Урок 2. природа и рукотворный мир — Окружающий мир — 2 класс
Окружающий мир, 2 класс
Урок 2. «Природа и рукотворный мир»
Перечень вопросов, рассматриваемых на уроке:
- Что такое «природа».
- Что такое «рукотворный мир».
Глоссарий по теме:
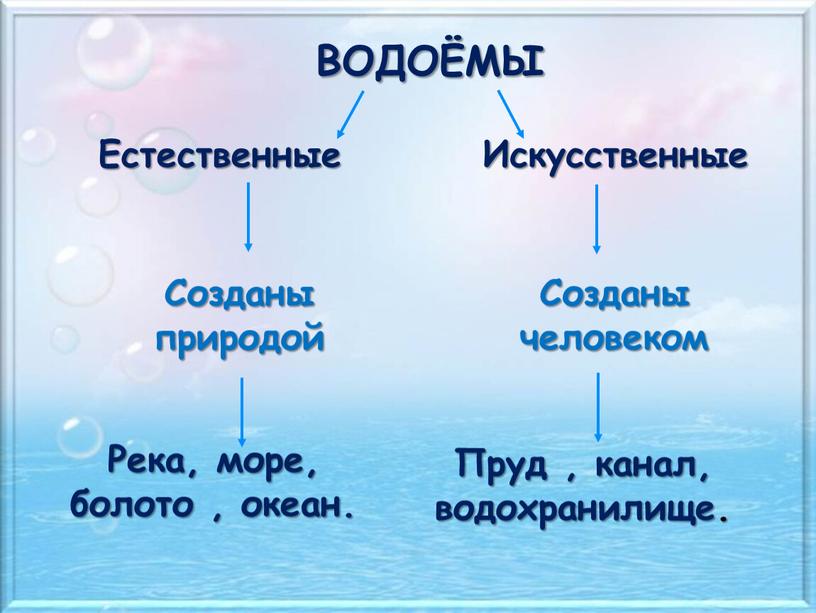
Природа – это то, что нас окружает.
Рукотворный мир – то, что создано людьми.
Экология – наука, изучающая взаимодействия между организмами и окружающей средой.
Основная и дополнительная литература по теме урока:
- Окружающий мир. Рабочая тетрадь. 2 кл.: учеб. пособие для общеобразоват. организаций. В 2 ч. / А. А. Плешаков. – М.: Просвещение, 2017. С. 14–16.
- Атлас – определитель «От земли до неба» С. 9, 15; 20; 120; 208.
- Пособие для учащихся общеобразовательных учреждений «Великан на поляне или первые уроки экологической этики». А.А. Плешаков, А.А.Румянцев. М: Просвещение,2012. С. 3–4.
- Плешаков А.


Теоретический материал для самостоятельного изучения
Всё, что нас окружает – это окружающий мир: объекты природы и предметы, которые изобрели люди. Природа – это всё, что нас окружает, что НЕ сделано руками человека. Но есть и предметы, созданными людьми. Вот и получается, что окружающий мир делится две группы.
Первая группа – объекты природы. Эти объекты существуют в природе и не зависят от человека.
Вторая группа – это предметы, сделанные руками человека. Вот поэтому и называем мы эту группу – рукотворный мир, т.е. руки человека сотворили. Всё, что нас окружает: солнце, растения, животные, вода, воздух, горы, камни, облака, луна, звёзды – это одним словом называется «природа». Она существует независимо от человека. Человек – тоже часть природы. Мир, созданный человеком, огромный. Человек окружил себя множеством предметов, которые помогают ему удобно жить, работать, учиться, отдыхать, передвигаться.
Наша планета очень велика в сравнении с каждым человеком, а он так мал в сравнении с ней! Но в то же время, человек, идущий по лесной поляне, опушке, лугу, превращается в огромного великана. Во-первых, потому что на полянах и опушках живут существа, которые меньше человека: жуки и муравьи, бабочки и шмели, гусеницы и пауки. Во-вторых, потому что даже большие обитатели Земли, например, вековые дубы перед человеком беззащитны. Причём могущественным Великаном на поляне может быть не только взрослый, но и маленький ребёнок. Как хорошо, что этот Великан добрый. Он не наступит на муравейник, не обидит гусеницу, не сорвет цветок, не оставит мусор после себя. И как печально, если великан окажется недобрым, равнодушным, жестоким к тому, что его окружает».
Наш окружающий мир удивительно красив. И очень важно правильно обращаться с такой красотой. Нужно беречь всё, что дает нам природа. Нужно сохранить наш мир зелёным и чистым. Ведь природа – наш зелёный дом, и нужно стараться, чтобы в доме всегда было чисто и красиво.
Примеры и разбор решения заданий
- Выберите объекты природы.
Правильный вариант ответа:
2. Подчеркните верные утверждения:
Не разоряйте муравейники!
Не ловите бабочек, стрекоз, жуков!
Приносите домой детёнышей лесных зверей и птиц!
Не ломайте ветки деревьев!
Правильный вариант/варианты:
Не разоряйте муравейники!
Не ловите бабочек, стрекоз, жуков!
Не ломайте ветки деревьев!
Изучая связь природы и искусства
Один из наиболее выдающихся художников всех времен Анри Матисс говорил: «Художник должен обладать природой. Он должен идентифицировать себя со своим ритмом, мерами, которые подготовят мастерство, позже позволящее ему выражать себя на его собственном языке».
Сколько существует искусство, художники всегда были очарованы природой. Помимо бесконечного источника вдохновения, для создания своих шедевров они всегда использовали природные средства, такие как дерево, уголь, глина, графит и вода.
Художники минувших эпох
Хотя при жизни Винсент ван Гог продал всего одну свою картину, он был единственным в своем роде. Он умел передавать такие аспекты природы, как простые цветы, подарив им новую жизнь на своих полотнах. Одна из таких работ – «Ирисы», особенно впечатляет своей жизненной силой, ведь цветы можно практически почувствовать. Еще один мировой гений художественного искусства – Моне, черпал вдохновение из мира природы. Серия его работ «Водяные лилии» – потрясающий пример безупречной игры теней, света и воды, а также «портрет» его сада во Франции. Цветы были одной из основных тем работ Моне последние 30 лет его жизни, идеально иллюстрируя, насколько огромным является влияние красоты природы на нас, а также на воображение людей искусства.
Современные художники, вдохновленные природой
Мэри Иверсон живет и работает в Сиэтле, штат Вашингтон. Она черпает вдохновение в окружающей ее невероятной красоте природы. Ее потрясающие картины предлагают довольно современный взгляд на традиционное искусство пейзажей: Мэри рисует известные памятники и национальные парки США. Больше всего ее вдохновляли такие места, как порт Сиэтла и гора Рейнер, национальные парки Норт-Каскейдс и Олимпийский национальный парк. Работы Мэри появлялись на обложках журнала Juxtapoz в 2015 году, а также на страницах газеты Хаффингтон Пост, Бостон Ревью и журнала Зарубежная политика. Художница тесно сотрудничает с рядом галерей в Германии, Франции, Нидерландах и США, а также преподает визуальные искусства в Колледже долины Скагит в районе горы Вернон, где она с нескрываемым энтузиазмом делится своей любовью к природе со своими студентами.
Ее потрясающие картины предлагают довольно современный взгляд на традиционное искусство пейзажей: Мэри рисует известные памятники и национальные парки США. Больше всего ее вдохновляли такие места, как порт Сиэтла и гора Рейнер, национальные парки Норт-Каскейдс и Олимпийский национальный парк. Работы Мэри появлялись на обложках журнала Juxtapoz в 2015 году, а также на страницах газеты Хаффингтон Пост, Бостон Ревью и журнала Зарубежная политика. Художница тесно сотрудничает с рядом галерей в Германии, Франции, Нидерландах и США, а также преподает визуальные искусства в Колледже долины Скагит в районе горы Вернон, где она с нескрываемым энтузиазмом делится своей любовью к природе со своими студентами.
Британские художники всю жизнь находят вдохновение в природе
Британский художник дикой природы Джонатан Сэнсбури известен своим потрясающим талантом запечатлевать мимолетные сцены из жизни дикой природы. Наблюдая и рисуя различные сцены большую часть своей жизни, Сэнсбури стал мастером акварельной живописи и стиля, в котором объединены техника рисунка углем и акварелью, вдыхающие в его работы чувство настоящего движения. Помимо того, что он умеет запечатлеть саму суть бесчисленных сцен из природы, Сэнсбури удается писать природу в метафорическом срезе, делая ссылки на нашу повседневную жизнь. Работами Джонатана можно полюбоваться в Галерее Викхем в Стокбридже, Галерее Стратхерн и выставочном центре искусств Данкельд.
Помимо того, что он умеет запечатлеть саму суть бесчисленных сцен из природы, Сэнсбури удается писать природу в метафорическом срезе, делая ссылки на нашу повседневную жизнь. Работами Джонатана можно полюбоваться в Галерее Викхем в Стокбридже, Галерее Стратхерн и выставочном центре искусств Данкельд.
Несмотря на то что с каждым днем наш мир становится все более и более технологичным, и все же в нем существует не так много того, что может вдохновить гениев искусства так, как это удается природе. От лепестка розы, плавно падающего на землю, и до мощного орлана, падающего камнем в погоне за своей добычей, – все эти проявления природы не перестанут завораживать и дарить вдохновение, превращаясь в выдающиеся творения искусства.
Отдых на природе организовали для пожилых людей в Уйском районе
С возрастом у людей появляется всё больше проблем со здоровьем. Иногда, не обойтись без лечебного и, одновременно, оздоровительного отдыха в соответствующем учреждении. К сожалению, многие пенсионеры отказывают себе в подобном удовольствии ввиду того, что считают, что пребывание в санатории — дорогое удовольствие. Уединение с природой бывает для них нечастая радость, да и не каждый из них может себе это позволить.
К сожалению, многие пенсионеры отказывают себе в подобном удовольствии ввиду того, что считают, что пребывание в санатории — дорогое удовольствие. Уединение с природой бывает для них нечастая радость, да и не каждый из них может себе это позволить.
В этом году Комплексный центр социального обслуживания населения очередной раз организовал десятидневный бесплатный отдых на природе 55 гражданам пожилого возраста. Оздоровительный отдых был организован на средства целевой программы «Развитие социальной защиты населения в Уйском муниципальном районе на 2018–2020 годы» подпрограмма «Повышение качества жизни граждан пожилого возраста в Уйском муниципальном районе».
Полноценное четырехразовое питание, спортивные мероприятия, занятия по интересам, танцевальные вечера, мероприятия на воздухе, концертные программы и лекции и многое другое, чем занимались отдыхающие ветераны.
«Погода радовала — спокойствие и благодать! Как нам в жизни этого не хватает, особенно пожилому человеку. Что себе может позволить обычный российский пенсионер? Копаться на грядках, чтобы прожить до следующей пенсии, а тут бесплатно отдохнули и набрались сил», — делится впечатлениями Любовь Мурзина. «Наши «бабули» и «дедули» получили огромный заряд и положительные эмоции. Хоть на время забылись печаль, проблемы, боль, одиночество», — добавила медсестра Светлана Стругова. Домой ехали с песнями, хорошим настроением и со словами благодарности, немного грустные с надеждой встретится в следующим году!
Что себе может позволить обычный российский пенсионер? Копаться на грядках, чтобы прожить до следующей пенсии, а тут бесплатно отдохнули и набрались сил», — делится впечатлениями Любовь Мурзина. «Наши «бабули» и «дедули» получили огромный заряд и положительные эмоции. Хоть на время забылись печаль, проблемы, боль, одиночество», — добавила медсестра Светлана Стругова. Домой ехали с песнями, хорошим настроением и со словами благодарности, немного грустные с надеждой встретится в следующим году!
«Ничто не может заменить для человека оздоровительного влияния природы. Красота родного края, чистый насыщенный кислородом и сосновым запахом воздух, единение с природой, приятное времяпровождение среди друзей – все это лечит нервную систему лучше лекарств. Выход из четырех стен, общение с природой, двигательная активность оказывают на пожилого человека все виды оздоровления. Это главная цель национального проекта «Демография», регионального проекта «Старшее поколение»»,
— комментирует директор центра Ольга Леонтьева.
Профилактика клещевого вирусного энцефалита — ДЗМ
Весеннее пробуждение природы сопровождается подъёмом биологической активности животных, птиц и членистоногих, которые являются источниками либо переносчиками различных инфекционных заболеваний. Риск заражения человека отдельными инфекционными болезнями при тесном контакте с природой многократно увеличивается.
Одной из больших опасностей для москвичей за городом являются клещи – переносчики целого ряда инфекционных заболеваний, в том числе самого опасного из них — клещевого вирусного энцефалита (КВЭ). Клещевой вирусный энцефалит – это острое вирусное инфекционное заболевание, характеризующееся лихорадкой, интоксикацией, поражением вещества и оболочек головного и спинного мозга. Исход заболевания зависит от объёма поражения центральной нервной системы. В 25-30% случаев заболевание заканчивается смертью больного. Также возможно развитие серьезных осложнений (параличей, эписиндрома, снижения интеллекта), приводящих к инвалидизации человека.
Где можно заразиться клещевым вирусным энцефалитом?
Ареал обитания клещей – переносчиков клещевого вирусного энцефалита – достаточно широк и захватывает многие административные территории Российской Федерации. За природными территориями, населёнными клещами, ведётся плановое наблюдение с исследованием клещей на наличие вируса клещевого энцефалита. Отдельные территории являются благополучными по КВЭ, в других – при исследовании клещей регулярно выделяется вирус клещевого энцефалита. Территории, где обитают клещи, заражённые вирусом, определены как эндемичные.
С перечнем территорий нашей страны, эндемичных по клещевому вирусному энцефалиту, можно ознакомиться на сайте Федеральной службы по надзору в сфере защиты прав потребителей и благополучия человека: https://rospotrebnadzor.ru/deyatelnost/epidemiological-surveillance/?ELEMENT_ID=13603
Эндемичные районы Московской области по клещевому энцефалиту -Дмитровский и Талдомский.
Начало активности клещей совпадает с появлением в лесу первых проталин, поэтому уже в апреле возможны первые укусы.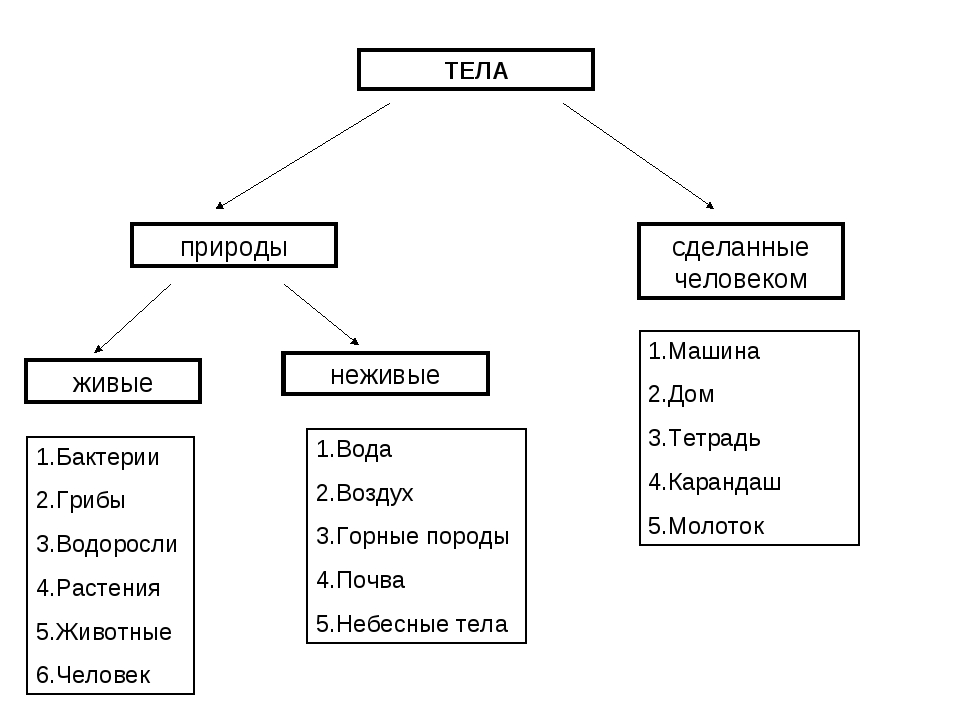 С каждым днём количество активных клещей увеличивается, достигая пика в мае, и остается высокой до середины или конца июня. Поэтому наиболее опасны выезды на природу в майские праздники.
С каждым днём количество активных клещей увеличивается, достигая пика в мае, и остается высокой до середины или конца июня. Поэтому наиболее опасны выезды на природу в майские праздники.
К заражению КВЭ восприимчивы все люди, независимо от возраста и пола.
Как можно защититься от клещевого энцефалита?
Наиболее эффективным средством защиты от клещевого вирусного энцефалита является вакцинация. Всем лицам, планирующим выезд в эндемичные по клещевому энцефалиту территории, необходимо заранее сделать профилактические прививки против клещевого энцефалита.
Вакцинация проводится двукратно с минимальным интервалом между первой и второй прививками 1 месяц и ревакцинацией через год. Последующие ревакцинации проводятся каждые 3 года. Для выработки иммунитета требуется не менее двух недель, поэтому завершить прививочный курс (две прививки) против клещевого энцефалита необходимо за 2 недели до выезда в неблагополучную территорию.
Где и когда можно получить профилактические прививки против КВЭ?
Вакцинация осуществляется бесплатно в медицинских организациях по месту прикрепления.
Вакцинация проводится круглогодично, без календарных ограничений, т.е. если летом вы планируете поездку в эндемичный район, то уже зимой можно пройти вакцинацию против клещевого энцефалита в поликлинике по месту прикрепления.
Прививаются:
Лица, проживающие на эндемичных по клещевому вирусному энцефалиту территориях; лица, выезжающие на эндемичные по клещевому вирусному энцефалиту территории, а также прибывшие на эти территории лица, выполняющие следующие работы:
— сельскохозяйственные, гидромелиоративные, строительные, по выемке и перемещению грунта, заготовительные, промысловые, геологические, изыскательские, экспедиционные, дератизационные и дезинсекционные;
— по лесозаготовке, расчистке и благоустройству леса, зон оздоровления и отдыха населения.
Лица, работающие с живыми культурами возбудителя клещевого энцефалита.
Приказ Минздрава от 21032014 125н.pdf
Позаботьтесь о своём здоровье заранее!
Что делать и куда обращаться, если произошло присасывание клеща?
Не следует самостоятельно удалять клеща, необходимо немедленно обратиться в ближайший травматологический пункт. Там вам окажут квалифицированную медицинскую помощь и решат вопрос о необходимости проведения экстренной серопрофилактики (введение иммуноглобулина против клещевого энцефалита).
Там вам окажут квалифицированную медицинскую помощь и решат вопрос о необходимости проведения экстренной серопрофилактики (введение иммуноглобулина против клещевого энцефалита).
Экстренная серопрофилактика КВЭ проводится в случае присасывания клеща на эндемичной территории лицам:
— не привитым против КВЭ или получившим неполный курс вакцинации;
— привитым в случае повышенного риска заражения (выявлено инфицирование присосавшегося клеща, многократные укусы или одновременное присасывание нескольких клещей).
Введение иммуноглобулина необходимо провести как можно раньше, но не позднее 4-го дня (96 часов) после присасывания клеща.
В каких медицинских организациях Москвы проводится экстренная серопрофилактика клещевого энцефалита?
Экстренная серопрофилактика клещевого энцефалита в Москве проводится круглосуточно:
- взрослым — на базе Городского консультативного кабинета по вакцинно-сывороточной профилактике клещевого вирусного энцефалита в ГБУЗ «Инфекционная клиническая больница № 2 ДЗМ» по адресу: г. Москва, 8-я ул. Соколиной горы, д. 15, тел. 8(495)366-84-68, 8(49)365-01-47, 8(495)365-19-28
- детям — на базе ГБУЗ «Детская клиническая больница № 13
им. Н. Ф. Филатова ДЗМ» по адресу: г. Москва, ул. Садовая-Кудринская, д. 15, тел.: 8(499)254-34-30.
 Москва, 8-я ул. Соколиной горы, д. 15, тел. 8(495)366-84-68, 8(49)365-01-47, 8(495)365-19-28
Москва, 8-я ул. Соколиной горы, д. 15, тел. 8(495)366-84-68, 8(49)365-01-47, 8(495)365-19-28Получить дополнительную информацию, а также проконсультироваться по вопросам клещевого вирусного энцефалита и укусам клещей можно в Городском консультативном кабинете по вакцинно-сывороточной профилактике клещевого вирусного энцефалита ГБУЗ «Инфекционная клиническая больница № 2 ДЗМ» по адресу: г. Москва, 8-я ул. Соколиной горы, д. 15, тел.: 8 (495) 366-84-68,
8(495)365-59-47.
