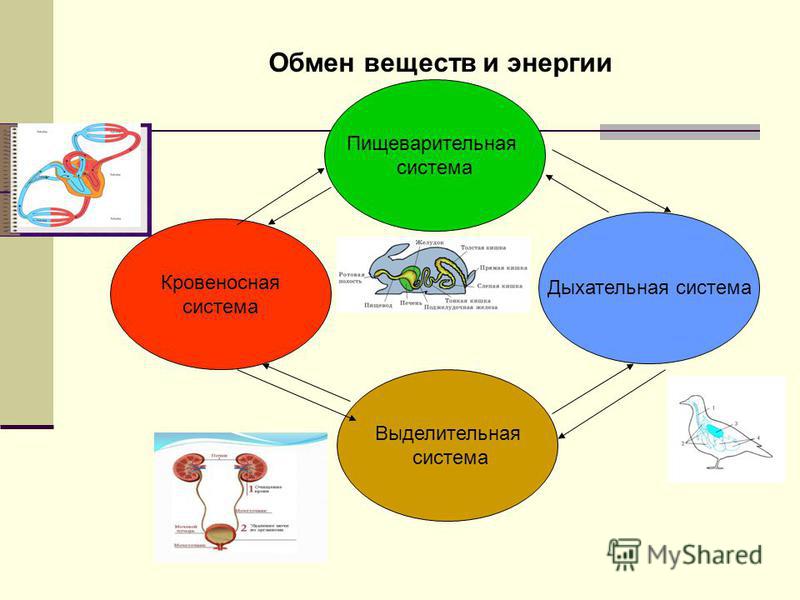 3. Анализ мультиомного баланса потока
3. Анализ мультиомного баланса потокаХотя некоторые биологические результаты могут быть выяснены с помощью одномомного анализа (например, белок-белковое взаимодействие или генные сети), подавляющее большинство фенотипических наблюдаемых явлений является результатом более всесторонних взаимосвязей между множественными омиками [64].Регуляторные механизмы также имеют место на разных атомных уровнях (включая транскрипцию, трансляцию и метаболические реакции). Сложное взаимодействие между этими уровнями отвечает за поведение клеток. Для интеграции одномерных сетей был предложен многомерный анализ, основанный на сетях совместного выражения или взаимодействия [65]. Однако в таких методах часто не учитываются механистические связи между атомными слоями, полученные из предшествующих молекулярных знаний.
Идея мультиомного FBA состоит в том, что, отображая атомные данные на in silico моделях метаболизма, можно получить обогащенное метаболизмом представление любого заданного омического профиля.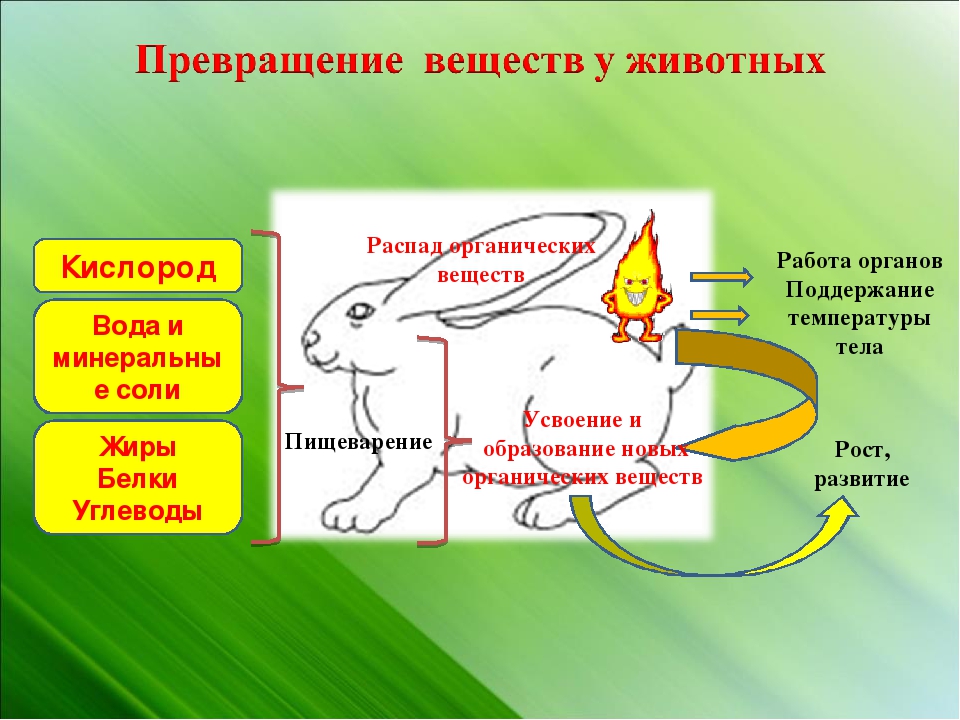 В этом контексте, а также позволяя моделировать большие, обычно в масштабе генома, системы за несколько секунд процессорного времени, FBA также имеет то преимущество, что облегчает введение дополнительных ядерных слоев экспериментальных данных, которые могут быть наложены на модель. , используя правила GPR или аннотации метаболитов, чтобы наложить ограничения на границы потока. Омические метаболические модели рассматривают механистические отношения между омиками, поэтому правильно представляют априорную информацию, имеющуюся в биохимических сетях [66].Мультиомный FBA позволяет определить важность гена или фермента через его прогнозируемую функцию, что позволяет избежать приближений или статистических методов, основанных только на значении уровня его экспрессии.
В этом контексте, а также позволяя моделировать большие, обычно в масштабе генома, системы за несколько секунд процессорного времени, FBA также имеет то преимущество, что облегчает введение дополнительных ядерных слоев экспериментальных данных, которые могут быть наложены на модель. , используя правила GPR или аннотации метаболитов, чтобы наложить ограничения на границы потока. Омические метаболические модели рассматривают механистические отношения между омиками, поэтому правильно представляют априорную информацию, имеющуюся в биохимических сетях [66].Мультиомный FBA позволяет определить важность гена или фермента через его прогнозируемую функцию, что позволяет избежать приближений или статистических методов, основанных только на значении уровня его экспрессии.