Если нет возможности сразу обратиться в травматологический пункт для удаления присосавшегося клеща, и вы самостоятельно смогли его удалить, сохраните его и доставьте в лабораторию для исследования. Это важно сделать, так как при укусе клещом кроме клещевого энцефалита можно заразиться клещевым боррелиозом. Вакцины для профилактики клещевого боррелиоза не существует. Если от присосавшегося клеща выделен возбудитель боррелиоза, то необходимо обратиться к врачу для дальнейшего назначения лечения.
Если от присосавшегося клеща выделен возбудитель боррелиоза, то необходимо обратиться к врачу для дальнейшего назначения лечения.
 It has been emphasized that environmental problems are not limited to the problem of childhood, environmental security, which in themselves are paradoxical (the need to protect people from the consequences of the environmental crisis generated by them). The problems make obvious the need to find causes of environmental problems in human culture and society, and their solution — in formation of a new cultural code based on interiorization by personalities and society of ecocultural values, their ecological culture.phenomenon of childhoodchildhood ecologyhuman ecologycultureeducationsocializationprofessional competence of teachersфеномен детстваэкология детстваэкология человекакультуравоспитаниесоциализацияпрофессиональная компетентность педагога1.Агаджанян Н. А. Экология души человека и природы. 2-е изд., доп.и перераб. М. : РУДН, 2012. 265 с.2.Анисимов О. С., Глазачев С. Н. Экологическая культура: сущность и пути формирования в рамках профессионального образования // Вестник Международной академии наук (Русская секция).
It has been emphasized that environmental problems are not limited to the problem of childhood, environmental security, which in themselves are paradoxical (the need to protect people from the consequences of the environmental crisis generated by them). The problems make obvious the need to find causes of environmental problems in human culture and society, and their solution — in formation of a new cultural code based on interiorization by personalities and society of ecocultural values, their ecological culture.phenomenon of childhoodchildhood ecologyhuman ecologycultureeducationsocializationprofessional competence of teachersфеномен детстваэкология детстваэкология человекакультуравоспитаниесоциализацияпрофессиональная компетентность педагога1.Агаджанян Н. А. Экология души человека и природы. 2-е изд., доп.и перераб. М. : РУДН, 2012. 265 с.2.Анисимов О. С., Глазачев С. Н. Экологическая культура: сущность и пути формирования в рамках профессионального образования // Вестник Международной академии наук (Русская секция). 2012. № 2. С. 5-17. URL: http://www.heraldrsias.ru/online/2012/2/230/ (дата обращения 03.05.2013)3.Вагнер (Цветкова) И. В. Экологическое воспитание младших школьников. М. : Педагогическое общество России, 2000. 275 с.4.Вагнер И. В., Глазачев О. С., Глазачев С. Н. Фактор культуры в проблемном поле экологии человека: информационное общество и экологическая культура // Вестник МГГУ им. М. А. Шолохова. Социально-экологические технологии. 2013. № 2. С. 21-37.5.Глазачев С. Н., Гагарин А. В. Экологическая культура как вершинное достижение личности // Акмеология. 2013. № 51 (47). С. 63-71.6.Глазачев С. Н. Гагарин А. В. Экологическая акмеология: о трендах современной науки // Акмеология. 2013. № 51(49). С. 149.7.Гришаева Ю. М. Концепция формирования экологопрофессиональной компетентности студентов гуманитарного вуза : автореф. дис.. д-ра пед. наук. Москва, 2014. 38 с.8.Гудков А. Б. Северное направление научной школы академика Н. А. Агаджаняна. К 85-летнему юбилею учёного // Экология человека. 2013.
2012. № 2. С. 5-17. URL: http://www.heraldrsias.ru/online/2012/2/230/ (дата обращения 03.05.2013)3.Вагнер (Цветкова) И. В. Экологическое воспитание младших школьников. М. : Педагогическое общество России, 2000. 275 с.4.Вагнер И. В., Глазачев О. С., Глазачев С. Н. Фактор культуры в проблемном поле экологии человека: информационное общество и экологическая культура // Вестник МГГУ им. М. А. Шолохова. Социально-экологические технологии. 2013. № 2. С. 21-37.5.Глазачев С. Н., Гагарин А. В. Экологическая культура как вершинное достижение личности // Акмеология. 2013. № 51 (47). С. 63-71.6.Глазачев С. Н. Гагарин А. В. Экологическая акмеология: о трендах современной науки // Акмеология. 2013. № 51(49). С. 149.7.Гришаева Ю. М. Концепция формирования экологопрофессиональной компетентности студентов гуманитарного вуза : автореф. дис.. д-ра пед. наук. Москва, 2014. 38 с.8.Гудков А. Б. Северное направление научной школы академика Н. А. Агаджаняна. К 85-летнему юбилею учёного // Экология человека. 2013. № 1. С. 3-7.9.Дерябо С. Д., Ясвин В. А. Экологическая педагогика и психология. Ростов-на-Дону, 1996. 480 с.10.Лихачев Б. Т. Философия воспитания. М. : Владос, 2010. 336 с.11.Панов В. И. Введение в экологическую психологию : учебное пособие. 2-е изд., перераб. и доп. М. : НИИ Школьных технологий, 2006. 184 с.12.Панов В. И. Экопсихологические аспекты детства // Мир психологии. 1997. № 1. С. 55-67.13.Реймерс Н. Ф. Экология (теории, законы, правила, принципы и гипотезы). М., 1994. 367 с.14.Ткачёва А. Н. Готовность современного учителя к воспитательной деятельности и развитие системы подготовки педагогов к организации воспитательного процесса // Социальная педагогика. 2012. № 2. С. 29-39.15.Учителю об экологии детства / под ред. В. Лебедевой, В. Панова. М. : РАО. Психологический институт. ЦКФЛ, 1996. 231 с.16.Шалаева С. Л. Детство в ситуации постмодерна // Ученые записки Казанского государственного университета. 2006. Т. 148. Кн. 1. С. 169-182.17.Шалаева С. Л. Диалектика детства в технологизированном социуме: аксиологический аспект // Вестник Нижегородского университета Н.
№ 1. С. 3-7.9.Дерябо С. Д., Ясвин В. А. Экологическая педагогика и психология. Ростов-на-Дону, 1996. 480 с.10.Лихачев Б. Т. Философия воспитания. М. : Владос, 2010. 336 с.11.Панов В. И. Введение в экологическую психологию : учебное пособие. 2-е изд., перераб. и доп. М. : НИИ Школьных технологий, 2006. 184 с.12.Панов В. И. Экопсихологические аспекты детства // Мир психологии. 1997. № 1. С. 55-67.13.Реймерс Н. Ф. Экология (теории, законы, правила, принципы и гипотезы). М., 1994. 367 с.14.Ткачёва А. Н. Готовность современного учителя к воспитательной деятельности и развитие системы подготовки педагогов к организации воспитательного процесса // Социальная педагогика. 2012. № 2. С. 29-39.15.Учителю об экологии детства / под ред. В. Лебедевой, В. Панова. М. : РАО. Психологический институт. ЦКФЛ, 1996. 231 с.16.Шалаева С. Л. Детство в ситуации постмодерна // Ученые записки Казанского государственного университета. 2006. Т. 148. Кн. 1. С. 169-182.17.Шалаева С. Л. Диалектика детства в технологизированном социуме: аксиологический аспект // Вестник Нижегородского университета Н. И. Лобачевского. Серия «Социальные науки». 2011. № 4 (24). С. 155-16018.Шульженко А. К. Эколого-эстетическое воспитание молодежи в европейской и отечественной педагогике : дис.. д-ра. пед. наук. Москва, 2006. 45 с.19.Экология человека в современном понимании // Экология человека. Информационный портал. URL: www. humanecology.ru/page0.htm (дата обращения: 03.05.2013).20.Этика / Т. В. Мишаткина, З. В. Бражникова, Н. И. Мушинский и др. ; под ред. Т. В. Мишаткиной, Я. С. Яскевич. 2-е изд. Минск : Новое знание, 2002. 494 с.21.Ясвин В. А. Психология отношения к природе. М. : Смысл. 2000. 456 с.22.Umweltethik zur Einführung (Broschiert) von Konrad Ott, Ernst Moritz Arndt Universität, 2010.23.Umweltethik, Ein Lehr- und Lesebuch von Andreas Brenner von Paulusverlag, Fribourg, 2008.24.Umweltethik: Manifest eines verantwortungsvollen Umgangs mit der Natur von Gerhard Pretzmann von Stocker, Stuttgart, 200125.Umweltethik: Wieder die Ökologische Krise: Ein kritischer Vergleich der Positionen von Vittorio Hösle und Hans Jonas von Alexander Klier von Tectum-Verlag, 2007.
И. Лобачевского. Серия «Социальные науки». 2011. № 4 (24). С. 155-16018.Шульженко А. К. Эколого-эстетическое воспитание молодежи в европейской и отечественной педагогике : дис.. д-ра. пед. наук. Москва, 2006. 45 с.19.Экология человека в современном понимании // Экология человека. Информационный портал. URL: www. humanecology.ru/page0.htm (дата обращения: 03.05.2013).20.Этика / Т. В. Мишаткина, З. В. Бражникова, Н. И. Мушинский и др. ; под ред. Т. В. Мишаткиной, Я. С. Яскевич. 2-е изд. Минск : Новое знание, 2002. 494 с.21.Ясвин В. А. Психология отношения к природе. М. : Смысл. 2000. 456 с.22.Umweltethik zur Einführung (Broschiert) von Konrad Ott, Ernst Moritz Arndt Universität, 2010.23.Umweltethik, Ein Lehr- und Lesebuch von Andreas Brenner von Paulusverlag, Fribourg, 2008.24.Umweltethik: Manifest eines verantwortungsvollen Umgangs mit der Natur von Gerhard Pretzmann von Stocker, Stuttgart, 200125.Umweltethik: Wieder die Ökologische Krise: Ein kritischer Vergleich der Positionen von Vittorio Hösle und Hans Jonas von Alexander Klier von Tectum-Verlag, 2007.
10 правил съемки дикой природы
1. Я уверен, что каждому фотографу необходим индивидуальный проект, который является для автора значимым, который нужно знать и раскрыть до конца. Необходима идея. Вообще фотография без идеи – это тупиковый путь. Кроме того, необходим внутренний «запал» и желание, которые будут двигать вперед. Когда в съемку вкладывается душа, то фотография получается и говорит сама за себя.
2. Важный момент для любого фотографа дикой природы: дикая природа в первую очередь, фотография – во вторую. Мы пришли на территорию, где животные – хозяева, они здесь главные. Уважать дикую природу – означает не принуждать животное совершать какие-либо действия или вредить ему. Позволяйте всему идти своим чередом…
3. Наблюдая за животным, можно снять момент, который длится всего секунду, который случайный человек не смог бы никогда увидеть. Чтобы получить хорошие фотографии, следует быть настойчивым и набраться терпения. Я довольно упрям и усидчив, и готов ждать долго. Именно так можно запечатлеть решающие моменты. Хорошие моменты стоят большого труда.
Я довольно упрям и усидчив, и готов ждать долго. Именно так можно запечатлеть решающие моменты. Хорошие моменты стоят большого труда.
4. Чтобы начать фотографировать, чтобы понимать, куда двигаться, нужно знать, от чего отталкиваться. А оттолкнуться можно только от хорошей фотографии. Надо хорошо знать, что вы хотите, и что уже сделано и делается в мире фотографии. Для этого надо смотреть работы других фотографов. Копировать в этом жанре нереально, нужно учиться, а также знать, что уже было сделано другими фотографами. Пробовать снять что-то новое и лучше. В фотографии диких животных, как в прочем и везде, постоянно поднимается планка, и чтобы быть конкурентноспособным, нужно постоянно самосовершенствоваться, устанавливая для себя все новые и новые стандарты. Фотографировать нужно с наслаждением, а навыки приобретаются во время съемки. Снимать нужно много, с наслаждением и не откладывая на завтра. Иначе потом приходится жалеть и кусать локти. Снимая, ошибайтесь, анализируйте и снова снимайте. Чем больше фотографируешь и анализируешь, тем удачливее получаются картинки.
Чем больше фотографируешь и анализируешь, тем удачливее получаются картинки.
5. Если вам повезло и вы встретили животное, которое готово вам позировать, не упускайте момент, работайте с ним столько, сколько это возможно. Завтра этого шанса может не быть, переснять, как это показывает практика, невозможно.
6. Предсказать поведение животного является важным навыком. Нужно быстро оценивать происходящую ситуацию, когда на счету каждая секунда, необходимо успеть извлечь из нее максимум. Тогда, когда вы поймете это, получить хороший снимок будет намного проще.
7. Я люблю возвращаться в одни и те же места в разное время года и по несколько раз. «Почему?», – спросите вы. Маловероятно, что в первый визит можно получить лучший кадр. Он может быть хорош, но велик шанс, что он окажется банальным. Когда у меня уже есть стандартные кадры, можно копнуть глубже и создать оригинальную фотографию. И поэтому я возвращаюсь в одни и те же места по много раз.
И поэтому я возвращаюсь в одни и те же места по много раз.
8. Не жалейте свои фотографии, будьте беспощадны. Просматривая свои снимки, оценивайте что «работает», а что нет. И если нет, то в корзину. Не нужно плодить мусор. Лучше показать миру одну стоящую работу, чем пятьдесят второстепенных.
9. Невозможно овладеть азами съемок живой природы, сидя на диване, дикие животные не живут в студии. Чтобы понять диких животных и сделать хороший снимок, нужно находиться рядом с ними. Нужно раствориться и быть незамеченными, стараться быть частью дикой природы. Животных нужно снимать там, где они живут, растут и погибают, поэтому стоит отправиться туда, где водятся дикие животные, которые вас интересуют. Например, на Камчатку, крайний Север или в Африку. Большинство событий из жизни дикой природы происходит в отдаленных и труднодоступных местах. Поэтому следует быть готовым к дискомфорту, вполне вероятно, что придется забраться глубоко в горы, тундру, густую непроходимую тайгу и ночевать под открытым небом.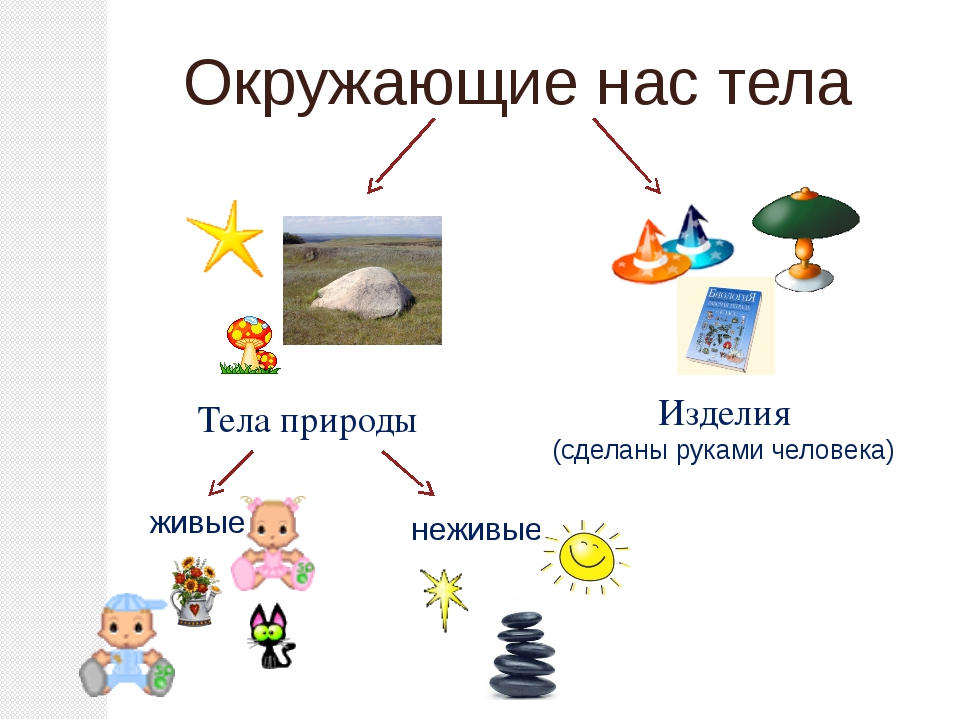 Изучайте повадки и поведение животных – это залог вашей безопасности.
Изучайте повадки и поведение животных – это залог вашей безопасности.
10. Сейчас я выработал для себя главное правило: не нужно гоняться за популярностью, нужно снимать в свое удовольствие. Если есть, что смотреть, зрители это увидят!
Роль природы в жизни человека
«Роль природы в жизни человека».Природа — это все то, что окружает человека и сделано не его руками, а именно атмосфера, ландшафт, деревья, животные, вода и т.д.
Природа — мать человека, она его создала и она влияет на его жизнь самым непосредственным образом. Так , природа дает здоровье человеку, воздух, которым он дышит, землю по которой он ходит, полезные ископаемые и еду.
Природа сделала человека биосоциальным существом, что означает наличие у человека как биологических потребностей, так и социальных. К биологическим (природным) потребностям относят нужду в еде, питье, безопасности, воспроизводстве рода. А к социальным (приобретенным) относят потребность в знаниях, общении, престиже, самореализации и т.д.
А к социальным (приобретенным) относят потребность в знаниях, общении, престиже, самореализации и т.д.
Еще с древних времен люди выбирали место своего жительства исходя из природных условий и окружающего их ландшафта. Так, древние славяне селились вдоль русла рек, чтобы был постоянный источник воды и добычи рыбы. Современный мир трудно представить без борьбы за полезные ископаемые, что также свидетельствует о зависимости человека от природы. Однако, стоит отметить что не только человек зависит от природы. Взаимоотношения человека и природы основаны на взаимосвязи и взаимозависимости. Эта зависимость может проявляться в следующих связях: природа может влиять на общество (как позитивно, так и негативно), общество может влиять на природу (как конструктивно, так и неконструктивно).
Природа дает обществу высокий урожай — это пример позитивного влияния природы на человека. Стихийное бедствие в виде урагана, наводнения, извержения вулкана, землетрясения — негативное влияние природы на общество.
Человек засаживает пустырь деревьями, охраняет редкие виды животных и растений — пример конструктивного влияния человека на природу. Общество загрязняет окружающую среду, убивает животных, вырубает леса и т.д. — некоструктивное взаимодействие.
Таким образом, в современном мире, стали больше внимания уделять глобальным проблемам экологии, так как невооруженным взглядом видно, что если человечество не пересмотрит свое потребительское отношение к природе, то от природы скоро ничего не останется, человек уничтожит свой дом. Поэтому сейчас набирают популярность такие общественные организации и движения как «Гринписс», «Дружины в области охраны природы», «Всемирный фонд дикой природы», Программа ООН «ЮНЕП» и др.
Ответы на вопросы самостоятельной работы в учебниках можно посмотреть В РАЗДЕЛЕ ОБЩЕСТВОЗНАНИЕ (5-11 КЛАСС) или перейти по ссылке.
Различий между натуральными и искусственными материалами
Природные материалы фундаментально отличаются от материалов, созданных руками человека — первые получены из природы, а вторые — из научной лаборатории. Различные типы материалов имеют разное применение, будь то повседневное или специализированное. Вы ежедневно сталкиваетесь со всеми видами природных и искусственных материалов, даже когда идете по улице.
Различные типы материалов имеют разное применение, будь то повседневное или специализированное. Вы ежедневно сталкиваетесь со всеми видами природных и искусственных материалов, даже когда идете по улице.
Происхождение
Хотя все материалы получены из природы, на определенном этапе их производства природные материалы подвергаются меньшей обработке и переработке, чем материалы, созданные руками человека.Натуральные материалы поступают непосредственно из природы — хлопок собирают с хлопковых растений, кукурузу собирают с кукурузных полей, а гранит добывают в карьерах. Искусственные материалы, с другой стороны, проходят тщательную обработку, чтобы изменить материал так, чтобы он соответствовал своему прямому назначению. Обычные искусственные материалы включают пластмассы, которые используются во всем, от напитков в бутылках до одежды и строительства.
Долговечность
Искусственные материалы обычно намного более долговечны, чем их природные аналоги. Фактически, долговечность искусственных материалов, таких как пластмассы, является центральным элементом экологического движения за сокращение потребления и отходов, потому что искусственные материалы накапливаются на свалках, быстро приближаясь к пределу своей емкости, когда они не перерабатываются. Однако природные материалы имеют более короткий срок службы, потому что они когда-то были живыми и постепенно со временем погибают. Деревянная мебель, если не обрабатывать лаком и морилкой, будет гнить, поскольку влага просачивается в ее рамы, а на одежде из натуральных материалов появляются дыры и выцветание.
Фактически, долговечность искусственных материалов, таких как пластмассы, является центральным элементом экологического движения за сокращение потребления и отходов, потому что искусственные материалы накапливаются на свалках, быстро приближаясь к пределу своей емкости, когда они не перерабатываются. Однако природные материалы имеют более короткий срок службы, потому что они когда-то были живыми и постепенно со временем погибают. Деревянная мебель, если не обрабатывать лаком и морилкой, будет гнить, поскольку влага просачивается в ее рамы, а на одежде из натуральных материалов появляются дыры и выцветание.
Уход и обслуживание
Уход за искусственными материалами требует меньше ухода и внимания, чем натуральные материалы. Искусственные материалы легко доступны, потому что они дешевы, долговечны и выносливы — с ними можно обращаться в более грубых условиях, чем с натуральными материалами, и они устойчивы к поломке и повреждению. Например, стирка одежды из полиэстера требует меньшего внимания к усадке, чем стирка одежды из хлопка, потому что ткань специально сделана так, чтобы быть удобной для пользователя. Натуральные материалы могут быть повреждены, если для дезинфекции или обработки их поверхностей использовать синтетическое мыло, красители или другие чистящие средства.
Натуральные материалы могут быть повреждены, если для дезинфекции или обработки их поверхностей использовать синтетическое мыло, красители или другие чистящие средства.
Воздействие на окружающую среду
Искусственные материалы не только способствуют постоянно растущим свалкам во всем мире, но и могут оказывать негативное воздействие на окружающую среду, поскольку они не являются экологически безопасными. Агентство по охране окружающей среды (EPA) определяет устойчивость как «политику и стратегии, которые удовлетворяют нынешние потребности общества без ущерба для способности будущих поколений удовлетворять свои собственные потребности.«Искусственные продукты на нефтяной основе, включая семейство пластмасс, не классифицируются как экологически безопасные, поскольку они зависят от добычи и переработки нефти, ограниченного природного ресурса.
Все больше и больше коммерческих компаний обращаются к экологически чистым материалам, таким как бамбук, чтобы привлечь внимание общественности к проблемам окружающей среды и уменьшить воздействие определенных потребительских товаров на землю. Бамбук — это эластичный природный материал, который можно легко обрабатывать и который быстро растет, не повреждая землю и не потребляя слишком много природных ресурсов. .
Бамбук — это эластичный природный материал, который можно легко обрабатывать и который быстро растет, не повреждая землю и не потребляя слишком много природных ресурсов. .
Природа, созданная людьми — Новая концепция природы • TEDxVienna
Природа исчезает! Большинство из вас согласятся с последним предложением. В наших городах меньше зелени, нет нетронутых ландшафтов или тихих мест из-за шума транспорта. У природы часто бывает этот романтизированный образ : она спокойная, гармоничная, красивая и полезная для нас . Природный бренд «Это хорошо для вас» также используется для продажи продуктов. Такие этикетки, как « 100% натуральный » (может ли что-то быть менее 100% натуральным?) Создают образ предположительно здорового, безопасного и экологически чистого продукта.Немного отстраняясь, довольно быстро понимаешь, что природа, напротив, также может быть непредсказуемой, вредной и уродливой. Подумайте, например, о наводнениях, землетрясениях и тараканах. Это действительно не очень приятно, но, похоже, люди сильно забывают об этом, когда думают о щедрой матери-природе.
Это действительно не очень приятно, но, похоже, люди сильно забывают об этом, когда думают о щедрой матери-природе.
Однако перед лицом технологических достижений с почти неограниченной скоростью, технофобия растет. Люди не придумывают приятных описаний технологии, а скорее ассоциируют «искусственное» как вредное и плохое.
Самый большой парадокс состоит в том, что природа улучшается, делая ее красивее и привлекательнее для нас. Так сказать, сделать природу более естественной. Только желтый, блестящий банан без косточек имеет вкусный вкус. Светящиеся рыбки обновляют обычные аквариумы в жилых комнатах людей. Эта « гиперприрода » на самом деле в высшей степени искусственна.
Что такое природа?
Природа и наши представления о природе изучаются с момента возникновения цивилизации.Объекты и события подразделяются на « рожденных, » и, следовательно, «естественные» или «вызванные или созданные человеком, », что связано с термином «искусственные или ненатуральные». Таким образом, культура, включая людей и создаваемые ими сущности, находится вне природы, хотя люди рождаются из природы. Эта сильная грань между двумя противоположными категориями теперь исчезает в нашем высокотехнологичном обществе. Например, синтетическая биология позволяет нам создавать клетки, управляемые синтетическими геномами, которые воспроизводятся сами по себе.Но тогда, считаем ли мы естественными новорожденные клетки искусственной клетки?
Таким образом, культура, включая людей и создаваемые ими сущности, находится вне природы, хотя люди рождаются из природы. Эта сильная грань между двумя противоположными категориями теперь исчезает в нашем высокотехнологичном обществе. Например, синтетическая биология позволяет нам создавать клетки, управляемые синтетическими геномами, которые воспроизводятся сами по себе.Но тогда, считаем ли мы естественными новорожденные клетки искусственной клетки?
Природа создана людьми
Когда вы смотрите вокруг на улице, вы можете быть окружены обычными вещами, такими как пробки, Макдональдс, рестораны и т. Д. — мир, созданный людьми . Со временем культура может стать природой или Next Nature , как это называется в философской концепции Кёрта Ван Менсворта и Хендрика-Яна Гривинка. Эта возникшая в культурном отношении природа более знакома нам, чем старая природа, созданная из деревьев, животных и растений.Поскольку технологии становятся настолько распространенными, мы уже воспринимаем их как естественные.
Вездесущее понятие природы, эволюции, также вторгается в культуру. Экономика и технологии действуют как факторы окружающей среды, движущие дарвиновским отбором. Эта негенетическая эволюция может привести к появлению новых видов продуктов, таких как Razorius Gillettus , бритвы с пятью лезвиями, которая произошла от прежней модели с двумя лезвиями в результате такого эволюционного развития, а не просто из-за потребности в лучшем бритье.
Одновременно возделывается традиционная природа . Одним из примеров является приручение диких животных покорным питомцам. В общем, человечество постоянно меняет природу, и, в свою очередь, человек эволюционируют вместе с технологиями, которые они создают. Это изображение природы как высокодинамичного мира , который никогда не останется прежним, в отличие от столь часто оплакиваемой потерянной природы, которую считают статичной.
Next Nature — КОНТРОЛИРУЕМЫЙ против НЕКонтролируемого
Ван Менсвоорт и Гривинк радикально меняют наше представление о природе, предлагая выбросить за борт наш дуализм «рожденное против сделанного» и, скорее, думать о природе как о неподвластной нам. Рожденные и неконтролируемые объекты и процессы составляют старую природу, включая, например, солнце и грозы, тогда как следующая природа создается людьми, но все еще неконтролируема, как пробки на дорогах. Легко управляемые системы, такие как дерево бонсай или противозачаточные средства, выглядят в этом свете как культурная природа и культура, соответственно
Рожденные и неконтролируемые объекты и процессы составляют старую природу, включая, например, солнце и грозы, тогда как следующая природа создается людьми, но все еще неконтролируема, как пробки на дорогах. Легко управляемые системы, такие как дерево бонсай или противозачаточные средства, выглядят в этом свете как культурная природа и культура, соответственно
Посмотрите выступление TED на Next Nature:
Мы живем в ноосфере
Заставляющая задуматься концепция Next Nature создает новый словарный запас, позволяющий смотреть на мир вокруг нас.Наша технология превращает природу в новый вид природы, который так же реален, как и старая природа, поскольку она дикая и непредсказуемая, как всегда. Еще важнее то, что прогресс присущ людям, и поэтому сохранить прошлую природу невозможно. Около пяти миллиардов лет назад Земля была покрыта геосферой, которую сменила биосфера, позволившая зародиться жизни. Владимир Верандский и Тейяр де Шарден предположили в начале 20-го века, что биосфера превратилась в ноосферу (от греческих слов «разум» и «сфера») в результате появления человеческого познания, и описывает коллективное человеческое сознание. Остается только предположить, что будет после ноосферы, но определенно возникнет что-то новое.
Владимир Верандский и Тейяр де Шарден предположили в начале 20-го века, что биосфера превратилась в ноосферу (от греческих слов «разум» и «сфера») в результате появления человеческого познания, и описывает коллективное человеческое сознание. Остается только предположить, что будет после ноосферы, но определенно возникнет что-то новое.
Необходимо отказаться от нашего нынешнего ограниченного взгляда на природу, чтобы развить осознание того, что мы меняем природу, и это неплохо. Напротив, наша технология может направлять развитие природы. Да, мы даже можем использовать Next Nature, чтобы спасти старую природу . Просто подумайте об использовании генной инженерии, чтобы вернуть вымершие виды.
Как только мы осознаем, что мы не можем отступить ни на шаг, но можем сформировать то, что будет, мы можем начать думать о том, как мы хотим это сделать.
Заголовочные изображения с сайта Pixabay & Gratisography
Урок в детском саду Естественное и рукотворное
В этой части урока я использую свою доску SMART Board. Если у вас есть SMART Board, файл Natural vs. Man-Made Notebook File можно легко загрузить и открыть. Если у вас есть интерактивная доска другого типа, вы все равно можете использовать этот урок, открыв файл в Smart Notebook Express. Также имеется PDF-файл слайдов, чтобы вы могли воссоздать эту часть урока: Естественный и искусственный PDF-файл слайдов
Если у вас есть SMART Board, файл Natural vs. Man-Made Notebook File можно легко загрузить и открыть. Если у вас есть интерактивная доска другого типа, вы все равно можете использовать этот урок, открыв файл в Smart Notebook Express. Также имеется PDF-файл слайдов, чтобы вы могли воссоздать эту часть урока: Естественный и искусственный PDF-файл слайдов
Я собираю своих учеников перед SMART Board.У меня есть карточки с именами каждого ученика. Эти карточки используются для выбора тех, кто пойдет на SMART Board.
Я открываю первый слайд (слайд Smartboard Slide 1) с целью урока, написанной «дружественными для учащихся» терминами. Есть цель содержания и цель языка, чтобы помочь сосредоточиться на расширении словарного запаса для моих изучающих английский язык (EL), чтобы соответствовать методам обучения SIOP. Я зачитываю эти цели своим ученикам.
Цель содержания
Я могу отличить что-то от природы от того, что создано руками человека.
Языковая цель
Я могу сказать другу, создано ли что-то природное или рукотворное.
Затем я перехожу к Slide Two .
Слайд 2: Если что-то от природы, это вообще не было изменено людьми. Эти вещи от природы. Вы знаете, что это такое? Мы просматриваем слайд, и студенты рассказывают мне, что представляет собой каждый предмет. Я прошу учеников привести мне другие примеры вещей, происходящих от природы.
Слайд третий: Если что-то создано руками человека, то это было создано людьми. Вы знаете, что это за предметы? Мы снова просматриваем слайд, предлагая студентам назвать каждый элемент, а я предлагаю студентам привести мне примеры других вещей, созданных руками человека.
Слайд четвертый: Мои ученики знакомы с «Повернись и говори». Мы используем Turn and Talk, чтобы помочь развить навыки устной речи, особенно у моих учеников EL.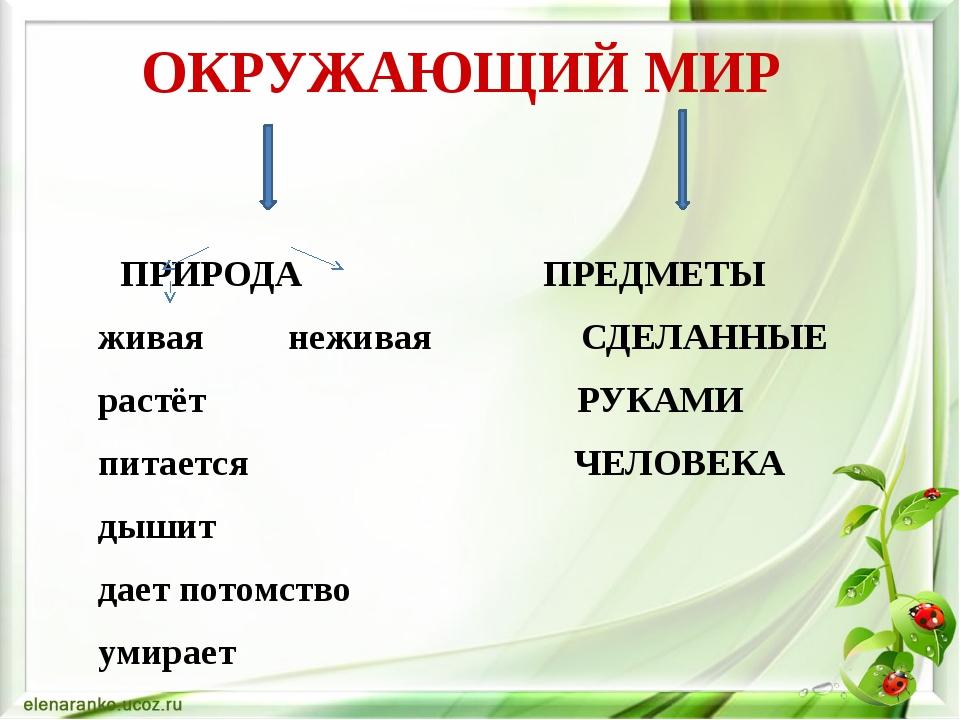 У моих учеников есть назначенный партнер «Повернись и поговори».Затем я говорю студентам: А теперь повернитесь к своему партнеру «Повернись и поговори» и ответь на этот вопрос: этот предмет исходит от природы или создан руками человека?
У моих учеников есть назначенный партнер «Повернись и поговори».Затем я говорю студентам: А теперь повернитесь к своему партнеру «Повернись и поговори» и ответь на этот вопрос: этот предмет исходит от природы или создан руками человека?
После того, как у студентов была возможность поговорить, я прошу добровольца поделиться тем, что придумала их группа. Мы говорим о том, что, несмотря на то, что орел создан природой, кто-то сделал эту статую, поэтому она сделана руками человека.
Затем я прошу студентов вернуться к своим столам, потому что мы собираемся вернуться к сортировке карточек с начала урока.
Границы | Слуховая категоризация искусственных звуков по сравнению с естественными звуками с помощью функциональной связи мозга MEG
Введение
Текущие знания о нейронных сетях, лежащих в основе слухового восприятия, остаются фрагментарными, несмотря на тот факт, что слух широко изучен (Zatorre et al., 2002). Основные сетевые свойства, которые были предложены для слуховой модальности, напоминают структуру визуальной, разделяя пути обработки информации в дорсальной и вентральной сетях, соответствующие обработке информации на «где» и «что», соответственно (Romanski et al.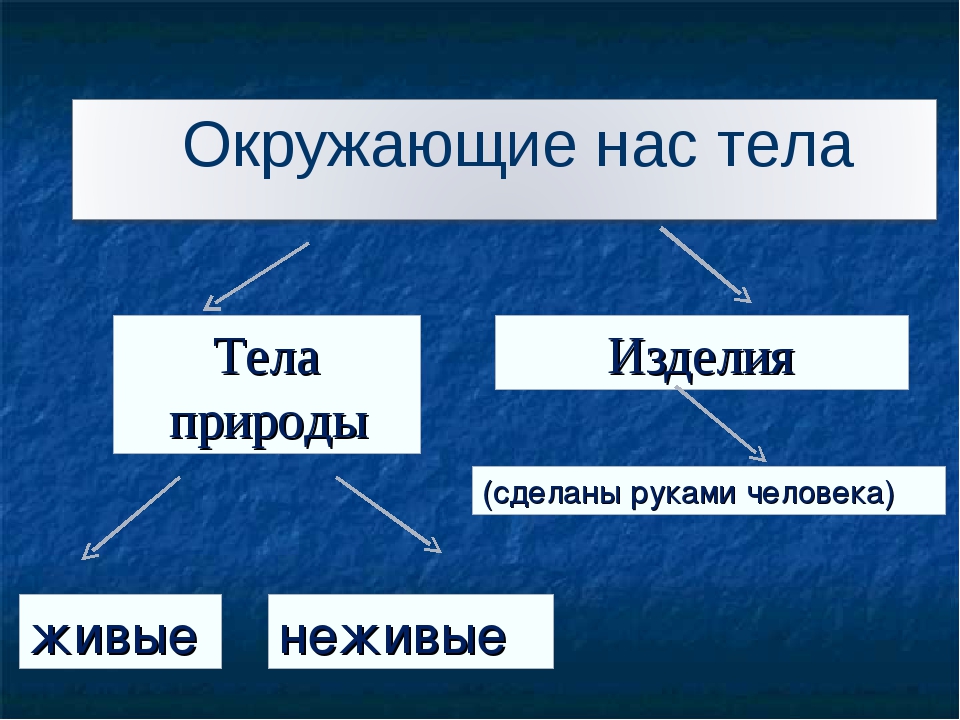 al., 2000; Кубовы и Ван Валкенбург, 2001; Арнотт и др., 2004; Huddleston et al., 2008; Asplund et al., 2010). Следуя пути «что», физические характеристики звукового стимула изначально кодируются в первичной и вторичной слуховой коре, а также в их ассоциативных областях, прежде чем они будут интегрированы в более абстрактное представление (Griffiths and Warren, 2004; Bregman, 2017). ). В рамках этого пути обработка слуховой информации, по-видимому, осуществляется в каналах, специфичных для звуковой категории (Caramazza and Mahon, 2003).В этой строке высказывались предположения о функциональной специфичности обработки различных типов звуков (Belin et al., 2000; Zatorre and Belin, 2001; Patterson et al., 2002; Lewis et al., 2004; Zatorre et al. , 2004; Hunter et al., 2010).
al., 2000; Кубовы и Ван Валкенбург, 2001; Арнотт и др., 2004; Huddleston et al., 2008; Asplund et al., 2010). Следуя пути «что», физические характеристики звукового стимула изначально кодируются в первичной и вторичной слуховой коре, а также в их ассоциативных областях, прежде чем они будут интегрированы в более абстрактное представление (Griffiths and Warren, 2004; Bregman, 2017). ). В рамках этого пути обработка слуховой информации, по-видимому, осуществляется в каналах, специфичных для звуковой категории (Caramazza and Mahon, 2003).В этой строке высказывались предположения о функциональной специфичности обработки различных типов звуков (Belin et al., 2000; Zatorre and Belin, 2001; Patterson et al., 2002; Lewis et al., 2004; Zatorre et al. , 2004; Hunter et al., 2010).
Попытки разделить обработку различных категорий звуков на корковом уровне были сделаны в тематических исследованиях поражений головного мозга (Clarke et al., 2000; Taniwaki et al., 2000; Mendez, 2001; Steinke et al. , 2001). Случаи, подобные слуховой агнозии, представляют собой нарушение способности распознавать звуки при сохранении периферического слуха.Однако это ухудшение не обязательно распространяется на все категории звука; это скорее может быть категориально в зависимости от повреждения мозга. Например, пациент с очаговым повреждением в правой лобно-теменной области смог идентифицировать звуки окружающей среды и назвать музыкальные инструменты, но не смог распознать музыку (Steinke et al., 2001). С другой стороны, левостороннее височное поражение или лево-лобно-височная ишемия вызывают агнозию, ограниченную звуками окружающей среды (Clarke et al., 2000). Однако следует подчеркнуть, что исследования отдельных случаев пациентов с поражением головного мозга очень разнородны и поэтому не могут предоставить подробную модель обработки звука коры головного мозга.
, 2001). Случаи, подобные слуховой агнозии, представляют собой нарушение способности распознавать звуки при сохранении периферического слуха.Однако это ухудшение не обязательно распространяется на все категории звука; это скорее может быть категориально в зависимости от повреждения мозга. Например, пациент с очаговым повреждением в правой лобно-теменной области смог идентифицировать звуки окружающей среды и назвать музыкальные инструменты, но не смог распознать музыку (Steinke et al., 2001). С другой стороны, левостороннее височное поражение или лево-лобно-височная ишемия вызывают агнозию, ограниченную звуками окружающей среды (Clarke et al., 2000). Однако следует подчеркнуть, что исследования отдельных случаев пациентов с поражением головного мозга очень разнородны и поэтому не могут предоставить подробную модель обработки звука коры головного мозга.