Следуя этому подходу, FBA и его многомерные модификации были использованы для прогнозирования метаболического ответа на данное состояние в свете множества клеточных целей, которым клетка должна соответствовать [67, 68]. Недавние исследования также были нацелены на улучшение предсказательной способности метаболической модели и выяснение взаимосвязи генотип-фенотип с помощью компьютерного анализа на нескольких атомных уровнях. Прогнозы распределения потоков после интеграции многомерных данных в сочетании с экспериментальными методами успешно использовались для формулирования новых биологических гипотез. Эти методы сводят проблему определения распределения потока через все реакции в системе при заданном росте или физиологических условиях до легко управляемой линейной программы в предположении устойчивого состояния и оптимальности. Благодаря своей масштабируемости и точности эти методы широко используются, например, для прогнозирования фенотипов роста бактерий в конкретных условиях окружающей среды [69, 70], для определения новых терапевтических мишеней против инфекций [71, 72], для характеристики метаболизма рака различных клеточных линий [73–76], а также для создания тканеспецифичных моделей рака и нормальных тканей для 17 тканей [77].Дополнительные примеры того, как этот подход использовался для характеристики метаболизма болезни, приведены в разделе 4. Для всестороннего обзора методов многомерной интеграции в моделях FBA читатель может отослать читателя к недавним обзорам метаболического моделирования, основанного на омике [78–81] .
Прогнозы распределения потоков после интеграции многомерных данных в сочетании с экспериментальными методами успешно использовались для формулирования новых биологических гипотез. Эти методы сводят проблему определения распределения потока через все реакции в системе при заданном росте или физиологических условиях до легко управляемой линейной программы в предположении устойчивого состояния и оптимальности. Благодаря своей масштабируемости и точности эти методы широко используются, например, для прогнозирования фенотипов роста бактерий в конкретных условиях окружающей среды [69, 70], для определения новых терапевтических мишеней против инфекций [71, 72], для характеристики метаболизма рака различных клеточных линий [73–76], а также для создания тканеспецифичных моделей рака и нормальных тканей для 17 тканей [77].Дополнительные примеры того, как этот подход использовался для характеристики метаболизма болезни, приведены в разделе 4. Для всестороннего обзора методов многомерной интеграции в моделях FBA читатель может отослать читателя к недавним обзорам метаболического моделирования, основанного на омике [78–81] .
Текущие усилия по моделированию направлены на включение дополнительных биологических знаний в модели. Чтобы учесть промискуитет ферментов при сверхэкспрессии или недостаточной экспрессии гена, правила GPR были предложены в качестве дополнительных строк стехиометрической матрицы, как подматрица ферментов по реакциям [53].Например, если несколько реакций катализируются одним и тем же неразборчивым ферментом, изменение потока в результате недостаточной / сверхэкспрессии фермента распределяется между реакциями (также с учетом других ферментов, участвующих в реакциях), а не запускает a priori эквивалентное увеличение / уменьшение потока во всех реакциях. Использование кодонов также может быть интегрировано через правила GPR [71]. Аннотации сплайс-изоформ, изначально утерянные, а затем просто игнорируемые при реконструкциях человека, также недавно привлекли внимание [82, 83].
Мягкие ограничения могут использоваться для интеграции омических данных в зависимости от реакции, а не с помощью одного математического правила, изменяющего ограничения для всех реакций [84].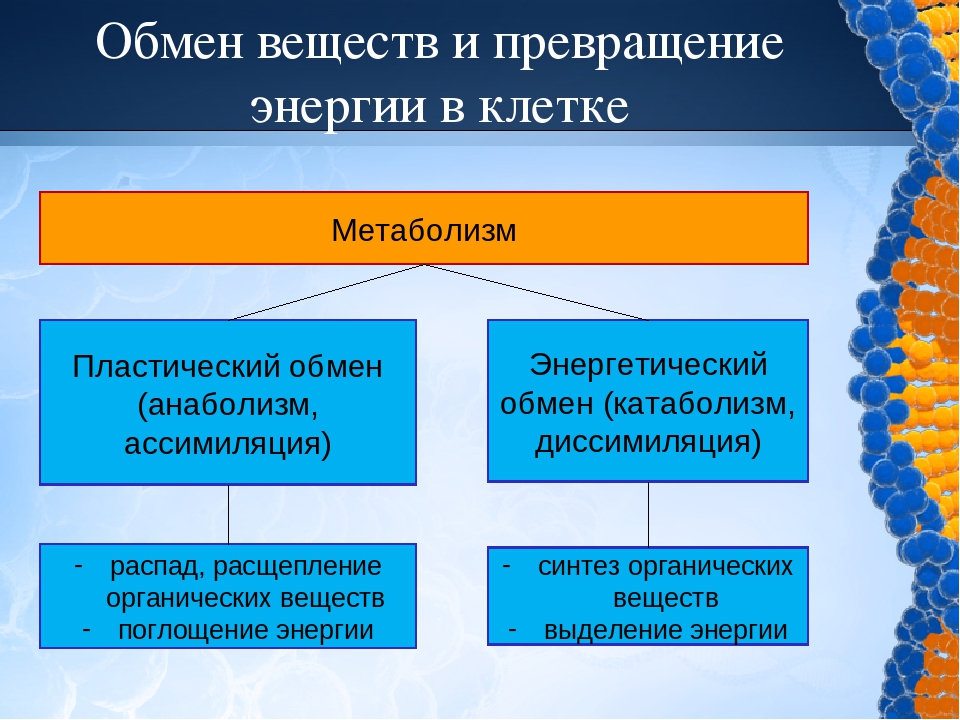 Дополнительные ограничения были также добавлены для интеграции моделей регуляции транскрипции и метаболических моделей. Идея состоит в том, что ограничение моделирует корреляцию между уровнем экспрессии целевого гена и уровнем его регулирующих генов [6]. Для дальнейшего анализа распределения потоков реакции, предсказываемых многомерным FBA, можно провести анализ чувствительности генов, реакций или путей.Определения показателей чувствительности основаны на эффектах нокаута генов [85], реальных возмущениях генов [69] или роли каждого метаболита в снижении термодинамической неопределенности модели [86].
Дополнительные ограничения были также добавлены для интеграции моделей регуляции транскрипции и метаболических моделей. Идея состоит в том, что ограничение моделирует корреляцию между уровнем экспрессии целевого гена и уровнем его регулирующих генов [6]. Для дальнейшего анализа распределения потоков реакции, предсказываемых многомерным FBA, можно провести анализ чувствительности генов, реакций или путей.Определения показателей чувствительности основаны на эффектах нокаута генов [85], реальных возмущениях генов [69] или роли каждого метаболита в снижении термодинамической неопределенности модели [86].
2.4. Выбор целевой функции и учет нескольких клеточных целей
Выбор подходящей целевой функции все еще является проблемой при метаболическом моделировании. Распространенное предположение в системной биологии состоит в том, что клетки стремятся оптимизировать свою метаболическую сеть, чтобы максимизировать скорость роста (биомассу).Однако это до сих пор остается предметом дискуссий [87], как с точки зрения ее состава [88], так и потому, что во многих случаях лучшая цель для клетки не связана с ростом [89]. В то время как модели микроорганизмов могут предполагать, что клетка нацелена на максимальную скорость роста, человеческие клетки могут не обязательно стремиться к максимальной биомассе (хотя это широко принято в качестве цели для раковых клеток). Более того, теперь очевидно, что клеточная цель может меняться между разными клетками в ткани, между тканями, а также со временем для одной и той же клетки.Для тканеспецифических исследований, начиная с общей реконструкции метаболизма человека (например, Recon) и используя универсальный состав биомассы типовых моделей человека, может оказаться ненадежным. Поэтому недавние исследования продвигаются к оценке состава биомассы для конкретных клеток [13, 90]. Также были предложены алгоритмы для генерации и стандартизации реакции биомассы [91, 92], а также для автоматического создания целевой функции [93].
В то время как модели микроорганизмов могут предполагать, что клетка нацелена на максимальную скорость роста, человеческие клетки могут не обязательно стремиться к максимальной биомассе (хотя это широко принято в качестве цели для раковых клеток). Более того, теперь очевидно, что клеточная цель может меняться между разными клетками в ткани, между тканями, а также со временем для одной и той же клетки.Для тканеспецифических исследований, начиная с общей реконструкции метаболизма человека (например, Recon) и используя универсальный состав биомассы типовых моделей человека, может оказаться ненадежным. Поэтому недавние исследования продвигаются к оценке состава биомассы для конкретных клеток [13, 90]. Также были предложены алгоритмы для генерации и стандартизации реакции биомассы [91, 92], а также для автоматического создания целевой функции [93].
Особенно в контексте метаболизма человека, вопрос «Что делает конкретная клетка?» часто имеет более одного правильного ответа. Появляется все больше свидетельств того, что клетки должны решать несколько, обычно конкурирующих, целей для одновременной оптимизации [94]. Одна целевая функция FBA не может охватить их все. Также вероятно, что метаболизм не полностью оптимизирован для какой-либо конкретной цели [95], и эволюция сформировала клетки, чтобы достичь оптимального компромисса между всеми целями [96].
Появляется все больше свидетельств того, что клетки должны решать несколько, обычно конкурирующих, целей для одновременной оптимизации [94]. Одна целевая функция FBA не может охватить их все. Также вероятно, что метаболизм не полностью оптимизирован для какой-либо конкретной цели [95], и эволюция сформировала клетки, чтобы достичь оптимального компромисса между всеми целями [96].
Поэтому были предложены процессы оптимизации, учитывающие несколько целей (например,g., производство белка или энергии, детоксикация, пролиферация) с преимуществом обеспечения метаболической гибкости для возможных реорганизаций, выполняемых во время адаптации к изменениям в условиях окружающей среды [97]. Эти методы включают вероятностный подход [98], лексикографическое упорядочение [99] и сведение к единой цели посредством определения весов для целей [100, 101]. Подходы, основанные на этих идеях, бывают быстрыми и могут создать фронт Парето; однако они упускают субоптимальные решения, решения в невыпуклых областях [102, 103] или отдают предпочтение одной цели [104], поэтому обращаются к многоуровневой проблеме, а не к строго многокритериальной проблеме. Хотя и медленнее, методы, основанные на эволюционных алгоритмах, способны исследовать невыпуклые компромиссы, не требуя объединения целей в единую целевую функцию [105–107]. Изучение набора компромиссных решений между конкурирующими целями, а не единственного решения по максимизации биомассы, также учитывает неоптимальные решения [108].
Хотя и медленнее, методы, основанные на эволюционных алгоритмах, способны исследовать невыпуклые компромиссы, не требуя объединения целей в единую целевую функцию [105–107]. Изучение набора компромиссных решений между конкурирующими целями, а не единственного решения по максимизации биомассы, также учитывает неоптимальные решения [108].
3. 15 лет моделирования метаболизма человека
Изучение метаболизма человека становится все более важным для биомедицинских приложений как подход к пониманию многих заболеваний и аспектов здоровья [109, 110].Понимание метаболического поведения на системном уровне становится возможным благодаря доступности высококачественных реконструкций в масштабе генома, объединяющих обширную метаболическую информацию из различных источников. Первые попытки метаболической реконструкции в масштабе генома были сосредоточены на бактериях ( Haemophilus influenzae [111], затем Escherichia coli [112]) из-за их простоты, доступных последовательностей генома и хорошо известных механизмов утилизации субстрата.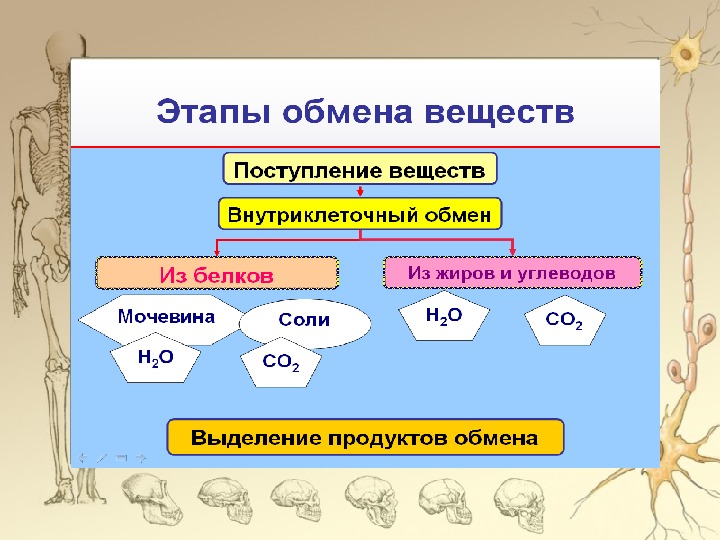 . Реконструкция метаболизма человека, которая потребовала гораздо большего количества путей и большего количества основных питательных веществ, была предпринята на более позднем этапе, после публикации проекта и полных последовательностей генома человека в 2001 и 2004 годах [113, 114].