Несколько функциональных нейровизуализационных исследований со здоровыми участниками указывают на двустороннюю активацию слуховой коры для речевых звуков (Belin et al. , 2000; Zatorre and Belin, 2001) и правостороннюю активацию для неречевых звуков во время задач распознавания звука (для обзора см. Tervaniemi и Hugdahl, 2003). Сообщалось также о вовлечении других областей мозга, таких как нижняя лобная кора, что указывает на сеть, которая включает в себя другие когнитивные функции, помимо слуховых (см. Обзор в Price, 2012).До сих пор большинство текущих исследований в основном сосредоточено на дифференциальной обработке речи по сравнению с категориями неречевых звуков, хотя мы все еще плохо знаем о дифференциальной обработке в категории неречевых звуков. Существующие исследования показали, что дискриминация звуковых категорий с большей вероятностью зависит от связанных с ней манипулятивных характеристик (Lewis et al., 2004, 2005; Murray et al., 2006; De Lucia et al., 2009, 2012). Например, в контексте игры на гитаре мы слушаем звук, когда воспринимаем двигательные и визуальные действия игры на гитаре.Таким образом, на более высоком когнитивном уровне информация от всех сенсорных модальностей, которые получают входные данные от стимула, интегрируется, чтобы построить восприятие звука.
, 2000; Zatorre and Belin, 2001) и правостороннюю активацию для неречевых звуков во время задач распознавания звука (для обзора см. Tervaniemi и Hugdahl, 2003). Сообщалось также о вовлечении других областей мозга, таких как нижняя лобная кора, что указывает на сеть, которая включает в себя другие когнитивные функции, помимо слуховых (см. Обзор в Price, 2012).До сих пор большинство текущих исследований в основном сосредоточено на дифференциальной обработке речи по сравнению с категориями неречевых звуков, хотя мы все еще плохо знаем о дифференциальной обработке в категории неречевых звуков. Существующие исследования показали, что дискриминация звуковых категорий с большей вероятностью зависит от связанных с ней манипулятивных характеристик (Lewis et al., 2004, 2005; Murray et al., 2006; De Lucia et al., 2009, 2012). Например, в контексте игры на гитаре мы слушаем звук, когда воспринимаем двигательные и визуальные действия игры на гитаре.Таким образом, на более высоком когнитивном уровне информация от всех сенсорных модальностей, которые получают входные данные от стимула, интегрируется, чтобы построить восприятие звука. Подобная модель (Heekeren et al., 2008) была предложена в прошлом для зрительной и соматосенсорной системы, указывая на мультисенсорную интеграцию уже на ранних стадиях когнитивной обработки на низком уровне.
Подобная модель (Heekeren et al., 2008) была предложена в прошлом для зрительной и соматосенсорной системы, указывая на мультисенсорную интеграцию уже на ранних стадиях когнитивной обработки на низком уровне.
Данные о функциональной организации слухового восприятия показывают, что звуки можно разделить на «живые» и «искусственные» стимулы (Lewis et al., 2005), предполагая дифференциальную активацию мозга. В частности, искусственный объект по сравнению со звуком животного может потребовать нисходящего механизма, который объединяет семантические и мультисенсорные особенности, связанные в большей степени с действием. Аналогичным образом Мюррей и др. (2006) продемонстрировали с помощью ЭЭГ, что «искусственные» звуки демонстрируют более сильную активацию мозга в слуховом пути «что?» По сравнению с «живыми» объектами, и что в основном задействованы области правого полушария и премоторная кора.Сообщалось также о других различиях в категории искусственных объектов. Основная идея заключается в том, что звуки повседневных предметов, такие как звонок телефона, могут вызывать большую реакцию на действие, чем типичный тон музыкального инструмента (у не музыкантов), и, таким образом, будет вызвана более сильная активация мозга (Де Люсия и др. ., 2009). Интересно, что исследования ЭЭГ, сфокусированные на временной динамике, показали, что процесс различения категорий происходит вокруг компонента N1, уже через 70 мс после появления стимула (Murray et al., 2006; Де Люсия и др., 2012).
., 2009). Интересно, что исследования ЭЭГ, сфокусированные на временной динамике, показали, что процесс различения категорий происходит вокруг компонента N1, уже через 70 мс после появления стимула (Murray et al., 2006; Де Люсия и др., 2012).
Тем не менее, то, как отдельные сети работают с разными категориями звуков, все еще плохо изучено. Хотя вышеупомянутые исследования дали некоторое представление о том, когда и где появляется эта дифференциация, анализ исходного пространства предлагает простые карты активированных областей мозга, а не показывает, как эти области распределенной системы функционально связаны для выполнения конкретной задачи. На сегодняшний день исследование сложных сетей было разработано методологически, что дало возможность изучить кортикальную реорганизацию, лежащую в основе связанных когнитивных процессов (Rogers et al., 2007; Bullmore and Sporns, 2009). Таким образом, в текущем исследовании мы стремились дополнительно изучить функциональную связность пути слуховой обработки в различных категориях неречевых звуков. Сравнивались корковые ответы трех разных категорий звуков. А именно, категория Musical и категория Artificial (звуки повседневно используемых / слышимых объектов), представляющие звуковые категории искусственных объектов и категорию Natural (в основном вокализации животных).Насколько нам известно, это первое исследование, в котором изучается функциональная связь между различными категориями неречевых звуков с помощью магнитоэнцефалографии (МЭГ), которая имеет высокое пространственное разрешение и отличное временное разрешение. Принимая во внимание существующую литературу о живых и созданных человеком звуках, мы ожидали, что звуки Musical и Artificial будут демонстрировать более сильные корковые реакции, которые будут включать области, связанные с моторикой, и значительные взаимосвязи между этими областями по сравнению с Natural звуков.Кроме того, следует отметить, что хотя звуки Musical и Artificial относятся к категории искусственных, ранее сообщалось о доказательствах дифференциальной активации между этими группами (De Lucia et al.
Сравнивались корковые ответы трех разных категорий звуков. А именно, категория Musical и категория Artificial (звуки повседневно используемых / слышимых объектов), представляющие звуковые категории искусственных объектов и категорию Natural (в основном вокализации животных).Насколько нам известно, это первое исследование, в котором изучается функциональная связь между различными категориями неречевых звуков с помощью магнитоэнцефалографии (МЭГ), которая имеет высокое пространственное разрешение и отличное временное разрешение. Принимая во внимание существующую литературу о живых и созданных человеком звуках, мы ожидали, что звуки Musical и Artificial будут демонстрировать более сильные корковые реакции, которые будут включать области, связанные с моторикой, и значительные взаимосвязи между этими областями по сравнению с Natural звуков.Кроме того, следует отметить, что хотя звуки Musical и Artificial относятся к категории искусственных, ранее сообщалось о доказательствах дифференциальной активации между этими группами (De Lucia et al. , 2009) в зависимости от ежедневное использование. Слуховое вызванное поле N1 было априори , установленным как интересующий временной интервал на основании предыдущих электрофизиологических данных, которые сообщают о ранних ответах при обработке различения звуковых категорий (Murray et al., 2006; Де Люсия и др., 2009, 2012).
, 2009) в зависимости от ежедневное использование. Слуховое вызванное поле N1 было априори , установленным как интересующий временной интервал на основании предыдущих электрофизиологических данных, которые сообщают о ранних ответах при обработке различения звуковых категорий (Murray et al., 2006; Де Люсия и др., 2009, 2012).
Материалы и методы
Субъекты
Настоящее исследование было проведено с выборкой из 20 молодых людей (средний возраст = 27,19, SD = 5,59, 8 мужчин). В соответствии с клинической аудиометрической оценкой, они были отобраны из пула субъектов нашего института среди лиц с нормальным слухом. Все испытуемые были правшами, согласно Эдинбургской инвентаризации рук (Oldfield, 1971). Участники были проинформированы о цели исследования, а желающим принять участие была предоставлена форма согласия, которая гарантировала конфиденциальность их личности.Исследование было проведено в соответствии с Хельсинкской декларацией и одобрено этическим комитетом медицинского факультета Мюнстерского университета.
Стимулы
Стимулы состояли из трех различных категорий звуков: Естественные , Музыкальные и Искусственные . Звуки Natural и Artificial были записями, полученными из онлайн-баз данных звуков (Free Sounds Effects; SoundBible; ZapSplat). Музыкальные звука были получены из звукового банка «Мастер сэмплов Университета Макгилла», который был создан для исследования восприятия, связанного с психологией музыки.Программное обеспечение Audacity использовалось для повторной дискретизации всех звуков с частотой 44 100 Гц и для реализации линейных наклонов начала / смещения в 20 мс. Затем монофонические звуки были преобразованы в стереозвук. С помощью редактора звука WavePad они были нормализованы на –10 дБ RMS на основе метода нормализации средней громкости.
Парадигма стимулов была выполнена с помощью программного обеспечения Presentation (версия 18.0, Neurobehavioral systems, Inc., Беркли, Калифорния, США). Он состоял из двух блоков с небольшим перерывом между ними. Каждый блок включал в себя представление трех различных категорий звуков, которые были псевдослучайно представлены по блокам и по предметам, тогда как звуки каждой категории всегда были представлены в одном и том же порядке: A = Искусственный , M = Музыкальный и N = натуральный ; Блок 1: A1-A2-… -An-M1-M2-… -Mn-N1-N2-… -Nn; Блок 2: M1-M2-… -Mn-N1-N2-… -Nn-A1-A2-… -An. Каждый блок содержал 144 стимула, по 48 на каждую категорию, что в сумме составляет 288 стимулов для всего эксперимента; 48 (звуков на категорию) × 3 (категории) × 2 (блоки).Каждый блок содержал 144 стимула, по 48 на каждую категорию, что в сумме составляет 288 стимулов для всего эксперимента; 48 (звуков на категорию) × 3 (категории) × 2 (блоки). Каждый стимул длился 1 с с рандомизированным интервалом между стимулами (ISI) от 0,7 до 1,3 с, чтобы избежать ожидания и ритмичности. Natural звуки содержат звуки живых объектов. Музыкальные звуки содержали ноты различных музыкальных инструментов, тогда как Искусственные звуки были повседневными предметными звуками.
Каждый блок включал в себя представление трех различных категорий звуков, которые были псевдослучайно представлены по блокам и по предметам, тогда как звуки каждой категории всегда были представлены в одном и том же порядке: A = Искусственный , M = Музыкальный и N = натуральный ; Блок 1: A1-A2-… -An-M1-M2-… -Mn-N1-N2-… -Nn; Блок 2: M1-M2-… -Mn-N1-N2-… -Nn-A1-A2-… -An. Каждый блок содержал 144 стимула, по 48 на каждую категорию, что в сумме составляет 288 стимулов для всего эксперимента; 48 (звуков на категорию) × 3 (категории) × 2 (блоки).Каждый блок содержал 144 стимула, по 48 на каждую категорию, что в сумме составляет 288 стимулов для всего эксперимента; 48 (звуков на категорию) × 3 (категории) × 2 (блоки). Каждый стимул длился 1 с с рандомизированным интервалом между стимулами (ISI) от 0,7 до 1,3 с, чтобы избежать ожидания и ритмичности. Natural звуки содержат звуки живых объектов. Музыкальные звуки содержали ноты различных музыкальных инструментов, тогда как Искусственные звуки были повседневными предметными звуками. Примеры звуковых файлов, использованных в исследовании, можно найти в Дополнительных материалах.
Примеры звуковых файлов, использованных в исследовании, можно найти в Дополнительных материалах.
MEG записей
Участников обследовали в магнитно-экранированной и акустически тихой комнате с помощью 275-канальной системы для всей головы (OMEGA 275, CTF, VSM Medtech Ltd., Ванкувер, Британская Колумбия, Канада). Данные непрерывно записывались с частотой дискретизации 600 Гц, что приводило к частоте отсечки в автономном режиме 150 Гц. Участники сидели в вертикальном положении, а их голова была стабилизирована с помощью ватных дисков внутри шлема MEG.Немой фильм демонстрировался на экране проектора, установленном на гентри системы MEG, размещенном в соответствии с наилучшим углом обзора участников, чтобы они не теряли бдительности во время эксперимента; как применялось в предыдущих слуховых экспериментах (Pantev et al., 2004; Okamoto et al., 2008; Paraskevopoulos et al., 2018). После прохождения электростатических преобразователей слуховые стимулы доставлялись через силиконовые трубки длиной 60 см и внутренним диаметром 5 мм, заканчивающиеся силиконовым наушником, индивидуально подогнанным к уху каждого испытуемого. Перед стимуляцией был проведен аудиологический тест на определение порога слуха с точностью 5 дБ на частоте 1 кГн. Уровни звукового давления стимула были установлены на 60 дБ SL выше индивидуального порога слышимости. Весь эксперимент длился около 30 мин.
Перед стимуляцией был проведен аудиологический тест на определение порога слуха с точностью 5 дБ на частоте 1 кГн. Уровни звукового давления стимула были установлены на 60 дБ SL выше индивидуального порога слышимости. Весь эксперимент длился около 30 мин.
Протокол МРТ
МРТ-изображение, взвешенное по Т1, было выполнено для всех участников на сканере 3 Тесла (Gyroscan Intera T30, Philips), чтобы получить модель конечных элементов (МКЭ) головы индивидуума.В файлах представлены изображения 400 однослойных срезов толщиной 0,5 мм в сагиттальной плоскости (TR = 7,33,64 мс, TE = 3,31 мс). Размер матрицы каждого среза составлял 512 × 512 с размером вокселя 0,5 × 0,58 × 0,58 мм 3 . Чтобы обеспечить надежность исследования структуры мозга внутри и между субъектами, мы использовали SPM12 (статистическое параметрическое отображение) для регулирования неоднородности интенсивности (Ganzetti et al., 2016), и поэтому изображения были преобразованы в изотропные воксели 2 × 2 × 2 мм.
Анализ данных MEG
Анализ данных MEG проводился в соответствии с ранее разработанным анализом, применяемым для функциональных сетей связи в различных слуховых парадигмах (Paraskevopoulos et al., 2015, 2018). Программное обеспечение для анализа электрического источника мозга (BESA MRI, версия 2.0, Megis Software, Гейдельберг, Германия) использовалось для расчета модели головы человека путем сегментации четырех различных тканей головы (скальп, череп, спинномозговая жидкость и мозг) на основе МКЭ. Четырехслойная модель FEM дает более точные результаты по сравнению с другими моделями, так как она включает CSF (Ramon et al., 2004; Wendel et al., 2008), который является высокопроводящим слоем и важен для реконструкции источника MEG (Wolters et al., 2006). Датчики МЭГ были совместно зарегистрированы и настроены на структурную МРТ человека с помощью назиона, а также левого и правого входов в ушные каналы в качестве ориентиров. С помощью интерполяции 3D-сплайном МРТ были преобразованы в ACPC (передне-заднюю поясную извилину) и в пространство Talairach. Предварительно определенный вариант значений проводимости (см. Wolters et al., 2006) был установлен для кожного отсека на 0.33 См / м, для черепа до 0,0042 См / м, для спинномозговой жидкости до 0,79 См / м и для ткани мозга до 0,33 См / м.
Предварительно определенный вариант значений проводимости (см. Wolters et al., 2006) был установлен для кожного отсека на 0.33 См / м, для черепа до 0,0042 См / м, для спинномозговой жидкости до 0,79 См / м и для ткани мозга до 0,33 См / м.
Для предварительной обработки данных МЭГ использовалось исследовательское программное обеспечение BESA (версия 6.0, Megis Software, Гейдельберг, Германия). Для отклонения артефактов использовалась автоматическая электрокардиограмма (ЭКГ) и обнаружение и коррекция артефактов моргания глаз, предоставленные BESA (Ille et al., 2002). Данные были отфильтрованы в автономном режиме с помощью режекторного фильтра 50 Гц, фильтра нижних частот с нулевой фазой 45 Гц и фильтра верхних частот с нулевой фазой 0.5 Гц. Данные были разделены на эпохи 1000 мс после начала и 500 мс до начала стимула. Была применена коррекция базовой линии на основе интервала до стимула 100 мс. Во время усреднения эпох, связанных со стимулом, испытания с амплитудами более 3 pT и данными, превышающими 15% отклоненных испытаний, были исключены из анализа. Затем два блока измерения были усреднены для каждого участника, чтобы улучшить отношение сигнал / шум.
Затем два блока измерения были усреднены для каждого участника, чтобы улучшить отношение сигнал / шум.
Для реконструкции плотности тока мы использовали временное окно вокруг главного компонента N1 медленного слухового вызванного поля (Pantev et al., 1993; c.f. Рисунок 2), который в соответствии с глобальной мощностью поля данных общего среднего значения был между 80 мс и 120 мс после начала стимула, включая возрастающий наклон N1. Электромагнитная томография низкого разрешения (LORETA), предоставленная BESA, была применена для реконструкции источника для каждого объекта и каждой категории звуков, поскольку она обеспечивает плавное распределение источников в качестве обратного решения (Pascual-Marqui et al., 1994). Он основан на методе взвешенной минимальной нормы (Grech et al., 2008) и не опирается на определение a priori активированных источников.
Статистический анализ
Для статистического анализа реконструкции LORETA мы использовали SPM12, работающий на программном обеспечении Matlab (версия R2016b; MathWorks Inc. , Натик, Массачусетс, США). Была установлена явная маска, чтобы включать результаты только для серого вещества, тем самым уменьшая объем поиска. Односторонний анализ ANOVA проводился с тремя различными категориями звуков ( Natural , Artificial и Musical ) в качестве фактора внутри субъектов.Затем были разработаны таблицы-матрицы F- и t-контраста (на основе общей линейной модели) для проверки статистических различий между тремя категориями и между категориями, соответственно. Для управления множественными сравнениями была реализована функция Family Wise Error (FWE).
, Натик, Массачусетс, США). Была установлена явная маска, чтобы включать результаты только для серого вещества, тем самым уменьшая объем поиска. Односторонний анализ ANOVA проводился с тремя различными категориями звуков ( Natural , Artificial и Musical ) в качестве фактора внутри субъектов.Затем были разработаны таблицы-матрицы F- и t-контраста (на основе общей линейной модели) для проверки статистических различий между тремя категориями и между категориями, соответственно. Для управления множественными сравнениями была реализована функция Family Wise Error (FWE).
Анализ связи
Чтобы изучить кортикальную сеть из важных источников, полученных из анализа SPM12, мы дополнительно реализовали анализ связности. Определив активированные области в пространстве источника с помощью описанного выше анализа, мы использовали эквивалентную дипольную модель тока, установив один диполь на пик каждого значимого кластера, полученного с помощью F-контраста.В результате получилось пять эквивалентных токовых диполей. В связи с тем, что SPM выражает координаты на основе стандартизованного мозга Монреальским неврологическим институтом (координаты MNI), координаты были перенесены в пространство Talairach, чтобы соответствовать координатам мозга программного обеспечения BESA, где была запущена дипольная модель. Для преобразования использовался апплет «NMI2TAL» из пакета Yale BioImage Suite (sprout022.sprout.yale.edu), который основан на координатах отображения Lacadie et al. (2008).Ориентация диполей была подобрана на основе индивидуального объемного проводника МКЭ, тогда как координаты были фиксированными для всех субъектов и условий, как определено выше. Результаты содержали пять форм сигналов источника, соответствующих каждому диполю, включая интервал 80–120 мс.
В связи с тем, что SPM выражает координаты на основе стандартизованного мозга Монреальским неврологическим институтом (координаты MNI), координаты были перенесены в пространство Talairach, чтобы соответствовать координатам мозга программного обеспечения BESA, где была запущена дипольная модель. Для преобразования использовался апплет «NMI2TAL» из пакета Yale BioImage Suite (sprout022.sprout.yale.edu), который основан на координатах отображения Lacadie et al. (2008).Ориентация диполей была подобрана на основе индивидуального объемного проводника МКЭ, тогда как координаты были фиксированными для всех субъектов и условий, как определено выше. Результаты содержали пять форм сигналов источника, соответствующих каждому диполю, включая интервал 80–120 мс.