. Реконструкция метаболизма человека, которая потребовала гораздо большего количества путей и большего количества основных питательных веществ, была предпринята на более позднем этапе, после публикации проекта и полных последовательностей генома человека в 2001 и 2004 годах [113, 114].
HumanCyc [115] (в 2004 г.) и Reactome knowledgebase [116, 117] (в 2005 г.) были первыми успешными попытками создать тщательно подобранную коллекцию биохимических реакций в клетках человека. Первые модели метаболизма человека, опубликованные в 2007 г. — Recon 1 [118] и EHMN (реконструкция метаболической сети человека в Эдинбурге и ее последующая разделенная версия в 2010 г.) [119, 120], позволили лучше охватить метаболическую сеть человека, особенно в компартменты кроме цитозоля. Recon 1 содержит примерно в раз больше уникальных метаболитов и в разы больше реакций, чем HumanCyc.
В 2013 году значительное улучшение количества метаболических процессов, охваченных реконструкцией, было достигнуто с помощью Recon 2 [121]. Это согласованная модель, которая включает все реакции из EHMN, Recon 1, HepatoNet1 [122], а также модуль для окисления ацилкарнитина и жирных кислот [123]. По сравнению с Recon 1, Recon 2 представляет собой значительное улучшение с примерно в раз большим количеством уникальных метаболитов и в 2 раза большим количеством реакций. В 2015 году Recon 2 был доработан с обновленными ассоциациями ген-реакция.Одновременно с этим независимо была построена более крупная реконструкция под названием HMR (база данных метаболических реакций человека) [73] на основе HepatoNet1, Recon1, EHMN, Reactome, HumanCyc, KEGG [31] и Атласа метаболизма человека [124]. Затем в 2014 году HMR был расширен, чтобы включить метаболизм липидов, таким образом, генерируя HMR2 [125].
Несмотря на то, что он содержит большой набор реакций и является консенсусной моделью, прогнозы Recon 2 в некоторых случаях были неверными, в основном из-за ряда неправильно сбалансированных реакций.На этом этапе основной фокус сообщества сместился на курирование существующих моделей. В период с 2014 по 2015 год для Recon 2 был опубликован ряд небольших обновлений и исправлений [126–129]. В 2016 году Swainston et al. провел обширную работу по курированию Recon 2 и его обновлений, что привело к Recon 2.2 [130]. Впервые значительное улучшение существующей модели содержало меньше генов, чем исходная. Фактически, из-за удаления дубликатов и псевдогенов, а также благодаря новому уникальному соглашению об идентификаторах HGNC, Recon 2.2 содержит 112 генов, кодирующих ферменты, меньше, чем Recon 2, и впервые с унифицированными идентификаторами (объединение изоформ сплайсинга в один ген). Recon 2.2 — это серьезное усовершенствование связанного с энергией метаболизма, с новым отделением, имитирующим митохондриальное внутримембранное пространство. В результате прогнозы потока АТФ и биомассы были значительно улучшены, и впервые были получены точные прогнозы максимального выхода АТФ при 14 источниках углерода, как в аэробных, так и анаэробных условиях, и при 20 дополнительных источниках углерода аминокислот в аэробных условиях [130 ].
В 2017 году была получена новая метаболическая модель человека, названная iHsa [131], как расширение и ручное лечение HMR2. iHsa является результатом согласования с метаболической моделью iRno крысы, также представленной в той же статье. Обе модели были созданы параллельно; специфичные для крыс реакции, термодинамически недопустимые реакционные петли и другие неправильные реакции, присутствующие в HMR2 и Recon 2, были удалены из iHsa, в то время как новые реакции были добавлены из KEGG и MetaCyc / BioCyc [132]. Впервые специфические для крыс реакции были удалены из метаболической реконструкции человека.Фактически, обе модели были согласованы и вручную отредактированы с упором на видоспецифические метаболические различия. Четкое определение различий между двумя метаболическими сетями в сочетании с наличием уникальных реакций в обеих моделях открывает возможности для межвидового сравнения человека / крысы.
В 2018 году Recon 3D [133] был разработан на основе Recon 2 путем включения HMR2 и ряда дополнительных наборов реакций, включая реакции, моделирующие взаимодействие хозяина и микроба, реакции для моделирования воздействия лекарств на метаболизм человека, реакции на абсорбцию пищевых соединений, реакции липидного обмена и реакции из наборов данных метаболомики.В модель также были включены трехмерные белковые структуры, данные фармакогеномики и атомно-атомные сопоставления. После обширных этапов ручного лечения и уточнения правил георадара модель была протестирована на согласованность при воспроизведении 431 важной функции человеческого тела. На сегодняшний день Recon 3D является крупнейшей из доступных для метаболизма человека реконструкцией метаболизма, содержащей в раз больше уникальных метаболитов и в разы больше реакций, чем Recon 2.
На рис. 3 обобщен прогресс, достигнутый сообществом моделирования метаболизма за последние 15 лет.Количество генов, метаболитов и реакций указано на рисунке. Чтобы лучше количественно оценить связанность модели, мы вычислили количество генов, нокаут которых дает измеримые эффекты в модели, измеренные с помощью предсказанного потока биомассы. Чтобы учесть толерантность линейного решателя, мы определили ген , необходимый , если его нокаут производит биомассу; мы определили ген неотъемлемый , если его нокаут вызывает изменение биомассы. По сравнению с Recon 2, увеличение числа генов, вариация которых оказывает существенное влияние на скорость роста, показывает, что были успешно предприняты усилия по курированию, заполнению пробелов и проверкам согласованности, как подробно описано в Разделе 3.1.
3.1. От ориентированных на размер «моделей» к ручному курированию
Когда в 2007 году были опубликованы первые реконструкции человеческого метаболизма Recon 1 [118] и EHMN [119], во многих исследованиях предпочтение было отдано Recon 1, поскольку оно содержало больше реакций. Фактически, это было только из-за компартментализации Recon 1, когда метаболиты повторяются в разных компартментах и транспортных реакциях между компартментами. Количество уникальных реакций действительно было больше в EHMN (на 1028 больше реакций и на 1202 метаболитов), и была создана последующая секционированная версия EHMN [120].
Эти частые сравнения между различными моделями показывают, что изначально основное внимание было уделено включению как можно большего числа реакций — подход, ориентированный на размер, — часто за счет курирования и точности. И наоборот, подход, включающий все доступные биологические компоненты в модель, теперь всегда сочетается с обширным групповым курированием, проверками согласованности, единообразием аннотаций и заполнением пробелов. Нередко достигается лучшая модель, удаляя реакции из существующей.Это побудило сообщество сосредоточиться на точности используемых моделей и решателей, а не только на размере модели.
Ручное курирование поэтому считается одним из основных шагов в создании метаболической модели. Такие инструменты, как MetaNetX [134], упрощают курирование и предлагают сопоставления идентификаторов для унификации идентификаторов реакций и метаболитов. Такие усилия по курированию, вероятно, станут еще более важными в ближайшем будущем, учитывая недавние достижения в области автоматизированных инструментов реконструкции, которые теперь могут генерировать полный рабочий проект модели, т.е.g., MicrobesFlux [135], Pathway Tools [136], PathwayBooster [137], CoReCo [138] и Merlin [139]. Учитывая важность получения согласованных и высококачественных моделей, MetaNetX также предоставляет базу данных моделей, созданных после согласования метаболитов и биохимических реакций [134]. Совсем недавно CarveMe [140] и RAVEN 2.0 [59] продемонстрировали высокий потенциал в автоматизации ручных шагов, впервые достигнув точности, напрямую сопоставимой с вручную подобранными моделями при прогнозировании экспериментальных фенотипов.Также доступны инструменты для проверки несоответствий и для визуального изучения модели [141].
За последние несколько лет усилия по курированию привели к сокращению количества несбалансированных реакций, а также количества заблокированных реакций и тупиковых метаболитов. Например, Recon 2.2 не имеет несбалансированных реакций, кроме целевых функций биомассы, а Recon 3 теперь имеет менее 15 заблокированных реакций и тупиков. Более того, поскольку создание контекстно-зависимых моделей с использованием omics зависит от правил георадара, точность этих моделей строго зависит от точности георадара в базовой модели.Как показано на Рисунке 3, покрытие георадара увеличивается. Наконец, соответствие между теоретическими и предсказанными моделями выходами АТФ было улучшено в самых последних моделях. Мы рассчитали среднеквадратичную ошибку (RMSE) между теоретическим и прогнозируемым моделью максимальным выходом АТФ на единицу источника углерода, который был уменьшен с (Recon 2 и более ранние модели) до (в Recon 3) и (в Recon 2.2). . Таблица выходов АТФ и расчеты RMSE представлены в качестве дополнительных материалов.
4.Тканевые и специфические для пациентов данные о метаболизме заболеваний человека
Начиная с общих метаболических реконструкций человека, был опубликован ряд успешных попыток реконструкции тканеспецифичных моделей. К ним относятся мозг [142], адипоциты [143], рак груди [144], сердце [145], почки [146], миоциты [147] и гепатоциты [125, 148]. Дальнейший набор из 32 тканеспецифичных и органоспецифичных моделей был создан путем картирования экспрессии белка на общей человеческой модели HMR2 [125].
Тканевые модели обычно строятся как сокращение от общей модели человека. Было обнаружено, что реакции, которые удаляются при построении тканеспецифической модели, не активны в этой ткани. Реакции удаляются в соответствии с транскриптомными или протеомными данными, собранными в этой ткани, и поэтому тканеспецифичная модель содержит меньше реакций, чем обычная. Неудивительно, что в большинстве случаев они превосходят обычные аналоги, из которых они возникли. На этом этапе необходимо создать новое уравнение биомассы для конкретной ткани или настроить модель, чтобы клетка могла найти альтернативные пути для поддержания своей жизни.Хотя многие методы создания тканеспецифичных моделей ставят цель создать минимальную метаболическую модель, которая удовлетворяет жизнеспособность или набор метаболических задач, использование неминималистических методов для создания тканеспецифичных моделей должно быть предпочтительным. Например, рассмотрение существенности перед удалением реакций из общей модели дает неминимальную тканеспецифичную модель, но улучшает функциональность модели и согласование с экспериментальными данными [149]. Поскольку общие модели представляют собой надмножество моделей, специфичных для ткани, прогнозы общих моделей могут быть верными в задачах, специфичных для ткани, но менее точными.
Контекстно-зависимые модели метаболизма человека пролили свет на начало и прогрессирование болезни в ряде недавних тематических исследований. В этом контексте возобновление интереса к метаболизму болезней связано с тем, что для полной характеристики большинства заболеваний необходимо целостное представление в масштабе генома, а не подход на основе одного гена. Например, при раке различия в метаболических путях между раковыми клетками и их родительской тканью были охарактеризованы с использованием данных омики и метаболической модели человека [75, 150, 151].В результате, тканевые и клеточно-специфические метаболические модели были успешно использованы для идентификации — и последовательной проверки — конкретных мишеней для лекарств, которые ингибируют пролиферацию рака, но не влияют на нормальную пролиферацию клеток [74, 152]. При аналогичном подходе субмодели, построенные на основе моделей в масштабе человеческого генома, были использованы для генерации нескольких проверяемых гипотез, например, для сравнения почечных мышей дикого типа и Fh2-дефицитных клеток и для прогнозирования дальнейших нокаутов генов, которые влияют на рост Fh2- дефицитные клетки, но не влияют на клетки дикого типа, что позволяет предположить мишени для лечения наследственного лейомиоматоза и почечно-клеточного рака [153].