Набор инструментов HERMES (Niso et al., 2013) в Matlab использовался для построения матрицы смежности 5 × 5 для каждого объекта и каждого условия на основе алгоритма взаимной информации (MI), который измеряет взаимозависимость между переменными и определяет корреляции случайных величин с нелинейной мерой зависимости (Zeng, 2015).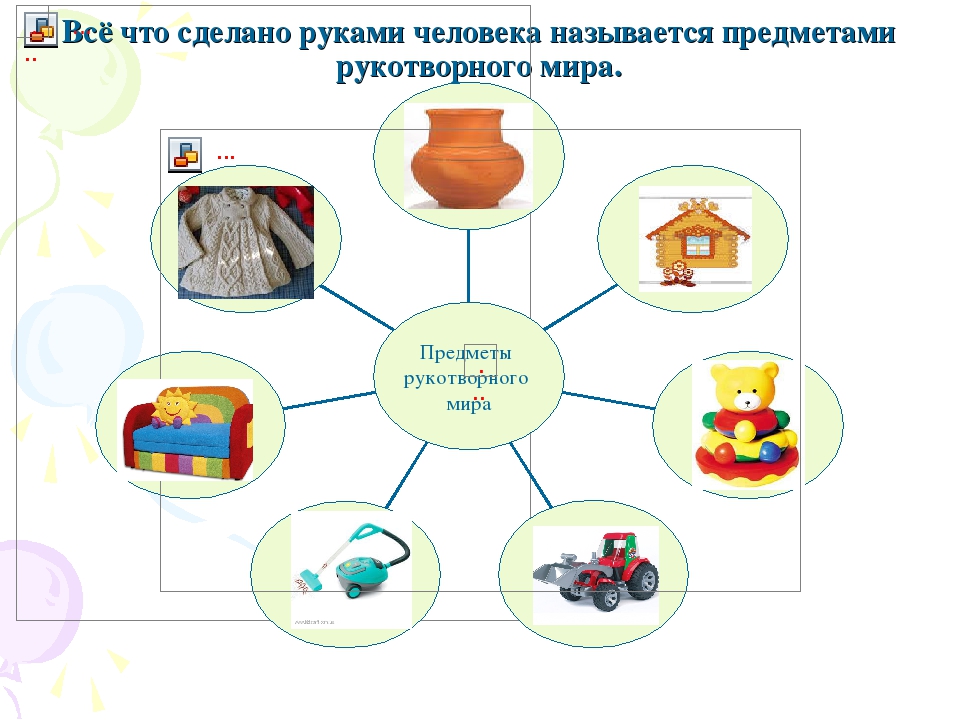 Затем результаты были перенесены в Network Based Statistic Toolbox (NBS; Zalesky et al., 2010) для изучения статистически значимых связей. Односторонний внутрисубъектный дисперсионный анализ ANOVA проводился с тремя условиями в качестве фактора внутри субъектов. Метод NBS был установлен для множественной коррекции на уровне значимости p > 0,05 (см. Рисунок 1 для инструментов конвейера анализа). Это привело к функциональному графу связности с узлами и ребрами, представляющими важные активированные области и их значительную взаимосвязь, соответственно.
Затем результаты были перенесены в Network Based Statistic Toolbox (NBS; Zalesky et al., 2010) для изучения статистически значимых связей. Односторонний внутрисубъектный дисперсионный анализ ANOVA проводился с тремя условиями в качестве фактора внутри субъектов. Метод NBS был установлен для множественной коррекции на уровне значимости p > 0,05 (см. Рисунок 1 для инструментов конвейера анализа). Это привело к функциональному графу связности с узлами и ребрами, представляющими важные активированные области и их значительную взаимосвязь, соответственно.
Рисунок 1. Конвейер инструментов анализа. Первый шаг: индивидуальная сегментация МРТ на основе модели объемного проводника МКЭ была рассчитана с помощью программного обеспечения BESA MRI. Предварительная обработка временного ряда и усреднение вызванных полей были выполнены в исследовательском программном обеспечении BESA, а для реконструкции источника было применено решение LORETA. Второй шаг: набор инструментов статистики SPM в Matlab использовался для статистического анализа данных. Третий шаг: для анализа связности эквивалентные диполи тока были установлены для значимых регионов, выявленных в ходе предыдущего анализа.Четвертый шаг: набор инструментов HERMES в Matlab был использован для построения матрицы смежности 5 × 5 для каждого предмета и каждого условия на основе алгоритма взаимной информации. Пятый шаг: статистический анализ функциональной связи был выполнен в наборе инструментов NBS, и данные были перенесены в набор инструментов BrainNet для визуализации статически значимых сетей связи.
Третий шаг: для анализа связности эквивалентные диполи тока были установлены для значимых регионов, выявленных в ходе предыдущего анализа.Четвертый шаг: набор инструментов HERMES в Matlab был использован для построения матрицы смежности 5 × 5 для каждого предмета и каждого условия на основе алгоритма взаимной информации. Пятый шаг: статистический анализ функциональной связи был выполнен в наборе инструментов NBS, и данные были перенесены в набор инструментов BrainNet для визуализации статически значимых сетей связи.
Результаты
Исходное пространство
Слуховое вызванное поле N1 было априори , установленным в качестве интересующего временного окна.Среднеквадратичный временной ряд общего среднего для субъектов был вычислен в пространстве сенсора, чтобы отобразить временное окно в мс около априори установленного максимума слухового поля N1. На рисунке 2 показано среднее значение среднеквадратичных значений каждой звуковой категории, а также максимальные и минимальные значения их доверительных интервалов. Интервал 80–120 мс был определен для следующих реконструкций источников, где мы проводили статистику F- и t-контраста с помощью программного обеспечения SPM12.На рис. 3 и в таблице 1 показаны значимые кластеры, полученные с помощью контраста Musical ≠ Artificial ≠ Natural . Самый большой по размеру кластер включал части правой и левой лобной коры, а также части височной доли. Пик текущего кластера располагался в передней части правой височной коры, в самой дорсальной области верхней височной извилины (STG) ( x = 43, y = 14, z = −29 ; F (1, 20) = 13.1, размер кластера = 3128, p <0,001 FWE, скорректированный на уровне кластера). Второй кластер располагался в правой нижней теменной доле, с пиком в правой надмаргинальной извилине (SMG) (координаты: x = 56, y = -27, z = 27; F (1 , 20) = 12.80, размер кластера = 513, p <0.001 FWE, скорректированный на уровне кластера). Третий кластер располагался в задней части поясной извилины (PCC; перекрывается с кластером два на рисунке), который включает заднюю поясную извилину и медиальную часть теменной доли (координаты: x = 10, y = −52 , z = 39, F (1, 20) = 12.
Интервал 80–120 мс был определен для следующих реконструкций источников, где мы проводили статистику F- и t-контраста с помощью программного обеспечения SPM12.На рис. 3 и в таблице 1 показаны значимые кластеры, полученные с помощью контраста Musical ≠ Artificial ≠ Natural . Самый большой по размеру кластер включал части правой и левой лобной коры, а также части височной доли. Пик текущего кластера располагался в передней части правой височной коры, в самой дорсальной области верхней височной извилины (STG) ( x = 43, y = 14, z = −29 ; F (1, 20) = 13.1, размер кластера = 3128, p <0,001 FWE, скорректированный на уровне кластера). Второй кластер располагался в правой нижней теменной доле, с пиком в правой надмаргинальной извилине (SMG) (координаты: x = 56, y = -27, z = 27; F (1 , 20) = 12.80, размер кластера = 513, p <0.001 FWE, скорректированный на уровне кластера). Третий кластер располагался в задней части поясной извилины (PCC; перекрывается с кластером два на рисунке), который включает заднюю поясную извилину и медиальную часть теменной доли (координаты: x = 10, y = −52 , z = 39, F (1, 20) = 12. 42, размер кластера = 650, p <0,001 FWE, исправленное на уровне кластера).
42, размер кластера = 650, p <0,001 FWE, исправленное на уровне кластера).
Рис. 2. Среднеквадратичное значение больших средних значений слухового вызванного поля (AEF) для звуков Musical , Artificial и Natural . Графики иллюстрируют активацию мозга во временной области (от –100 до начала стимула до 300 мс после стимула) с 0 мс, представляющим начало стимула. Обозначены компоненты AEF P1, N1 и P2, а также временной интервал 80–120 мс.Средства различных типов звуков отмечены разными типами линий. Соответствующие им 95% доверительные интервалы показаны заштрихованными областями. Интересующий временной интервал составляет около N1 компонента слухового вызванного поля (80–120 мс).
Рис. 3. Значительная дифференциальная активация коры в пространстве источника на основе F-контраста. На рисунках показано статистическое параметрическое сопоставление, полученное на основе одностороннего анализа ANOVA статистики SPM для взаимодействия Musical ≠ Artificial ≠ Natural . Вверху, на левой стороне — среднесагиттальный вид левого полушария ; справа вверху сбоку срединно-сагиттальный вид правого полушария ; внизу представлен вид спереди. Активируются левая и правая префронтальная кора и передняя STG, а также супрамаргинальная извилина и PCC (перекрываются на рисунке). Цветовая шкала представляет F -значения на уровне значимости p <0,001 (с поправкой на FWE).
Вверху, на левой стороне — среднесагиттальный вид левого полушария ; справа вверху сбоку срединно-сагиттальный вид правого полушария ; внизу представлен вид спереди. Активируются левая и правая префронтальная кора и передняя STG, а также супрамаргинальная извилина и PCC (перекрываются на рисунке). Цветовая шкала представляет F -значения на уровне значимости p <0,001 (с поправкой на FWE).
Таблица 1. Значимые кластеры на основе F-контраста.
Для сравнений между группами результаты показали значительные различия для Музыкальный > Естественный и Искусственный > Естественный сравнений, как показано на Рисунке 4 и в Таблице 2, отображающих координаты и результаты сопоставления источников для обоих сравнений. . Более подробно, два значимых кластера были получены для музыкальных > естественных звуков.Самый большой кластер включал в себя области левой и правой лобной коры, а также височную кору и SMG. Пик текущего кластера располагался на SMG ( x = 56, y = −27, z = 27, t (20) = 5,05, размер кластера: 109960, p > 0,001 FWE исправлено на уровне кластера). Второй кластер располагался в PCC (координаты: x = 8, y = −54, z = 40, t (20) = 4.61, размер кластера = 3142, p <0.001 FWE исправлено на уровне кластера). Для контраста Искусственный > Естественный мы обнаружили два кластера, показывающих значительную дифференциальную активацию; один значимый пик был расположен на PCC (координаты: x = 10, y = -37, z = 39, t (20) = 4,09, размер кластера = 675, p = 0,001 FWE корректируется на уровне кластера), а другой — в медиальной префронтальной коре mPFC (координаты: x = 13, y = 39, z = 18, t (20) = 4.08, размер кластера = 1007, p <0,001 FWE, исправленное на уровне кластера).
Пик текущего кластера располагался на SMG ( x = 56, y = −27, z = 27, t (20) = 5,05, размер кластера: 109960, p > 0,001 FWE исправлено на уровне кластера). Второй кластер располагался в PCC (координаты: x = 8, y = −54, z = 40, t (20) = 4.61, размер кластера = 3142, p <0.001 FWE исправлено на уровне кластера). Для контраста Искусственный > Естественный мы обнаружили два кластера, показывающих значительную дифференциальную активацию; один значимый пик был расположен на PCC (координаты: x = 10, y = -37, z = 39, t (20) = 4,09, размер кластера = 675, p = 0,001 FWE корректируется на уровне кластера), а другой — в медиальной префронтальной коре mPFC (координаты: x = 13, y = 39, z = 18, t (20) = 4.08, размер кластера = 1007, p <0,001 FWE, исправленное на уровне кластера).
Рисунок 4. Значительная корковая активация в исходном пространстве при сравнении между субъектами. На рисунках показано статистическое параметрическое сопоставление, полученное на основе анализа t -тестов статистики SPM для сравнений Musical > Natural и Artificial > Natural . Вверху, на левой стороне — среднесагиттальный вид левого полушария ; справа вверху сбоку срединно-сагиттальный вид правого полушария ; внизу представлен вид спереди.Активируются левая и правая префронтальная кора и передняя STG, а также части левой и правой PCC и супрамаргинальная извилина (перекрывающиеся на рисунке), как показано на панели на слева ( Musical > Natural ). Панель справа ( Искусственный > Натуральный ) отображает активированные части правой префронтальной коры , надмаргинальной извилины и PCC.Цветовая шкала представляет t -значения на уровне значимости p <0,001 (с поправкой на FWE).
Значительная корковая активация в исходном пространстве при сравнении между субъектами. На рисунках показано статистическое параметрическое сопоставление, полученное на основе анализа t -тестов статистики SPM для сравнений Musical > Natural и Artificial > Natural . Вверху, на левой стороне — среднесагиттальный вид левого полушария ; справа вверху сбоку срединно-сагиттальный вид правого полушария ; внизу представлен вид спереди.Активируются левая и правая префронтальная кора и передняя STG, а также части левой и правой PCC и супрамаргинальная извилина (перекрывающиеся на рисунке), как показано на панели на слева ( Musical > Natural ). Панель справа ( Искусственный > Натуральный ) отображает активированные части правой префронтальной коры , надмаргинальной извилины и PCC.Цветовая шкала представляет t -значения на уровне значимости p <0,001 (с поправкой на FWE).
Таблица 2. Значимые кластеры межсубъектных сравнений.
Результаты подключения
Для анализа связности пять эквивалентных токовых диполей были установлены на пиках значительно различающихся активированных областей, полученных из F-контраста. Это PCC, aSTG, SMG и двусторонняя префронтальная кора.Было проведено три анализа связности, основанные на значительном взаимодействии, обнаруженном в анализе исходного пространства, p <0,05, исправлено NBS (Network-Based Statistics; Zalesky et al., 2010).
Для контраста Musical ≠ Artificial ≠ Natural результаты выявили соединения всех узлов, имеющих в общей сложности шесть ребер. Как показано на рисунке 5, края между PCC и правой передней верхней височной извилиной (aSTG), а также между левой вентральной mPFC и правой медиальной дорсальной префронтальной корой имели самую сильную активацию (на что указывает значение F ).Левый mPFC давал значительные дифференциальные связи со всеми узлами, расположенными в правом полушарии, и он был единственным, соединяющимся с SMG.
Рис. 5. Сеть функционального взаимодействия с F-контрастом. Проиллюстрированы статистически значимые связи между узлами для контраста Музыкальный ≠ Искусственный Natural Естественный . Узлы представляют собой важные области, выявленные анализом исходного пространства, которые были установлены как эквивалентные токовые диполи.Края взвешиваются и окрашиваются в соответствии с их силой соединения, как показано значением F цветовой шкалы. Сети значимы при p <0,05, исправлено NBS. На рисунке слева вверху и на изображен фронтальный фронтальный вид, на рисунке справа вверху на показана аксиальная плоскость при взгляде сверху, а на рисунке внизу показана правая сагиттальная плоскость.
В сравнении Music-vs.-Natural была получена значимая дифференциальная сеть из 10 ребер, при этом все узлы в сети были соединены между собой (c.f. Рисунок 6). Согласно значению t , улучшенная функциональная связь была продемонстрирована между правым mPFC и PCC, а также между PCC и aSTG. При меньшем значении t левый mPFC обеспечил значительную взаимосвязь с правым PFC, aSTG, SMG и PCC. Более слабые соединения были получены между SMG и aSTG, PCC, а также левым mPFC.
При меньшем значении t левый mPFC обеспечил значительную взаимосвязь с правым PFC, aSTG, SMG и PCC. Более слабые соединения были получены между SMG и aSTG, PCC, а также левым mPFC.
Рисунок 6. Значимая функциональная сеть связи для межпредметных сравнений.Проиллюстрированы статистически значимые связи между узлами для сравнений Musical > Natural и Artificial > Natural соответственно. Узлы представляют собой важные области, выявленные анализом исходного пространства, которые были установлены как эквивалентные токовые диполи. Края взвешиваются и окрашиваются в соответствии с силой соединения, как показано значением t цветовой шкалы. Сети значимы при p <0.05, исправлено NBS. Рисунок вверху-слева, на обеих, левой и правой панелях, изображает фронтальный коронарный вид, вверху-справа показывает осевую плоскость, если смотреть сверху, а рисунок внизу иллюстрирует правую сагиттальную плоскость.
Что касается сравнений искусственного и естественного (см. Рисунок 6), результаты показали значительные дифференциальные взаимосвязи между всеми узлами, имеющими в общей сложности девять ребер. В частности, краевое соединение префронтального межполушария и соединение между узлами aSTG и PCC были наиболее выраженными согласно значению t .Края с меньшей прочностью были выявлены для остальных соединений, но между PCC и SMG не было обнаружено значительных соединений.
Обсуждение
В данном исследовании изучались нейронные реакции при обработке различных категорий звуков с помощью МЭГ. Кроме того, анализ функциональной связности на основе алгоритма MI использовался для изображения дифференциального коннектома областей, участвующих в различении звуковых категорий Natural , Musical и Artificial sound.Полученные результаты продемонстрировали статистически значимые различия между различными звуковыми условиями в верхней височной коре, задней поясной извилине, нижней теменной и двусторонней префронтальной коре между 80 и 120 мс после появления стимула. Прямые сравнения по различным категориям показали, что звуки Musical и Artificial продемонстрировали статистически значимые различия и более высокую активацию мозга в пространстве источников и анализе связности по сравнению с категорией звуков Natural .
Прямые сравнения по различным категориям показали, что звуки Musical и Artificial продемонстрировали статистически значимые различия и более высокую активацию мозга в пространстве источников и анализе связности по сравнению с категорией звуков Natural .
Наш вывод о том, что области теменной коры демонстрируют значительную категориальную модуляцию активации, что подтверждается анализом контраста, это согласуется с несколькими исследованиями нейровизуализации, изучающими организацию кортикального слухового восприятия (Lewis et al., 2005; Murray et al., 2006 ; Staeren et al., 2009). В нашем исследовании мы обнаружили две области в теменной доле, которые участвуют в дифференциальной обработке оцениваемых звуковых категорий, а именно PCC и правая нижняя теменная кора (IPC).Пик кластера IPC находится в SMG, которая является частью соматосенсорной коры (Buccino et al., 2001). Фактически, предполагается, что он участвует в представлении действия и, в частности, в мысленном представлении движения (Chong et al. , 2008; Tunik et al., 2008). В рамках этой интерпретации любое событие или стимулы, содержащие действия, будут задействовать области, связанные с моторикой, даже в отсутствие тактильной стимуляции (Nilsson et al., 2000). Дополнительные свидетельства связывают SMG с системой зеркальных нейронов, системой, участвующей в имитации и идентификации действий других людей (Carlson, 2012).На этом основании было предложено, чтобы правильный SMG был особенно задействован в моторном планировании, даже когда действия просто наблюдаются, а не обязательно выполняются (Chong et al., 2008). Кажется, что в ментальном представлении информация о том, как манипулируют, визуализируют инструмент или как воспроизводится звук, извлекается и интегрируется.
, 2008; Tunik et al., 2008). В рамках этой интерпретации любое событие или стимулы, содержащие действия, будут задействовать области, связанные с моторикой, даже в отсутствие тактильной стимуляции (Nilsson et al., 2000). Дополнительные свидетельства связывают SMG с системой зеркальных нейронов, системой, участвующей в имитации и идентификации действий других людей (Carlson, 2012).На этом основании было предложено, чтобы правильный SMG был особенно задействован в моторном планировании, даже когда действия просто наблюдаются, а не обязательно выполняются (Chong et al., 2008). Кажется, что в ментальном представлении информация о том, как манипулируют, визуализируют инструмент или как воспроизводится звук, извлекается и интегрируется.
Помимо теменной коры, наши результаты по связности дополнительно подчеркнули участие префронтальной коры и STG в обработке различных категорий звуков.Значительная функциональная связь между префронтальной корой и теменной корой была продемонстрирована в прошлом, предполагая, что эта сеть является частью сети рабочей памяти, связывающей восприятие и познание более высокого порядка.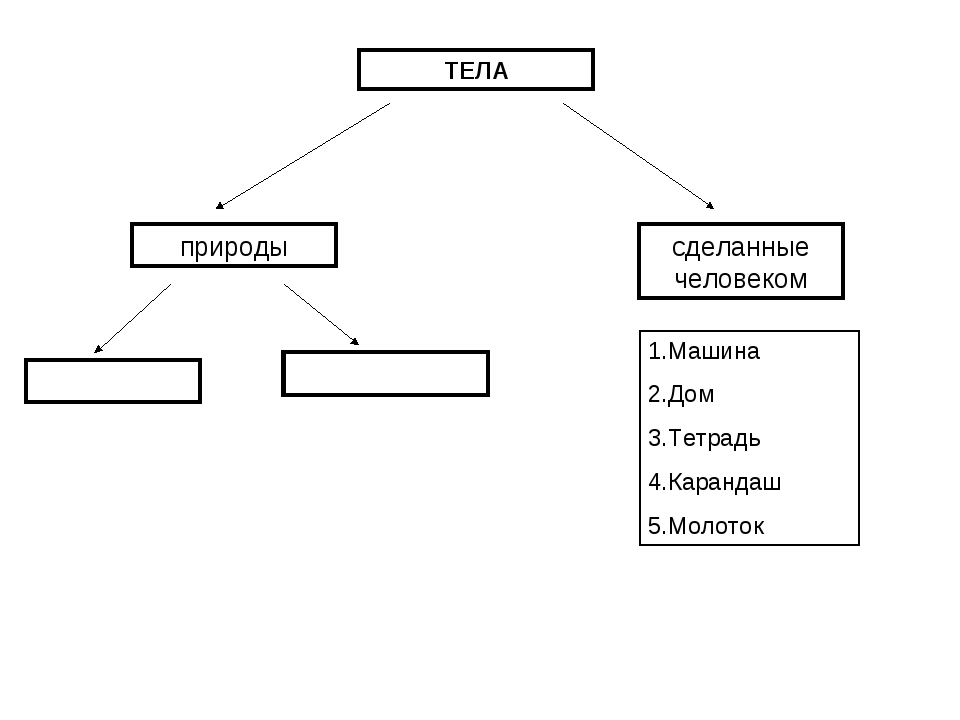 Это довольно многомодально и касается обработки как слуховых, так и визуальных объектов (Husain et al., 2004; Husain and Horwitz, 2006). Что касается STG, большинство исследований нейровизуализации показали значительное участие этой области в слуховой обработке. Однако, согласно метаанализу Arnott et al.(2004; включая 38 исследований различных методов нейровизуализации) кажется, что передняя область STG, как показано здесь, в большей степени связана с различением звуков. В соответствии с нашими результатами по связности, было обнаружено, что область STG значительно взаимодействует с префронтальной корой во время задачи распознавания категорий (Husain and Horwitz, 2006). Тот факт, что наш паттерн связности был латерализован в правом полушарии, согласуется с предположением, что это полушарие активируется сильнее, когда представлены неречевые или неживые стимулы (см. Zatorre et al., 2002; Терваниеми и Хугдал, 2003). Это основано не только на исследованиях поражений мозга, но и на исследованиях нейровизуализации здоровых участников (Belin and Zatorre, 2003; Murray et al.