Для конкретных заболеваний тканеспецифичные модели успешно используются для идентификации биомаркеров и терапевтических мишеней [125]. Использование уровней экспрессии RNA-Seq в сочетании с моделями в масштабе генома позволило реконструировать модели, специфичные для линий раковых клеток, что сделало возможным открытие метаболитов, поддерживающих пролиферацию, и антиметаболитов, ведущих к гибели клеток [154]. (Антиметаболит — это соединение, которое одновременно ингибирует ферменты, участвующие в метаболизме связанного эндогенного метаболита.Антиметаболиты могут влиять на несколько ферментов одновременно и могут снижать пролиферацию и поэтому используются в качестве противоопухолевых препаратов.) После интеграции с внеклеточными метаболомными данными и транскриптомными профилями, метаболическое моделирование надежно охарактеризовало внутриклеточный метаболизм клеточных линий лимфобластного лейкоза [155]. Комбинация различных данных omics и разделения потоков была использована для создания специфичных для рака моделей панели NCI60, достигая корреляции, близкой к 1 при прогнозировании оставшихся скоростей потока [13].
Следуя тому же направлению, персонализированные модели (например, использующие данные пациентов для ограничения общей реконструкции) обещают стать ключевыми для точной медицины (рис. 2). На сегодняшний день крупнейшим исследованием моделей, ориентированных на пациентов, является Атлас патологии человека [77]. Авторы построили персонализированные модели рака в масштабе генома у каждого пациента в 17 тканях. Это позволило исследовать метаболические различия между разными видами рака, а также биологические функции пациента.В другом недавнем исследовании [156] было набрано 86 пациентов с неалкогольной жировой болезнью печени, и их персонализированные метаболические модели в масштабе генома гепатоцитов были построены с использованием экспериментальных данных о потоках липопротеинов для конкретных пациентов. Новый молекулярный механизм заболевания был выяснен в результате персонализированного метаболического моделирования.
Генотипирование в сочетании с метаболическим моделированием для конкретного пациента открывает новые возможности для персонализированной медицины. Фактически, новые биомаркеры могут быть предсказаны индивидуально для каждого пациента, и поэтому могут быть разработаны индивидуальные методы лечения и впоследствии оценены с точки зрения их метаболических механических эффектов [157].Как показано в недавнем случае дефицита аргиназы (нарушение цикла мочевины), разные люди могут по-разному реагировать на одно и то же заболевание и лечение, и это часто можно отметить, наблюдая за их индивидуальной метаболической реакцией [158].
Из-за важности состава микробиома кишечника для здоровья человека недавно был предложен набор подходов к моделированию [159]. Используя моделирование микробиома хозяина, например, комбинируя моделирование метаболического сообщества с моделями метаболизма человека, можно исследовать взаимодействие между кишечными бактериями и их взаимодействие с окружающими человеческими клетками [160, 161].Персонализированные модели микробиома кишечника были построены на основе метагеномных данных, специфичных для каждого пациента, с целью выяснения индивидуального производства желчных кислот в микробиомах здоровых людей и пациентов с воспалительными заболеваниями кишечника [162]. Хотя некоторые проблемы моделирования еще предстоит решить [163], подход, основанный на метаболическом моделировании, вероятно, прольет свет на роль кишечных микробных сообществ человека в здоровье человека, что предполагает возможные диетические изменения или индивидуальное вмешательство в состав кишечника [164, 165 ].
5. Обсуждение и перспективы
Исследования в области вычислительной биологии привели к детальным моделям для лучшего понимания отдельных биологических компонентов, но, возможно, к менее четкой картине взаимодействий между компонентами, которые приводят к данному фенотипу [166] . Системная биология в масштабе генома может эффективно решить эту проблему. Например, в биомедицинских приложениях этот целостный взгляд необходим для характеристики фенотипа болезни пациента и предоставления точной медицины [17].
Поскольку метаболический гомеостаз и наблюдаемый фенотип строго связаны, метаболизм в настоящее время считается диагностическим признаком фенотипа и, следовательно, возможно, лучшим индикатором функционального состояния клетки. Метаболизм также можно использовать для определения приоритетов генов и оценки их функции и роли генных нарушений (включая нокауты). Без такого комплексного анализа ген может, например, ошибочно считаться важным только из-за его сильно изменчивого значения экспрессии.
Метаболические модели все чаще используются для построения многомасштабных, многоклеточных или многотканевых моделей.В 2012 году исследование Karr et al. [21] представили первую вычислительную модель целой клетки жизненного цикла небольшой патогенной бактерии, Mycoplasma genitalium . Модель включает метаболизм, репликацию генома и деление клеток. Несколько метаболических моделей могут быть объединены в рамки для исследования метаболических обменов между отдельными клетками и формирующегося поведения сообщества [167], с различными применениями, от микробных сообществ до взаимодействий хозяин-патоген и пролиферации рака [168–170].
Доступность тканеспецифичных моделей также привела к созданию многотканевых метаболических моделей [171]. Дальнейшие шаги в этом направлении вскоре позволят изучать модели метаболизма всего тела. В первом исследовании, направленном на достижение этой цели, использовалась динамическая экономная FBA для комбинирования стационарной FBA с дифференциальными уравнениями для концентрации метаболитов в каждом органе [172]. В результате была создана многомасштабная модель всего тела, включающая 14 органов плюс человеческая сыворотка. Совсем недавно были созданы первые метаболические модели всего тела, названные Харви и Харветта, где метаболическая сеть для каждого органа / ткани была построена одновременно из Recon3D [173].Следующим шагом в этом направлении кажется производство устройств «орган на кристалле» и, в конечном итоге, «тело на кристалле» с метаболическими моделями (см., Например, [174]).
Учитывая недавно возобновившийся интерес научного сообщества к пониманию метаболизма болезней, весьма вероятно, что метаболизм рака (и метаболизм при других заболеваниях) станет основной темой исследований при разработке лекарств. Это будет дополнять, а не заменять стандартные исследования, посвященные только транскриптоме, и обеспечит прокси для фенотипических и наблюдаемых результатов.Идея состоит в том, что для улучшения фенотипических прогнозов необходимо брать подписи из источников данных, которые ближе к фенотипу. Интеграция данных омики в метаболические модели позволила прогнозировать и последовательно подтверждать биомаркеры и терапевтические цели. В дизайне лекарств, хотя геномика, транскриптомика и протеомика часто считаются достаточными, метаболомика может прояснить механизмы, которые не видны по активности генов и белков. Что еще более важно, он может касаться случаев, когда гены, ответственные за заболевание, хорошо известны, но не поддаются лечению, но их соответствующие последующие реакции поддаются [175].
Геномический анализ обычно позволяет идентифицировать известные болезни через генные мутации. Однако он может быть не в состоянии пометить варианты известных заболеваний или идентифицировать новые заболевания. Это может произойти, например, если (i) генная мутация не отмечена как важная; (ii) ген вообще не проверяется; (iii) заболевание не является прямым следствием нарушения гена; (iv) люди по-разному реагируют на одну и ту же мутацию [158]. Модели в масштабе генома могут прояснить механизмы действия лекарств, побочные эффекты (как связывание лекарств вне мишени, так и последующие транскрипционные эффекты) и потенциальную токсичность лекарств путем связывания данных омики с фенотипом через модель, специфичную для состояния [8, 176] .В рамках индивидуального подхода к пациенту такие биологические данные и маркеры могут быть предсказаны индивидуально, что открывает путь для клинических испытаний in silico, [177].
Усилия сообщества метаболического моделирования далеки от завершения. В связи с этим мы предполагаем совместные усилия сообществ по моделированию метаболизма и полногеномных ассоциаций (GWAS) для выявления взаимодействий между генами и метаболитами, которые в настоящее время не включены в метаболические модели с использованием современных карт ассоциаций [178].Вычислительные инструменты, сочетающие оба подхода, будут способствовать обнаружению ошибок и пропущенных ферментативных реакций на уровне генома, что значительно улучшит предсказательную способность метаболических моделей. Например, это может быть достигнуто за счет минимизации ошибки между предсказаниями магнитной связи и экспериментальными данными коэкспрессии, интегрированными с GWAS [179, 180].
Точно так же методы интеграции данных omics в модели в масштабе генома все еще нуждаются в улучшении. Напр., В дрожжах и Escherichia coli , одномерная интеграция в FBA, как сообщается, дает аналогичную точность pFBA без омной интеграции [78].Обнадеживает то, что специфические для реакции правила ограничения скорости потока могут вдвое снизить нормированную ошибку pFBA [84]. В многоклеточных организмах можно ожидать лучшей корреляции между экспрессией генов и метаболическими потоками [78]. В клетках млекопитающих основной вклад в общий уровень экспрессии белка вносят уровни мРНК [181, 182], тогда как в большинстве нормальных и раковых клеточных линий уровни мРНК и белка положительно коррелируют [183, 184]. В то время как разные методы (и их параметры) могут давать разные контекстно-зависимые модели, точность прогнозирования основных генов почти всегда выше в контекстно-зависимых моделях, чем в общих человеческих моделях [81].Таким образом, метаболические модели можно рассматривать как полезные инструменты для механической связи транскриптомных данных со скоростью потока. Одновременное использование нескольких данных omics и термодинамических ограничений [185, 186] или комбинации регуляризованных методов FBA и данных omics может повысить надежность прогнозов [187].
По сравнению с хорошо изученными микробными метаболическими реконструкциями, пробелы в наших знаниях о метаболизме человека, включая характеристики ферментов, определение клеточно-специфических метаболических функций и тканеспецифических механизмов роста, затрудняют тестирование (и сравнить) метаболические модели и методы омической интеграции.Человеческие метаболические модели и методы были протестированы и перекрестно сравнены на предмет существенности генов с помощью CRISPR-Cas9-опосредованного скрининга потери функции, а также на их способность предсказывать скорость роста и воспроизводить известные метаболические функции [81]. Набор данных, включающий как данные omics, так и измеренную информацию о метаболизме, представляет собой панель NCI-60 с показателями поглощения и секреции метаболитов, опубликованными в 2012 году [188]. Хотя данные о E. coli и флуксомные дрожжи общедоступны из нескольких экспериментов [189–191], насколько нам известно, NCI-60 является единственным общедоступным набором данных о клетках человека, содержащем уровни экспрессии, скорости метаболического потока и скорости пролиферации. , и поэтому подходит для проверки методов FBA.Из-за отсутствия наборов флюксомных данных, сложности выбора надежной целевой функции и большего размера по сравнению с бактериальными моделями, требуется дальнейшая экспериментальная проверка метаболических моделей человека, особенно при использовании полного распределения потока для информирования решений или последующих алгоритмических шагов. .
Наконец, несмотря на многие недавние достижения, сбор информации на основе данных, полученных с помощью моделей, основанных на атомной информации, остается узким местом в системной биологии. Поэтому мы предполагаем, что метаболические модели в масштабе генома будут все больше исследоваться с помощью алгоритмов машинного / глубокого обучения с учетом специфики пациента [13, 192].При моделировании болезни, полезность общего обнаружения биомаркер является дискуссионной, когда информация о пациентах, не принимается во внимание, потому что большинство биологических компонентов будут помечены как пошевелить в общей «болезни по сравнению с нормальным» анализом. И наоборот, если соответствующие индивидуальные данные интегрированы в модель, биомаркеры могут стать центральной частью точной медицины, где данные, полученные о пациентах, определяют прогнозы, анализ и лечение [17]. По аналогии с моделированием, алгоритмы машинного обучения сами по себе не могут предоставить механистическую информацию о биологических процессах, которые они имитируют или имитируют.Однако, если машинное обучение сочетается с многомерным моделированием в масштабе генома, комбинация экспериментальных и созданных моделями атомных данных может предсказывать — и механистически объяснять — персонализированные прогнозы терапии путем включения ключевой биологической информации в процесс обучения.
Конфликт интересов
Автор заявляет, что у него нет конфликта интересов.