Это довольно многомодально и касается обработки как слуховых, так и визуальных объектов (Husain et al., 2004; Husain and Horwitz, 2006). Что касается STG, большинство исследований нейровизуализации показали значительное участие этой области в слуховой обработке. Однако, согласно метаанализу Arnott et al.(2004; включая 38 исследований различных методов нейровизуализации) кажется, что передняя область STG, как показано здесь, в большей степени связана с различением звуков. В соответствии с нашими результатами по связности, было обнаружено, что область STG значительно взаимодействует с префронтальной корой во время задачи распознавания категорий (Husain and Horwitz, 2006). Тот факт, что наш паттерн связности был латерализован в правом полушарии, согласуется с предположением, что это полушарие активируется сильнее, когда представлены неречевые или неживые стимулы (см. Zatorre et al., 2002; Терваниеми и Хугдал, 2003). Это основано не только на исследованиях поражений мозга, но и на исследованиях нейровизуализации здоровых участников (Belin and Zatorre, 2003; Murray et al. , 2006).
, 2006).
Другая возможная интерпретация наших результатов состоит в том, что обработка музыкальных и искусственных звуков в большей степени включает сеть режима по умолчанию (DMN). Согласно нашим результатам, область PCC играет центральную роль в DMN (для обзора см. Leech and Sharp, 2013) с нижней теменной долей, префронтальной корой и височной корой (однако, более медиальными структурами). в качестве основных узлов для внутренних связей (Raichle et al., 2001). Активация DMN в основном наблюдалась в условиях отсутствия задач, когда не требуется внимания к внешним стимулам (см. Обзор в Buckner et al., 2008). Однако также наблюдается повышенная активность во время когнитивной обработки, которая требует внутреннего внимания, например, восстановления памяти и планирования (Spreng, 2012). С этой точки зрения, обработка звуков искусственных объектов может потребовать более нисходящей обработки информации, чем звуки живых объектов. Более того, PCC был тесно связан с эмоциональной обработкой и состоянием возбуждения с более высокими реакциями и крупномасштабной функциональной связностью в состоянии возбуждения и снижался в состоянии сна (Leech and Sharp, 2013). Таким образом, стимуляция музыкальными и искусственными звуками может вызывать сильные эмоции по сравнению с естественными звуками. Тем не менее, отсутствие соответствующих поведенческих измерений не позволяет провести дальнейшую интерпретацию этого предположения, и это выходит за рамки данной статьи. В будущем поведенческий тест, оценивающий уровни эмоционального возбуждения к различным звукам, даст нам некоторое представление о том, влияет ли этот фактор существенно на дифференциальную активацию мозга.
Таким образом, стимуляция музыкальными и искусственными звуками может вызывать сильные эмоции по сравнению с естественными звуками. Тем не менее, отсутствие соответствующих поведенческих измерений не позволяет провести дальнейшую интерпретацию этого предположения, и это выходит за рамки данной статьи. В будущем поведенческий тест, оценивающий уровни эмоционального возбуждения к различным звукам, даст нам некоторое представление о том, влияет ли этот фактор существенно на дифференциальную активацию мозга.
Наша основная гипотеза заключалась в том, что искусственные звуки будут отличаться от звуков Natural из-за мысленного представления моторных характеристик, которые действительно были более выраженными.Далее мы стремились выяснить, применимы ли статистические различия к сравнению между Musical и Artificial , как сообщалось ранее; основанный на предположении, что звуки повседневно используемых предметов могут вызывать больше реакции на действие по сравнению с музыкальными инструментами (у не музыкантов). Наши результаты не выявили каких-либо существенных различий в анализе исходного пространства, однако в анализе связности, даже несмотря на то, что Музыкальное против естественного и Искусственное против естественного продемонстрировали аналогичные модели связности, силу взаимосвязей (как учитывая толщину кромки) был другим.Например, в сети «Искусственный против естественного» межполушарный префронтальный край и край STG-PCC продемонстрировали более сильные взаимосвязи между остальными, тогда как сеть «Музыкальное против естественного» дала более распределенную интенсивность по всей сети. соединения с меньшими значениями. Предыдущее исследование (De Lucia et al., 2009) показало различие между музыкальными и инструментальными звуками только после появления стимула в 300 мс. Исходя из этого, может случиться так, что различение более широких категорий звуков, таких как живые и искусственные звуки, уже происходит на ранних ответах, хотя за различением подкатегорий могут следовать более поздние ответы.
Наши результаты не выявили каких-либо существенных различий в анализе исходного пространства, однако в анализе связности, даже несмотря на то, что Музыкальное против естественного и Искусственное против естественного продемонстрировали аналогичные модели связности, силу взаимосвязей (как учитывая толщину кромки) был другим.Например, в сети «Искусственный против естественного» межполушарный префронтальный край и край STG-PCC продемонстрировали более сильные взаимосвязи между остальными, тогда как сеть «Музыкальное против естественного» дала более распределенную интенсивность по всей сети. соединения с меньшими значениями. Предыдущее исследование (De Lucia et al., 2009) показало различие между музыкальными и инструментальными звуками только после появления стимула в 300 мс. Исходя из этого, может случиться так, что различение более широких категорий звуков, таких как живые и искусственные звуки, уже происходит на ранних ответах, хотя за различением подкатегорий могут следовать более поздние ответы. В будущем было бы интересно исследовать более поздние временные интервалы, в которых могут потребоваться концептуальные процессы более высокого уровня для выделения искусственных подкатегорий. Следует отметить, что категория Искусственный в нашем исследовании содержала звуки, которые, как правило, являются искусственными объектами, однако некоторые из них были менее манипулятивными (например, сирена скорой помощи). Отсутствие более строгой категоризации, основанной на поведенческой оценке, которая разделяла бы объекты на основе знакомства и частоты использования, могло ограничить нашу интерпретацию существенных различий между звуковыми категориями.Изучение областей коры головного мозга в ответ на знакомство со звуком, узнаваемость и внимание может дать представление о роли, которую «какой» слуховой нейрональный путь играет в обработке звука. Предыдущие исследования показали важность звуковой новизны, требований задач и внимания для распознавания звука (Levy et al., 2001, 2003).
В будущем было бы интересно исследовать более поздние временные интервалы, в которых могут потребоваться концептуальные процессы более высокого уровня для выделения искусственных подкатегорий. Следует отметить, что категория Искусственный в нашем исследовании содержала звуки, которые, как правило, являются искусственными объектами, однако некоторые из них были менее манипулятивными (например, сирена скорой помощи). Отсутствие более строгой категоризации, основанной на поведенческой оценке, которая разделяла бы объекты на основе знакомства и частоты использования, могло ограничить нашу интерпретацию существенных различий между звуковыми категориями.Изучение областей коры головного мозга в ответ на знакомство со звуком, узнаваемость и внимание может дать представление о роли, которую «какой» слуховой нейрональный путь играет в обработке звука. Предыдущие исследования показали важность звуковой новизны, требований задач и внимания для распознавания звука (Levy et al., 2001, 2003). Однако другие утверждают, что различение живых и созданных руками объектов присутствует независимо от поведенческих навыков и, следовательно, от степени знакомства (De Lucia et al., 2012), когда правильно классифицированные звуки сравнивались с неправильно категоризированными звуками, а также независимо от сознания (Cossy et al., 2014) при обследовании коматозных пациентов. Этот вопрос все еще обсуждается в литературе и должен быть рассмотрен в будущих исследованиях.
Однако другие утверждают, что различение живых и созданных руками объектов присутствует независимо от поведенческих навыков и, следовательно, от степени знакомства (De Lucia et al., 2012), когда правильно классифицированные звуки сравнивались с неправильно категоризированными звуками, а также независимо от сознания (Cossy et al., 2014) при обследовании коматозных пациентов. Этот вопрос все еще обсуждается в литературе и должен быть рассмотрен в будущих исследованиях.
Возможное ограничение текущего исследования может заключаться в том, что категоризация сложных звуков может быть затруднена из-за физических различий стимулов (Belin et al., 2000; Staeren et al., 2009), что по своей цели здесь не контролировалось, во избежание искажения качества звука.Однако естественные звуки отличаются от звуков, созданных человеком, и более синтетических по своей природе, которыми мы не можем управлять. Согласно обзору Theunissen и Elie (2014), естественные звуки состоят из статистических свойств, которые следуют соотношению степенного закона и оптимально кодируются в восходящей системе слуховой обработки, в отличие от звуков с более случайными и плоскими спектрами мощности огибающей (такими как звуки в категории Искусственное). Тем не менее подобная категоризация звуков применялась в прошлом, и наши результаты согласуются (Murray et al., 2006).
Тем не менее подобная категоризация звуков применялась в прошлом, и наши результаты согласуются (Murray et al., 2006).
Результаты нашего исследования показывают, что диссоциация между живыми и созданными руками объектами основана на различных нейронных процессах. Однако причина этого явления до сих пор остается под вопросом. С другой точки зрения, возможно, звуки были концептуально специализированы на обработке различных категорий звуков из-за эволюционной адаптации; теория, которая также была сильно аргументирована в пользу зрительной системы (Caramazza and Mahon, 2003). В соответствии с этим, отдельные корковые пути, соответствующие различным категориям звуков, были разработаны аналогично звукам окружающей среды, которые люди испытывали на протяжении многих лет.Это согласуется с ранее упомянутым обзором, который предлагает оптимальное нейронное кодирование естественных звуков по сравнению со звуками созданных руками человека машин (Theunissen and Elie, 2014). С этой точки зрения, прослушивание звуков природы вызовет более слабую активацию мозга по сравнению с более «современными» звуками инструментов (например, звонком телефона), поскольку мы эволюционно более приспособлены к естественным звукам, а это, в свою очередь, потребует меньше когнитивные усилия (Buckner et al. , 2008).Это, в свою очередь, дало бы некоторое объяснение вышеупомянутым открытиям, согласно которым искусственные звуки показали более сильную активацию мозга по сравнению с «живыми» объектами. Кроме того, функциональные карты связности показали, что в категории искусственного происхождения могут быть «ключевые» связи для «Искусственное против естественного» по сравнению с «Музыкальное против естественного» , даже если узлы остались теми же самыми для обоих сравнений. . Это также может объяснить случаи семантических нарушений, когда пациенты страдают от очень специфической категории, в то время как остальные категории в той же области не затрагиваются (McCarthy and Warrington, 2016; Muhammed et al., 2018). Следовательно, кажется очень вероятным, что восприятие слуховых объектов опирается на крупномасштабную распределенную систему, которая следует различным нейрональным путям, диссоциированным на основе веса, который каждый узел имеет в сети. Исходя из наших выводов, на это предположение нельзя однозначно ответить, хотя оно дает повод для предстоящего исследования.
, 2008).Это, в свою очередь, дало бы некоторое объяснение вышеупомянутым открытиям, согласно которым искусственные звуки показали более сильную активацию мозга по сравнению с «живыми» объектами. Кроме того, функциональные карты связности показали, что в категории искусственного происхождения могут быть «ключевые» связи для «Искусственное против естественного» по сравнению с «Музыкальное против естественного» , даже если узлы остались теми же самыми для обоих сравнений. . Это также может объяснить случаи семантических нарушений, когда пациенты страдают от очень специфической категории, в то время как остальные категории в той же области не затрагиваются (McCarthy and Warrington, 2016; Muhammed et al., 2018). Следовательно, кажется очень вероятным, что восприятие слуховых объектов опирается на крупномасштабную распределенную систему, которая следует различным нейрональным путям, диссоциированным на основе веса, который каждый узел имеет в сети. Исходя из наших выводов, на это предположение нельзя однозначно ответить, хотя оно дает повод для предстоящего исследования. Анализ связности, включающий также общие активированные области, может дать лучшую картину для этого предположения, однако из-за того, что это требует другого анализа, это рекомендуется для будущего исследования.
Анализ связности, включающий также общие активированные области, может дать лучшую картину для этого предположения, однако из-за того, что это требует другого анализа, это рекомендуется для будущего исследования.
В целом факт, что обработка слуховых стимулов затрагивает области за пределами слуховой коры, такие как передняя височная и лобная доли (Maeder et al., 2001; Alain et al., 2008), хорошо документирован. Подобно результатам нашего анализа корковой сети, недавние исследования нейровизуализации подтверждают, что сеть лобно-височно-теменных областей вносит вклад в семантическую обработку (для обзора см. Thompson-Schill, 2003; Patterson et al., 2002). Было предложено связать эту сеть с восприятием идентификации как слуховых, так и визуальных объектов (Goll et al., 2010; Хантер и др., 2010; Брефчински-Льюис и Льюис, 2017). Однако то, что еще не очень хорошо задокументировано, — это то, как эти реакции мозга функционально связаны. Наше исследование связности подкрепляет связность областей мозга при распознавании звука. Чтобы получить лучшее представление о слуховой категоризации мозга, недостаточно исследовать только активированные области по отдельности, но лучше понять, как эти области взаимодействуют. В этом аспекте мы считаем, что наши результаты ценны для лучшего понимания человеческого мозга при распознавании звука.
Чтобы получить лучшее представление о слуховой категоризации мозга, недостаточно исследовать только активированные области по отдельности, но лучше понять, как эти области взаимодействуют. В этом аспекте мы считаем, что наши результаты ценны для лучшего понимания человеческого мозга при распознавании звука.
Заключение
Настоящее исследование продемонстрировало усиленную мозговую сеть искусственных звуков ( музыкальных и искусственных ) по сравнению с естественными звуками. До сих пор в литературе представлены простые карты активации мозга. Мы дополнительно показали, как эти дифференцированно активированные области мозга функционально связаны и связаны с соответствующими когнитивными процессами. Мы воспроизвели предыдущие результаты, подтверждающие участие других модальностей, помимо слуховых, для участия в обработке звуковых стимулов, как только они достигают уровня репрезентации объекта.Это, в свою очередь, похоже, основано на семантической категоризации стимула, следуя различным нейронным путям для живых и созданных руками объектов. В дополнение к предыдущим исследованиям, в которых изучалась только корковая активация различных категорий звуков, мы продемонстрировали значительные различия в функциональной связности между корковыми источниками, участвующими в обработке различных категорий звука.
В дополнение к предыдущим исследованиям, в которых изучалась только корковая активация различных категорий звуков, мы продемонстрировали значительные различия в функциональной связности между корковыми источниками, участвующими в обработке различных категорий звука.
Заявление о доступности данных
Наборы данных, созданные для этого исследования, доступны по запросу соответствующему автору.
Заявление об этике
Исследования с участием людей были рассмотрены и одобрены медицинским факультетом Мюнстерского университета. Пациенты / участники предоставили письменное информированное согласие на участие в этом исследовании.
Авторские взносы
VS внесла свой вклад в набор данных, анализ данных и написала первый черновик рукописи. EP и NC участвуют в статистическом анализе. КМ и AW задействованы в информационном обеспечении.CD, DK и CP внесли свой вклад в редактирование рукописи. Все авторы прочитали и одобрили представленный вариант рукописи.
Финансирование
Эта работа была поддержана Deutsche Forschungsgemeinschaft (DFG; PA 392 / 16-1, DO-711 / 10-1).
Конфликт интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Авторы хотели бы поблагодарить субъектов за их сотрудничество и технических специалистов Института биомагнетизма и биосигнала за поддержку сбора данных.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fnins.2019.01052/full#supplementary-material
Сноски
Список литературы
Ален, К., Макдональд, К. Л., Ковачевич, Н., и Макинтош, А. Р. (2008). Пространственно-временной анализ слуховой «что» и «где» рабочей памяти. Cereb. Cortex 19, 305–314. DOI: 10.1093 / cercor / bhn082
DOI: 10.1093 / cercor / bhn082
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Арнотт С. Р., Биннс М. А., Грейди К. Л. и Ален К. (2004). Оценка модели двойного слухового пути у людей. Neuroimage 22, 401–408. DOI: 10.1016 / j.neuroimage.2004.01.014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Асплунд, К. Л., Тодд, Дж. Дж., Снайдер, А. П., и Маруа, Р. (2010). Центральная роль боковой префронтальной коры в целенаправленном и стимулированном внимании. Nat. Neurosci. 13, 507–512. DOI: 10.1038 / nn.2509
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Белин П., Заторре Р. Дж., Лафай П., Ахад П. и Пайк Б. (2000). Голососелективные области в слуховой коре человека. Природа 403, 309–312. DOI: 10.1038 / 35002078
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Брефчински-Льюис, Дж. А., и Льюис, Дж. У. (2017). Восприятие слухового объекта: нейробиологическая модель и проспективный обзор. Neuropsychologia 105, 223–242. DOI: 10.1016 / j.neuropsychologia.2017.04.034
Neuropsychologia 105, 223–242. DOI: 10.1016 / j.neuropsychologia.2017.04.034
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Брегман А.С. (2017). «Задавая вопрос« зачем »в слуховом восприятии», в Perceptual Organization , ред. М. Кубови и Дж. Померанц, (Hillsdale, NJ: Routledge), 99–118. DOI: 10.4324 / 9781315512372-4
CrossRef Полный текст | Google Scholar
Буччино, Г., Бинкофски, Ф., Финк, Г. Р., Фадига, Л., Fogassi, L., Gallese, V., et al. (2001). Наблюдение за действием активирует премоторную и теменную области соматотопическим образом: исследование фМРТ. Eur. J. Neurosci. 13, 400–404.
Google Scholar
Буллмор, Э., Спорнс, О. (2009). Сложные сети мозга: теоретико-графический анализ структурных и функциональных систем. Nat. Rev. Neurosci. 10, 186–198. DOI: 10.1038 / nrn2575
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Карамацца, А., и Махон, Б. З. (2003). Организация концептуального знания: свидетельства категорийно-специфических семантических дефицитов. Trends Cogn. Sci. 7, 354–361. DOI: 10.1016 / s1364-6613 (03) 00159-1
З. (2003). Организация концептуального знания: свидетельства категорийно-специфических семантических дефицитов. Trends Cogn. Sci. 7, 354–361. DOI: 10.1016 / s1364-6613 (03) 00159-1
CrossRef Полный текст | Google Scholar
Карлсон, Н. Р. (2012). Физиология поведения , 11-е изд. Лондон: Пирсон.
Google Scholar
Чонг, Т. Т. Дж., Каннингтон, Р., Уильямс, М. А., Канвишер, Н., и Маттингли, Дж. Б. (2008). ФМРТ позволяет выявить зеркальные нейроны в нижней теменной коре головного мозга человека. Curr. Биол. 18, 1576–1580. DOI: 10.1016 / j.cub.2008.08.068
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кларк, С., Беллманн, А., Меули, Р. А., Ассал, Г., и Стек, А. Дж. (2000). Слуховая агнозия и слуховой пространственный дефицит после поражений левого полушария: доказательства различных путей обработки. Neuropsychologia 38, 797–807. DOI: 10.1016 / s0028-3932 (99) 00141-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Косси, Н. , Цовара, А., Симонин, А., Россетти, А., и Де Люсия, М. (2014). Четкое различение ответов ЭЭГ на категории звуков окружающей среды в ранней коме. Фронт. Psychol. 5: 155. DOI: 10.3389 / fpsyg.2014.00155
, Цовара, А., Симонин, А., Россетти, А., и Де Люсия, М. (2014). Четкое различение ответов ЭЭГ на категории звуков окружающей среды в ранней коме. Фронт. Psychol. 5: 155. DOI: 10.3389 / fpsyg.2014.00155
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Де Люсия, М., Камен, К., Кларк, С., и Мюррей, М. М. (2009). Роль действий в различении слуховых объектов. Neuroimage 48, 475–485. DOI: 10.1016 / j.neuroimage.2009.06.041
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Де Люсия, М., Цовара, А., Бернаскони, Ф., Шпирер, Л., и Мюррей, М. М. (2012). Принятие решений на основе слухового восприятия на основе семантической категоризации звуков окружающей среды. Neuroimage 60, 1704–1715. DOI: 10.1016 / j.neuroimage.2012.01.131
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ганцетти, М., Вендерот, Н., и Мантини, Д. (2016). Коррекция неоднородности интенсивности структурных МР-изображений: управляемый данными подход для определения входных параметров алгоритма. Фронт. Нейроинформ. 10:10. DOI: 10.3389 / fninf.2016.00010
Фронт. Нейроинформ. 10:10. DOI: 10.3389 / fninf.2016.00010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Голл, Дж. К., Костыль, С. Дж., И Уоррен, Дж. Д. (2010). Центральные слуховые расстройства: к нейропсихологии слуховых объектов. Curr. Мнение. Neurol. 23, 617–627. DOI: 10.1097 / WCO.0b013e32834027f6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Греч Р., Кассар Т., Маскат Дж., Камиллери, К. П., Фабри, С.Г., Зервакис М. и др. (2008). Обзор решения обратной задачи анализа источников ЭЭГ. J. Neuroeng. Rehabil. 5, 25. DOI: 10.1186 / 1743-0003-5-25
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хикерен, Х. Р., Марретт, С., и Унгерлейдер, Л. Г. (2008). Нейронные системы, которые опосредуют принятие решений человеком. Nat. Rev. Neurosci. 9, 467–479. DOI: 10.1038 / nrn2374
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хаддлстон, W. Э., Льюис, Дж. У., Финни, Р. Э., и ДеЙо, Э. А. (2008). Слуховое и визуальное видимое движение, основанное на внимании, имеет функциональные параллели. Восприятие. Психофизика. 70, 1207–1216. DOI: 10.3758 / PP.70.7.1207
Э., Льюис, Дж. У., Финни, Р. Э., и ДеЙо, Э. А. (2008). Слуховое и визуальное видимое движение, основанное на внимании, имеет функциональные параллели. Восприятие. Психофизика. 70, 1207–1216. DOI: 10.3758 / PP.70.7.1207
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хантер, М. Д., Эйкхофф, С. Б., Фазан, Р. Дж., Дуглас, М. Дж., Уоттс, Г. Р., Фэрроу, Т. Ф. и др. (2010). Состояние покоя: субъективное восприятие формируется контекстной модуляцией слуховой связи. Neuroimage 53, 611–618. DOI: 10.1016 / j.neuroimage.2010.06.053
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хусейн, Ф. Т., и Хорвиц, Б. (2006). Фреймворк экспериментального нейромоделирования для понимания обработки слуховых объектов: интеграция данных в различных масштабах. J. Physiol. Париж 100, 133–141. DOI: 10.1016 / j.jphysparis.2006.09.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хусейн, Ф.Т., Тагамец, М. А., Фромм, С.Дж., Браун, А.Р., и Хорвиц, Б. (2004). Связь нейрональной динамики для обработки слуховых объектов с нейровизуализационной активностью: компьютерное моделирование и исследование фМРТ. Neuroimage 21, 1701–1720. DOI: 10.1016 / j.neuroimage.2003.11.012
А., Фромм, С.Дж., Браун, А.Р., и Хорвиц, Б. (2004). Связь нейрональной динамики для обработки слуховых объектов с нейровизуализационной активностью: компьютерное моделирование и исследование фМРТ. Neuroimage 21, 1701–1720. DOI: 10.1016 / j.neuroimage.2003.11.012
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Иль Н., Берг П. и Шерг М. (2002). Коррекция артефактов текущей ЭЭГ с помощью пространственных фильтров на основе топографии артефактов и сигналов мозга. J. Clin. Neurophysiol. 19, 113–124.
Google Scholar
Лакади, К. М., Фулбрайт, Р. К., Радживан, Н., Констебль, Р. Т., и Пападеметрис, X. (2008). Более точные координаты Талаираха для нейровизуализации с использованием нелинейной регистрации. Neuroimage 42, 717–725. DOI: 10.1016 / j.neuroimage.2008.04.240
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Леви, Д.А., Гранот, Р., и Бентин, С. (2001). Специфика обработки стимулов человеческого голоса: электрофизиологические данные. Neuroreport 12, 2653–2657. DOI: 10.1097 / 00001756-200108280-00013
Neuroreport 12, 2653–2657. DOI: 10.1097 / 00001756-200108280-00013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Леви, Д.А., Гранот, Р., и Бентин, С. (2003). Нейронная чувствительность к человеческим голосам: ERP-свидетельство влияния задачи и внимания. Психофизиология 40, 291–305. DOI: 10.1111 / 1469-8986.00031
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Льюис, Дж. У., Брефчинский, Дж. А., Финни, Р. Э., Джаник, Дж. Дж., И ДеЙо, Э. А. (2005). Четкие корковые пути для обработки звуков инструмента и животных. J. Neurosci. 25, 5148–5158. DOI: 10.1523 / jneurosci.0419-05.2005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Льюис, Дж. У., Вайтман, Ф. Л., Брефчински, Дж. А., Финни, Р. Е., Биндер, Дж. Р., и ДеЙо, Э. А. (2004). Области человеческого мозга, участвующие в распознавании звуков окружающей среды. Cereb. Cortex 14, 1008–1021. DOI: 10.1093 / cercor / bhh061
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мейдер П. П., Меули, Р. А., Адриани, М., Беллманн, А., Форнари, Э., Тиран, Дж. П. и др. (2001). Определенные пути, участвующие в распознавании и локализации звука: исследование человека с помощью фМРТ. Нейроизображение 14, 802–816. DOI: 10.1006 / nimg.2001.0888
П., Меули, Р. А., Адриани, М., Беллманн, А., Форнари, Э., Тиран, Дж. П. и др. (2001). Определенные пути, участвующие в распознавании и локализации звука: исследование человека с помощью фМРТ. Нейроизображение 14, 802–816. DOI: 10.1006 / nimg.2001.0888
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Маккарти, Р. А., Уоррингтон, Э. К. (2016). Прошлое, настоящее и перспективы: размышления 40 лет спустя от избирательного нарушения семантической памяти (Warrington, 1975). Q.J. Exp. Psychol. 69, 1941–1968. DOI: 10.1080 / 17470218.2014.980280
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мендес, М. Ф. (2001). Генерализованная слуховая агнозия с нарушением распознавания музыки у левши. Разбор случая с правым височным инсультом. Cortex 37, 139–150. DOI: 10.1016 / s0010-9452 (08) 70563-x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мухаммед, Л., Харди, К. Дж., Рассел, Л. Л., Маршалл, К. Р., Кларк, К. Н., Бонд, Р. Л. и др. (2018). Агнозия для криков птиц. Neuropsychologia 113, 61–67. DOI: 10.1016 / j.neuropsychologia.2018.03.024
Л., Маршалл, К. Р., Кларк, К. Н., Бонд, Р. Л. и др. (2018). Агнозия для криков птиц. Neuropsychologia 113, 61–67. DOI: 10.1016 / j.neuropsychologia.2018.03.024
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мюррей М. М., Камен К., Андино С. Л. Г., Бовет П. и Кларк С. (2006). Быстрое распознавание мозгом звуков предметов. J. Neurosci. 26, 1293–1302. DOI: 10.1523 / jneurosci.4511-05.2006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Нильссон, Л.Г., Нюберг, Л., Клингберг, Т., Оберг, К., Перссон, Дж., И Роланд, П. Е. (2000). Активность в двигательных областях при запоминании событий действия. Neuroreport 11, 2199–2201. DOI: 10.1097 / 00001756-200007140-00027
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Niso, G., Bruña, R., Pereda, E., Gutiérrez, R., Bajo, R., Maestú, F., et al. (2013). HERMES: к интегрированному набору инструментов для характеристики функциональной и эффективной связи мозга. Нейроинформатика 11, 405–434.DOI: 10.1007 / s12021-013-9186-1
Нейроинформатика 11, 405–434.DOI: 10.1007 / s12021-013-9186-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Окамото, Х., Стракке, Х., и Пантев, К. (2008). Нейронные взаимодействия внутри и за пределами критической полосы, вызванные двумя одновременно представленными узкополосными шумами: магнитоэнцефалографическое исследование. Неврология 151, 913–920. DOI: 10.1016 / j.neuroscience.2007.11.038
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Олдфилд, Р. К. (1971).Оценка и анализ руки: Эдинбургская описи. Neuropsychologia 9, 97–113. DOI: 10.1016 / 0028-3932 (71) -4
CrossRef Полный текст | Google Scholar
Пантев, К., Эльберт, Т., Макейг, С., Хэмпсон, С., Юлитц, К., и Хок, М. (1993). Взаимосвязь переходных и установившихся слуховых вызванных полей. Электроэнцефалогр. Clin. Neurophysiol. 88, 389–396. DOI: 10.1016 / 0168-5597 (93)
-hPubMed Аннотация | CrossRef Полный текст | Google Scholar
Пантев, С. , Окамото, Х., Росс, Б., Столл, В., Чюрлиа-Гай, Э., Какиги, Р. и др. (2004). Боковое торможение и привыкание слуховой коры человека. Eur. J. Neurosci. 19, 2337–2344. DOI: 10.1111 / j.0953-816x.2004.03296.x
, Окамото, Х., Росс, Б., Столл, В., Чюрлиа-Гай, Э., Какиги, Р. и др. (2004). Боковое торможение и привыкание слуховой коры человека. Eur. J. Neurosci. 19, 2337–2344. DOI: 10.1111 / j.0953-816x.2004.03296.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Параскевопулос, Э., Чалас, Н., Карцидис, П., Воллбринк, А., и Бамидис, П. (2018). Статистическое изучение мультисенсорных закономерностей улучшено у музыкантов: исследование MEG. NeuroImage 175, 150–160.DOI: 10.1016 / j.neuroimage.2018.04.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Параскевопулос, Э., Кранебург, А., Херхольц, С. К., Бамидис, П. Д., и Пантев, К. (2015). Музыкальный опыт связан с измененной функциональной связью во время аудиовизуальной интеграции. Proc. Natl. Акад. Sci. 112, 12522–12527. DOI: 10.1073 / pnas.1510662112
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Паскуаль-Марки, Р. Д., Мишель, К. М. и Леманн Д. (1994). Электромагнитная томография низкого разрешения: новый метод определения электрической активности мозга. Внутр. J. Psychophysiol. 18, 49–65. DOI: 10.1016 / 0167-8760 (84)
М. и Леманн Д. (1994). Электромагнитная томография низкого разрешения: новый метод определения электрической активности мозга. Внутр. J. Psychophysiol. 18, 49–65. DOI: 10.1016 / 0167-8760 (84)
-x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Паттерсон К., Нестор П. Дж. И Роджерс Т. Т. (2007). Откуда ты знаешь то, что знаешь? Представление семантических знаний в человеческом мозге. Nat. Rev. Neurosci. 8, 976–987. DOI: 10.1038 / номер 2277
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Паттерсон, Р. Д., Уппенкамп, С., Джонсруд, И. С., и Гриффитс, Т. Д. (2002). Обработка временной высоты тона и мелодии в слуховой коре. Нейрон 36, 767–776.
Google Scholar
Прайс, К. Дж. (2012). Обзор и синтез первых 20 лет исследований прослушанной речи, разговорной речи и чтения с помощью ПЭТ и фМРТ. Neuroimage 62, 816–847.DOI: 10.1016 / j.neuroimage.2012.04.062
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Райхл, М. Э., МакЛауд, А. М., Снайдер, А. З., Пауэрс, В. Дж., Гуснард, Д. А., и Шульман, Г. Л. (2001). Режим работы мозга по умолчанию. Proc. Natl. Акад. Sci. 98, 676–682.
Э., МакЛауд, А. М., Снайдер, А. З., Пауэрс, В. Дж., Гуснард, Д. А., и Шульман, Г. Л. (2001). Режим работы мозга по умолчанию. Proc. Natl. Акад. Sci. 98, 676–682.
Google Scholar
Рамон, К., Шимпф, П., Хауэйзен, Дж., Холмс, М., Ишимару, А. (2004). Роль мягкой кости, спинномозговой жидкости и серого вещества в моделировании ЭЭГ. Brain Topogr. 16, 245–248. DOI: 10.1023 / B: брат.0000032859.68959.76
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Роджерс, Б. П., Морган, В. Л., Ньютон, А. Т., и Гор, Дж. К. (2007). Оценка функциональной связи в человеческом мозге с помощью фМРТ. Magn. Резон. Imaging 25, 1347–1357. DOI: 10.1016 / j.mri.2007.03.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Романски, Л. М., Тиан, Б., Фриц, Дж. Б., Мишкин, М., Гольдман-Ракич, П.С., и Раушекер, Дж. П. (2000). Ответьте на вопросы «Что?», «Где» и «Как в слуховой коре». Nat. Neurosci. 3: 966. DOI: 10. 1038 / 79892
1038 / 79892
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Стаерен Н., Ренвалл Х., Де Мартино Ф., Гебель Р. и Формизано Э. (2009). Звуковые категории представлены в виде распределенных паттернов в слуховой коре человека. Curr. Биол. 19, 498–502. DOI: 10.1016 / j.cub.2009.01.066
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Steinke, W.Р., Кадди, Л. Л., и Якобсон, Л. С. (2001). Диссоциация функциональных подсистем, управляющих распознаванием мелодии после повреждения правого полушария. Cogn. Neuropsychol. 18, 411–437. DOI: 10.1080 / 026432702
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Таниваки Т., Тагава К., Сато Ф. и Иино К. (2000). Слуховая агнозия ограничивается звуками окружающей среды после корковой глухоты и генерализованной слуховой агнозии. Clin. Neurol. Нейрохирургия. 102, 156–162. DOI: 10.1016 / s0303-8467 (00) 00090-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Терваниеми, М. , и Хугдал, К. (2003). Латерализация слуховой коры. Brain Res. Ред. 43, 231–246. DOI: 10.1016 / j.brainresrev.2003.08.004
, и Хугдал, К. (2003). Латерализация слуховой коры. Brain Res. Ред. 43, 231–246. DOI: 10.1016 / j.brainresrev.2003.08.004
CrossRef Полный текст | Google Scholar
Туник Э., Ло О. Ю., Адамович С. В. (2008). Транскраниальная магнитная стимуляция лобной крышки и надмаргинальной извилины нарушает планирование основанных на результате взаимодействий руки и объекта. J. Neurosci. 28, 14422–14427. DOI: 10.1523 / jneurosci.4734-08.2008
CrossRef Полный текст | Google Scholar
Вендел, К., Нарра, Н. Г., Ханнула, М., Кауппинен, П., и Мальмивуо, Дж. (2008). Влияние ЦСЖ на распределение чувствительности ЭЭГ многослойных моделей головы. IEEE Trans. Биомед. Англ. 55, 1454–1456. DOI: 10.1109 / TBME.2007.7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вольтерс, К. Х., Анвандер, А., Трикош, X., Вайнштейн, Д., Кох, М.А., и Маклеод, Р.С. (2006). Влияние анизотропии проводимости ткани на поле ЭЭГ / МЭГ и расчет обратного тока в реалистичной модели головы: моделирование и визуализация с использованием конечно-элементного моделирования с высоким разрешением. NeuroImage 30, 813–826. DOI: 10.1016 / j.neuroimage.2005.10.014
NeuroImage 30, 813–826. DOI: 10.1016 / j.neuroimage.2005.10.014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Залески, А., Форнито, А., Буллмор, Э. Т. (2010). Сетевая статистика: выявление различий в сетях мозга. Neuroimage 53, 1197–1207. DOI: 10.1016 / j.neuroimage.2010.06.041
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Заторре, Р. Дж., Буффар, М., и Белин, П. (2004). Чувствительность к свойствам слуховых объектов в височной неокортексе человека. J. Neurosci. 24, 3637–3642. DOI: 10.1523 / jneurosci.5458-03.2004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цзэн, К. (2015). Генерация ключей физического уровня в беспроводных сетях: проблемы и возможности. IEEE Commun. Mag. 53, 33–39. DOI: 10.1109 / mcom.2015.7120014
CrossRef Полный текст | Google Scholar
Поляризация: ключевое различие между искусственными и естественными электромагнитными полями с точки зрения биологической активности
Все критические биомолекулы либо электрически заряжены, либо полярны 11 . В то время как естественные неполяризованные ЭМП / ЭМИ любой интенсивности не могут вызвать какие-либо специфические / когерентные колебания на этих молекулах, поляризованные искусственные ЭМП / ЭМИ будут вызывать когерентные вынужденные колебания на каждой заряженной / полярной молекуле в биологической ткани.Это фундаментально для нашего понимания биологических явлений. Эти колебания будут наиболее очевидны для свободных (подвижных) ионов, которые несут чистый электрический заряд и существуют в больших концентрациях во всех типах клеток или внеклеточной ткани, определяя практически все клеточные / биологические функции 11 . Хотя все молекулы колеблются случайным образом с гораздо более высокими скоростями из-за теплового движения, это не имеет никакого биологического эффекта, кроме повышения температуры ткани. Но когерентные поляризованные колебания энергии даже в миллионы раз меньшей, чем средняя тепловая молекулярная энергия 26 , могут вызвать биологические эффекты.
В то время как естественные неполяризованные ЭМП / ЭМИ любой интенсивности не могут вызвать какие-либо специфические / когерентные колебания на этих молекулах, поляризованные искусственные ЭМП / ЭМИ будут вызывать когерентные вынужденные колебания на каждой заряженной / полярной молекуле в биологической ткани.Это фундаментально для нашего понимания биологических явлений. Эти колебания будут наиболее очевидны для свободных (подвижных) ионов, которые несут чистый электрический заряд и существуют в больших концентрациях во всех типах клеток или внеклеточной ткани, определяя практически все клеточные / биологические функции 11 . Хотя все молекулы колеблются случайным образом с гораздо более высокими скоростями из-за теплового движения, это не имеет никакого биологического эффекта, кроме повышения температуры ткани. Но когерентные поляризованные колебания энергии даже в миллионы раз меньшей, чем средняя тепловая молекулярная энергия 26 , могут вызвать биологические эффекты.
Вынужденные колебания подвижных ионов, вызванные внешней поляризованной ЭДС, могут привести к нерегулярному закрытию каналов электроприводных ионов на клеточных мембранах. Это было подробно описано в Panagopoulos et al . 19,20 . Согласно этой теории — правдоподобие которой в реальных биологических условиях было подтверждено численным тестом 27 — вынужденные колебания ионов в непосредственной близости от датчиков напряжения потенциалозависимых ионных каналов могут оказывать на эти датчики силы, равные или больше, чем известные физиологические силы, закрывающие эти каналы.Нерегулярное закрытие этих каналов может потенциально нарушить электрохимический баланс и функцию любой клетки 11 , что приведет к различным биологическим последствиям / последствиям для здоровья, включая наиболее пагубные, такие как повреждение ДНК, гибель клеток или рак 28 .
Большинство катионных каналов (Ca +2 , K + , Na + и т. Д.) На мембранах всех клеток животных управляются напряжением 11 . Они взаимно преобразуются между открытым и закрытым состояниями, когда электростатическая сила, действующая на электрические заряды их датчиков напряжения из-за трансмембранного изменения напряжения, превышает некоторое критическое значение.Датчики напряжения этих каналов представляют собой четыре симметрично расположенных трансмембранных положительно заряженных спиральных домена, каждый из которых обозначен как S4. Изменения трансмембранного потенциала порядка 30 мВ обычно требуются для закрытия электропроводных каналов 29,30 . Несколько ионов могут одновременно взаимодействовать с доменом S4 на расстоянии порядка 1 нм, поскольку — за исключением одного иона, который может проходить через поры канала, когда канал открыт — еще несколько ионов связаны близко к поры канала в определенных сайтах связывания ионов (например,грамм. три в калиевых каналах) 31 . Подробную информацию о структуре и функции катионных электросенсорных каналов можно найти в 11,29,31 .
Д.) На мембранах всех клеток животных управляются напряжением 11 . Они взаимно преобразуются между открытым и закрытым состояниями, когда электростатическая сила, действующая на электрические заряды их датчиков напряжения из-за трансмембранного изменения напряжения, превышает некоторое критическое значение.Датчики напряжения этих каналов представляют собой четыре симметрично расположенных трансмембранных положительно заряженных спиральных домена, каждый из которых обозначен как S4. Изменения трансмембранного потенциала порядка 30 мВ обычно требуются для закрытия электропроводных каналов 29,30 . Несколько ионов могут одновременно взаимодействовать с доменом S4 на расстоянии порядка 1 нм, поскольку — за исключением одного иона, который может проходить через поры канала, когда канал открыт — еще несколько ионов связаны близко к поры канала в определенных сайтах связывания ионов (например,грамм. три в калиевых каналах) 31 . Подробную информацию о структуре и функции катионных электросенсорных каналов можно найти в 11,29,31 .