Дополнительные материалы
Согласие между теоретическими и модельными выходами АТФ для самых последних моделей метаболизма человека.Мы рассчитываем среднеквадратичную ошибку (RMSE) между теоретическим и прогнозируемым моделями максимальным выходом АТФ (на единицу источника углерода) для самых последних моделей метаболизма человека. RMSE улучшился с ∞ (Recon 2 и более ранние модели) до 5,17 (в Recon 3) и 1,67 (в Recon 2.2). (Дополнительные материалы)
Ученые говорят: метаболизм | Новости науки для студентов
калорий Количество энергии, необходимое для повышения температуры 1 грамма воды на 1 градус Цельсия.Обычно он используется для измерения энергии, содержащейся в определенном количестве пищи.
клетка Наименьшая структурная и функциональная единица организма. Обычно она слишком мала, чтобы ее можно было увидеть невооруженным глазом, она состоит из водянистой жидкости, окруженной мембраной или стенкой. В зависимости от размера животные состоят из тысяч или триллионов клеток. Большинство организмов, таких как дрожжи, плесень, бактерии и некоторые водоросли, состоят только из одной клетки.
химический Вещество, состоящее из двух или более атомов, которые объединяются (связываются) в фиксированной пропорции и структуре.Например, вода — это химическое вещество, которое образуется, когда два атома водорода связываются с одним атомом кислорода. Его химическая формула — H 2 O. Химический также может быть прилагательным для описания свойств материалов, которые являются результатом различных реакций между различными соединениями.
химическая реакция Процесс, который включает перегруппировку молекул или структуры вещества в противоположность изменению физической формы (например, от твердого тела к газу).
ДНК (сокращение от дезоксирибонуклеиновой кислоты) Длинная, двухцепочечная и спиралевидная молекула внутри большинства живых клеток, несущая генетические инструкции.Он построен на основе атомов фосфора, кислорода и углерода. Эти инструкции сообщают клеткам, какие молекулы должны образовывать все живые существа, от растений и животных до микробов.
ферментов Молекулы, производимые живыми существами для ускорения химических реакций.
метаболизм (прил. Метаболизм) Набор поддерживающих жизнь химических реакций, которые происходят внутри клеток и более крупных структур, таких как органы. Эти реакции позволяют организмам расти, воспроизводиться, двигаться и иным образом реагировать на окружающую их среду.
молекула Электрически нейтральная группа атомов, представляющая минимально возможное количество химического соединения. Молекулы могут состоять из атомов одного или разных типов. Например, кислород в воздухе состоит из двух атомов кислорода (O 2 ), а вода состоит из двух атомов водорода и одного атома кислорода (H 2 O).
орган (в биологии) Различные части организма, которые выполняют одну или несколько определенных функций.Например, яичник — это орган, производящий яйца, мозг — это орган, воспринимающий нервные сигналы, а корни растений — это органы, которые поглощают питательные вещества и влагу.
белок Соединение, состоящее из одной или нескольких длинных цепочек аминокислот. Белки — неотъемлемая часть всех живых организмов. Они составляют основу живых клеток, мышц и тканей; они также выполняют работу внутри клеток. Среди наиболее известных автономных белков — гемоглобин (в крови) и антитела (также в крови), которые пытаются бороться с инфекциями.Лекарства часто действуют путем фиксации белков.
теплокровных Прилагательное для животных (в основном млекопитающих и птиц), которые поддерживают постоянную температуру тела, обычно выше температуры окружающей их среды. Ученые обычно предпочитают термин эндотермический для описания животных, которые выделяют тепло для контроля температуры своего тела.
отходы Любые материалы, оставшиеся от биологических или других систем, не представляющие ценности, поэтому их можно утилизировать как мусор или переработать для нового использования.
|
Обнаружена ошибка PHP
Уровень серьезности: Уведомление
Сообщение: Попытка получить свойство не-объекта
Имя файла: helpers / menu_helper.php
Номер строки: 271
Отслеживание:
Файл: /home/ashok191/public_html/heraldopenaccess.us/application/helpers/menu_helper.php
Строка: 271
Функция: _error_handler
Файл: / home / ashok191 / public_html / heraldopenaccess.us / application / controllers / journals / Home.php
Строка: 127
Функция: get_menuname_bylink
Файл: /home/ashok191/public_html/heraldopenaccess.us/index.php
Строка: 630
Функция: require_once
Обнаружена ошибка PHP
Уровень серьезности: Уведомление
Сообщение: Попытка получить свойство не-объекта
Имя файла: includes / header.php
Номер строки: 15
Отслеживание:
Файл: / home / ashok191 / public_html / heraldopenaccess.us / application / views / journals / includes / header.php
Строка: 15
Функция: _error_handler
Файл: /home/ashok191/public_html/heraldopenaccess.us/application/controllers/journals/Home.php
Строка: 129
Функция: просмотр
Файл: /home/ashok191/public_html/heraldopenaccess.us/index.php
Строка: 630
Функция: require_once
Обнаружена ошибка PHP
Уровень серьезности: Уведомление
Сообщение: Попытка получить свойство не-объекта
Имя файла: включает / заголовок.php
Номер строки: 15
Отслеживание:
Файл: /home/ashok191/public_html/heraldopenaccess.us/application/views/journals/includes/header.php
Строка: 15
Функция: _error_handler
Файл: /home/ashok191/public_html/heraldopenaccess.us/application/controllers/journals/Home.php
Строка: 129
Функция: просмотр
Файл: / home / ashok191 / public_html / heraldopenaccess.us / index.php
Строка: 630
Функция: require_once
Обнаружена ошибка PHP
Уровень серьезности: Уведомление
Сообщение: Попытка получить свойство не-объекта
Имя файла: journals / contentpage.php
Номер строки: 10
Отслеживание:
Файл: /home/ashok191/public_html/heraldopenaccess.us/application/views/journals/contentpage.php
Строка: 10
Функция: _error_handler
Файл: / home / ashok191 / public_html / heraldopenaccess.us / application / controllers / journals / Home.php
Строка: 130
Функция: view
Файл: /home/ashok191/public_html/heraldopenaccess.us/index.php
Строка: 630
Функция: require_once
Обнаружена ошибка PHP
Уровень серьезности: Уведомление
Сообщение: Попытка получить свойство не-объекта
Имя файла: journals / contentpage.php
Номер строки: 10
Отслеживание:
Файл: / home / ashok191 / public_html / heraldopenaccess.