Рассмотрим, например, четыре иона калия на расстоянии порядка 1 нм от каналов-датчиков (S4) и приложенная извне осциллирующая ЭДС / ЭМИ. Электрическая (и магнитная) сила, действующая на каждый ион из-за любого неполяризованного поля, равна нулю (уравнение 8). Напротив, сила, обусловленная поляризованным полем с электрическим компонентом E , составляет F = Ezq e .Для синусоидального переменного поля Ε = Ε 0 sin ωt уравнение движения свободного иона с массой м i , составляет 19,20 :
, где r — ион смещение из-за вынужденных колебаний, z — валентность иона ( z = 1 для ионов калия), q e = 1,6 × 10 −19 C — элементарный заряд, λ — затухание коэффициент для смещения иона (рассчитанный как имеющий значение внутри канала), ω 0 = 2πν 0 (ν 0 собственная частота колебаний иона, принятая равной зарегистрированной частоте спонтанных внутриклеточных колебаний иона порядка 0.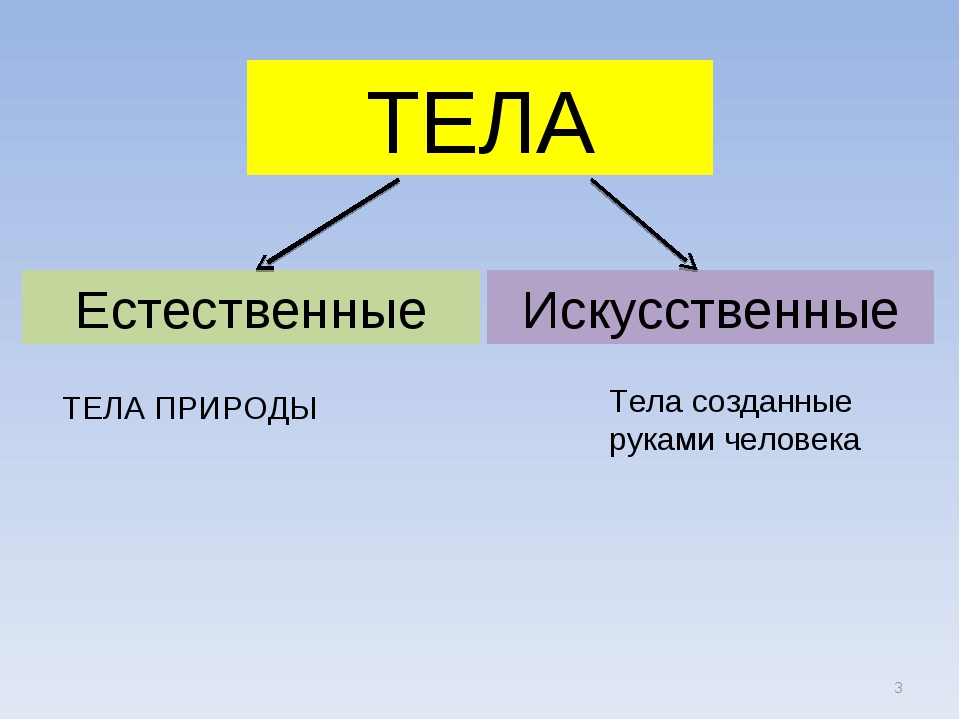 1 Гц), ω = 2πν (ν частота поля / излучения) и E 0 амплитуда поля 19,20 .
1 Гц), ω = 2πν (ν частота поля / излучения) и E 0 амплитуда поля 19,20 .
Общее решение уравнения. 22, это 19,20 :
Член в решении представляет постоянное смещение, но не влияет на колебательный член. Это постоянное смещение удваивает амплитуду вынужденных колебаний в тот момент, когда поле прикладывается или прерывается, или во время его первого и последнего периодов, и смещение иона будет вдвое больше амплитуды вынужденных колебаний.Для импульсных полей (таких как большинство областей современной цифровой связи) это будет происходить постоянно с каждым повторяющимся импульсом. Таким образом, импульсные поля — теоретически — в два раза более сильные, чем непрерывные / непрерывные поля с теми же другими параметрами, что согласуется с несколькими экспериментальными данными 1,32 .
Амплитуда вынужденных колебаний (без учета постоянного члена в уравнении 23) составляет:
Сила, действующая на эффективный заряд q домена S4 через колеблющийся одновалентно-свободный катион, составляет: , ( r — расстояние свободного иона от эффективного заряда S4).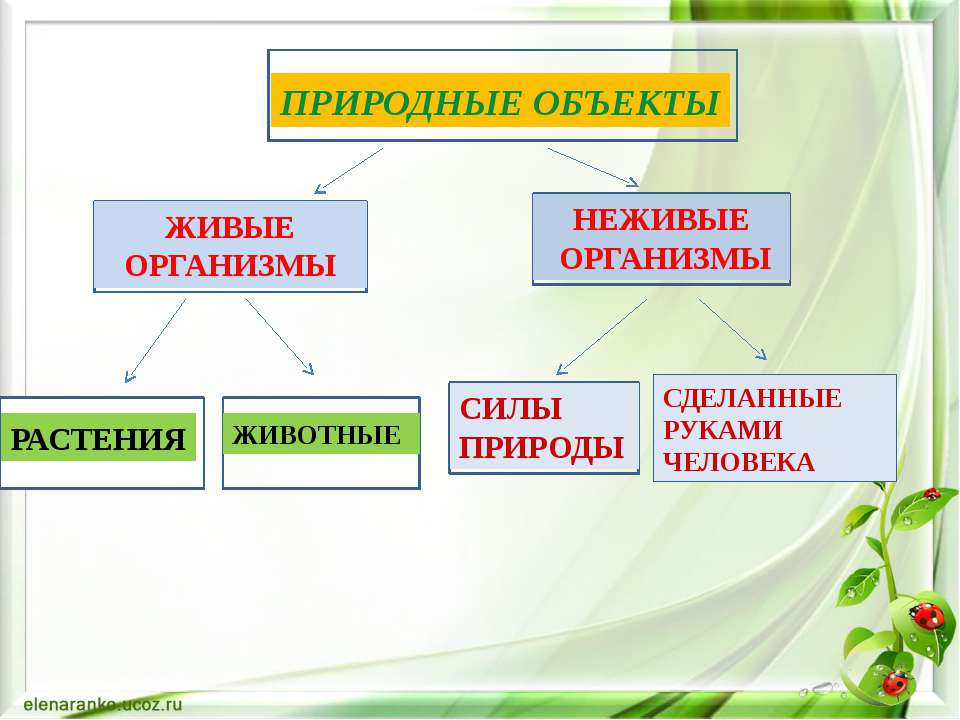 Каждый колебательный катион, смещенный на dr , индуцирует силу на каждом датчике S4:
Каждый колебательный катион, смещенный на dr , индуцирует силу на каждом датчике S4:
В то время как в случае неполяризованного приложенного поля и, в случае поляризованного приложенного поля, суммарная сила на датчик канала из всех четыре катиона, составляет:
Это даже более важное различие между поляризованными и неполяризованными ЭМП в отношении биологической активности, чем способность интерференции.
Эффективная плата за каждый домен S4 составляет: q = 1.7 q e 30 . Минимальная сила, действующая на этот заряд, обычно необходимая для закрытия канала, равная силе, создаваемой изменением на 30 мВ мембранного потенциала 30 , вычисляется 19 и составляет:
Смещение одного одновалентного катиона внутри канала, необходимое для приложения этой минимальной силы рассчитывается по формуле. 25 должно быть:
Для 4 катионов, колеблющихся в фазе и в параллельных плоскостях из-за внешнего поляризованного поля / излучения, минимальное смещение уменьшается до: dr = 10 −12 м.
Следовательно, любая внешняя поляризованная осциллирующая ЭДС, способная заставить свободные ионы колебаться с амплитудой, может беспорядочно закрывать катионные каналы на клеточных мембранах. Для z = 1 (ионы калия) и подставляя значения для q e , λ в последнем условии, получаем:
(ν в Гц, 0 в В / м)
Для двухвалентных катионов ( z = 2) (например, Ca +2 ) условие принимает следующий вид:
(ν в Гц, Ε 0 в В / м)
[Подробное описание Кратко представленный механизм можно найти в 19,20 .]
Для электрических полей (ν = 50 Гц) Условие 27 становится,
Таким образом, ЭДС промышленной частоты с интенсивностью, превышающей 5 мВ / м, потенциально способны нарушить функцию клетки. Для количества источников ЭМП N одинаковой поляризации (например, N количества параллельных линий электропередачи) последнее значение делится на N (согласно уравнению 19) в местах конструктивных помех и, следовательно, даже больше. уменьшилось. Такие минимальные значения напряженности поля промышленной частоты распространены в повседневной городской среде и даже ближе к высоковольтным линиям электропередачи 7 .
уменьшилось. Такие минимальные значения напряженности поля промышленной частоты распространены в повседневной городской среде и даже ближе к высоковольтным линиям электропередачи 7 .
Для импульсных полей вторая часть условия 27 делится на 2 и становится:
(ν в Гц, Ε 0 в В / м).
Для полей / излучения цифровой мобильной телефонии, излучающих КНЧ-импульсы с частотой повторения импульсов ν = 217 Гц (среди других частот КНЧ они передают) 33 , Условие 29 становится:
Для частоты повторения импульсов ν = 8,34 Гц ( также включается в сигналы мобильной телефонии) 33,34 , Условие 29 становится:
Как видно из описанного механизма, поле не закрывает канал силами, действующими непосредственно на датчики канала.Для этого потребуется поле порядка трансмембранного поля (10 6 –10 7 В / м). Именно посредничество колеблющихся свободных ионов в непосредственной близости к датчикам канала S4 позволяет таким слабым полям создавать необходимые силы для закрытия канала.
Таким образом, электрические поля СНЧ, излучаемые мобильными телефонами и базовыми станциями сильнее 0,0004 В / м, также потенциально способны нарушить работу любой живой клетки. Это значение интенсивности ELF излучается обычными сотовыми телефонами на расстоянии до нескольких метров и базовыми станциями на расстоянии до нескольких сотен метров 6,34,35 .Для числа вертикально ориентированных антенн мобильной телефонной связи N последнее значение делится на N (согласно уравнению 19) в местах возникновения конструктивных помех.
Мы не делаем различий между ЭМП, прикладываемой извне, и ЭМП, индуцированной внутри живой ткани, особенно в случае СНЧ по следующим причинам: 1. Живая ткань не является металлом для защиты от электрических полей и, конечно же, не является ферромагнитным металлом (Fe, Co, Ni) для защиты от магнитных полей.Более того, известно, что особенно поля КНЧ не могут быть легко экранированы даже клетками Фарадея, и для того, чтобы значительно их минимизировать, рекомендуется полностью заключать их в закрытые металлические коробки 6 . Таким образом, электрические поля СНЧ проникают в живую ткань с определенной степенью затухания, а магнитные поля проникают с нулевым затуханием. 2. Даже в том случае, если поля СНЧ значительно ослаблены во внутренних тканях живого тела, глаза, мозг, клетки кожи или мириады окончаний нервных волокон, которые попадают на внешний эпидермис, подвергаются прямому воздействию интенсивности поля, измеренные снаружи на поверхности живой ткани.
Таким образом, электрические поля СНЧ проникают в живую ткань с определенной степенью затухания, а магнитные поля проникают с нулевым затуханием. 2. Даже в том случае, если поля СНЧ значительно ослаблены во внутренних тканях живого тела, глаза, мозг, клетки кожи или мириады окончаний нервных волокон, которые попадают на внешний эпидермис, подвергаются прямому воздействию интенсивности поля, измеренные снаружи на поверхности живой ткани.
Было показано, что препараты ткани (такие как фибробласты крупного рогатого скота или сухожилия курицы) реагируют на приложенные извне импульсные или синусоидальные электрические поля СНЧ (изменениями скорости синтеза ДНК или белка, скорости пролиферации, выравнивания по направлению поля и т. Д. ), при очень низких порогах ~ 10 −3 В / м 1,36,37,38 . Эти пороговые значения очень близки к прогнозируемым в настоящем исследовании.
За исключением прямого воздействия электрического поля внешним полем, внутри тканей может быть электрическое поле, индуцированное приложенным извне осциллирующим магнитным полем, которое, как уже объяснялось, проникает в живую ткань с нулевым затуханием. Туор и др. . 34 измерили магнитные поля СНЧ от сотовых телефонов порядка 1 Гс (= 10 −4 Тл) при 217 Гц. Это может индуцировать электрические поля порядка ~ 0,1 В / м в человеческом теле, как может быть показано применением закона электромагнитной индукции Максвелла:
Туор и др. . 34 измерили магнитные поля СНЧ от сотовых телефонов порядка 1 Гс (= 10 −4 Тл) при 217 Гц. Это может индуцировать электрические поля порядка ~ 0,1 В / м в человеческом теле, как может быть показано применением закона электромагнитной индукции Максвелла:
(,,, напряженности магнитного и индуцированного электрического поля соответственно, приращение длины по замкнутому пути – циркуляции индуцированного электрического поля, охватывающего поверхность S.- единичный вектор, вертикальный к поверхности S ).
Предполагая, что параллельно и независимо от l , вертикально относительно S и l и независимо от них, круговой путь радиуса α, включая поверхность S , уравнение. 32 становится:
, что дает:
( E ind в В / м, B в T, α в м).
Путем замены в последнем уравнении α = 0,20 м (достаточно большой радиус для окружности тела взрослого человека) и [согласно Туору и др. . 34 ], получаем E ind ~ 0,1 В / м. Это напряженность электрического поля, наведенного в человеческом теле импульсами мобильной телефонной связи с частотой 217 Гц, и она примерно в десять раз больше минимального расчетного значения, способного вызвать биологические эффекты на этой частоте в соответствии с Условием 30.
. 34 ], получаем E ind ~ 0,1 В / м. Это напряженность электрического поля, наведенного в человеческом теле импульсами мобильной телефонной связи с частотой 217 Гц, и она примерно в десять раз больше минимального расчетного значения, способного вызвать биологические эффекты на этой частоте в соответствии с Условием 30.
Искусственный материал сейчас перевешивает все остальные живые существа на Земле
С тех пор, как началась промышленная революция, мы создали множество вещей.Скорее всего, это всего лишь часть человечества: от дорог и домов до всевозможных транспортных средств, нам нужны вещи. Но, возможно, мы здесь немного переборщили.
Согласно новому исследованию, созданные людьми объекты теперь весят примерно столько же, сколько все живые существа на Земле, а в этом году могут даже превзойти их.
Изображение предоставлено: Flickr / Alexandre Macedo Человечество постепенно стало доминирующей силой в формировании лица Земли. Его воздействие было настолько резким и значительным, что было предложено переименовать нынешнюю геологическую эпоху, голоцен, в антропоцен.Слово происходит от греческих слов Anthropos , означающих «человек», и cene, означающих «новый» — так что мы своими действиями в основном открыли новую геологическую эру.
Его воздействие было настолько резким и значительным, что было предложено переименовать нынешнюю геологическую эпоху, голоцен, в антропоцен.Слово происходит от греческих слов Anthropos , означающих «человек», и cene, означающих «новый» — так что мы своими действиями в основном открыли новую геологическую эру.
Это заставило исследователей из израильского Института науки Вейцмана задаться вопросом, сколько материальных продуктов производит деятельность человека и как это соотносится с общей природной биомассой. Они уже опубликовали оценку количества биомассы на Земле и теперь решили сделать еще один шаг, изучив искусственные объекты и сравнив их с естественной массой.
«Это исследование дает символическую и основанную на массе количественную характеристику антропоцена», — рассказали Scientific American два автора исследования, Эмили Эльхачам и Рон Майло. «Учитывая эмпирические данные о накопленной массе человеческих артефактов, мы больше не можем отрицать нашу центральную роль в мире природы».

Для исследования исследователи разделили все созданные человеком объекты на шесть основных категорий: бетон, заполнители (включая такие материалы, как гравий), кирпичи, асфальт, металлы и «другие» материалы, включая пластик, дерево, используемое для строительства, бумага и стекло.Они не учитывали отходы в своих расчетах, а это означает, что общая масса, созданная руками человека, уже даже превышает расчет исследователей.
Они обнаружили, что все искусственные объекты весят приблизительно 1,1 триллиона метрических тонн. Это равняется суммарному сухому весу всех растений, животных, грибов, бактерий, архей и простейших на планете.
В этой сравнительной шкале человечество работает в двух направлениях: это не только строительство искусственных объектов, но и разрушение природных ландшафтов.Со времени первой сельскохозяйственной революции человечество примерно вдвое уменьшило массу растений, примерно с двух тератонов до нынешней стоимости примерно 1,1 тератона. Общая масса одомашненных культур значительно перевешивается потерей массы растений в результате вырубки лесов и других изменений в землепользовании — очевидно, здоровый лес с высокими деревьями значительно превышает размер, покрытый пшеницей или другими сельскохозяйственными растениями.
Общая масса одомашненных культур значительно перевешивается потерей массы растений в результате вырубки лесов и других изменений в землепользовании — очевидно, здоровый лес с высокими деревьями значительно превышает размер, покрытый пшеницей или другими сельскохозяйственными растениями.
Это тоже не конец. По оценкам исследователей, 2020 год может быть годом, когда масса, созданная человеком, превзойдет общий вес биомассы.Искусственная масса производится со скоростью 30 гигатонн в год. Это означает, что еженедельно производятся материалы, превышающие вес каждого человека на планете.
«Воздействие человеческой деятельности было настолько резким и значительным, что было предложено переименовать нынешнюю геологическую эпоху в антропоцен. Наше исследование строго и количественно подтверждает это предположение. Параллельно это добавляет еще одно измерение к этой дискуссии — символическое количественное разграничение перехода к нашей эпохе », — пишут исследователи.

Несмотря на то, что ученые описывают находку как символическую, они утверждают, что верстовой столб дает человечеству возможность оценить, как мы сюда попали и как выглядит будущее. Это мощный сигнал для всех, который привлекает внимание к тому, как современные общества могут устойчиво развиваться в ближайшем будущем.
Исследование опубликовано в журнале Nature.
Контраст между искусством и природой часто изображается в искусстве, но не так очевидно, как птицы Сары Ландета, нарисованные на коробках с наркотиками.| by Galerie Project
Контраст между созданием человека и природой часто изображается в произведениях искусства, но, возможно, не так очевиден, как птицы Сары Ландета, нарисованные на коробках с наркотиками.
Хотя Ландета в прошлом создала множество изделий этой серии, она только что выпустила новую партию, которая включает 120 новых дизайнов.
Коробки были собраны у пациентов, которые использовали лекарства для лечения различных заболеваний — то, что Ландета стремится показать в своей работе.Каждая коробка была тщательно декорирована птицами и природными пейзажами с деревьями, растениями и яркими цветами.
Почему птицы? Потому что, по словам Ландеты, они — единственное животное, придающее смысл свободе. Птицы — единственные существа на планете, которые могут соединяться с землей и небом, но они также являются одним из основных видов животных, содержащихся в неволе. Это добавляет интересный поворот к понятию свободы, которое Ландета использует в своих проектах.
Именно тогда, когда эти изображения свободы или различных видов птиц, которые использует Ландета, сочетаются с синтетической идеей наркотиков, эти части действительно начинают обретать смысл.«Это сопоставление естественного и синтетического интерпретирует пациента как содержащееся в неволе животное», — говорит Ландета, добавляя, что птица является метафорой этой теории.
Landeta передает идею о том, что пациент становится пойманной птицей. «Они хотят летать», — говорит она о пациентах, добавляя, что это лекарства, которые поддерживают их жизнь; которые помогают им преодолеть свои болезни и снова стать свободными.
Однако источником вдохновения для создания этих произведений является не только идея свободы.На самом деле, Ландета многим обязана Джону Джеймсу Одубону, художнику 19-го века, который вдохновил эту серию.
Использование коробок с лекарствами в качестве холста позволяет Ландете резко показать контраст между искусственным миром и природным миром, а также его неразрывную связь. Она глубоко копает в своей работе, чтобы коснуться споров о внешнем и внутреннем, и активно использует идею о том, что для того, чтобы человечество могло выжить в течение любого периода времени, мы должны полагаться на природу, а не на искусственные фармацевтические препараты, в которых так много людей нуждаются.
Собрав коробки (все они были использованы и в прошлой жизни видели в руках своих пациентов), Ландета тщательно создает на них естественные сцены, когда они открываются для создания холста.