Примавера-В какую фазу просыпаться?
Фазы сна и когда лучше просыпаться, чтобы быть бодрым?
Давайте рассмотрим, все этапы сна и выберем более подходящий для пробуждения!
Рассматривая отдых, можно условно разделить весь период на 4 фазы длинного и 1 фазу короткого сна.
Что происходит на каждом отрезке сна:
1. Медленный сон:
Включает 4 периода, считается самым полноценным для отдыха, перезагрузки систем организма:
- Дремота – I фаза медленного сна длится 5-20 минут, момент пограничного состояния человека, когда он поддается самовнушению.
В дремоте мозг «завершает» обработку информации, которой был занят.
- Длительные II, III фазы медленного сна:
Занимают половину времени отдыха, характеризуются глубоким расслаблением мышц, систем жизнедеятельности. Человек легко пробуждается, но чувствует себя не совсем бодрым.
- Глубокий дельта-сон IV фаза:
В этом состоянии у вас понижается температура тела, замедляется ритм сердца, мышцы окончательно расслабляются, вырабатывается гормон роста.
2. Быстрый сон:
Отдельная фаза, характерная повышением деятельности всех органов, систем.
Мозг активен, пробуждение тяжелое, но следует вставать именно в этой фазе, поэтому период еще называется парадоксальным. Сновидение в этой фазе яркое, запоминающееся.
Почему вставать нужно именно в быструю фазу?
Из-за активизации мозговой деятельности. Состояние сонливости быстро проходит, смена режима не вызывает отторжения, организм готов работать, думать.
Внимание! Для человека, пробуждаемого в фазе быстрого сна, не имеет значения час вставания. Вне зависимости от того, день или ночь на улице, вы будете работать одинаково эффективно и с полноценной отдачей.
Основные предположения сводятся к мозговой деятельности:
Мозг систематизирует, упорядочивает информацию, вырабатывает стратегические приемы адаптации к задачам, их решению.
Быстрый сон помогает сохранить эмоциональное, психологическое спокойствие!
Спите правильно! Подглядывайте в наши лайфхаки.
Фазы сна (медленный, быстрый сон) и сновидения | Биология. Реферат, доклад, сообщение, краткое содержание, лекция, шпаргалка, конспект, ГДЗ, тест
Фаза медленного сна — глубокий без сновидений сон, что составляет 75—80 % от всей его длительности. В это время замедляется сердечная деятельность, интенсивность обмена веществ и процессов газообмена, снижается температура тела, расслабляются скелетные мышцы, почти исчезают все виды чувствительности (тактильная, болевая, слуховая и т. п.). Мышление прекращается (рис. 209).
Быстрый сон составляет 20—25 % всего сна. Во время этой фазы, по сравнению с фазой медленного сна, немного усиливается сердечная деятельность и дыхание, менее расслабленными становятся скелетные мышцы, несколько повышаются чувствительность, обмен веществ и температура тела. Однако растёт и торможение определённых участков коры головного мозга, поэтому человека тяжелее разбудить. Он начинает видеть сны. Глазные яблоки при этом быстро двигаются, как будто человек пересматривает кинофильм (рис. 210).
Во время этой фазы, по сравнению с фазой медленного сна, немного усиливается сердечная деятельность и дыхание, менее расслабленными становятся скелетные мышцы, несколько повышаются чувствительность, обмен веществ и температура тела. Однако растёт и торможение определённых участков коры головного мозга, поэтому человека тяжелее разбудить. Он начинает видеть сны. Глазные яблоки при этом быстро двигаются, как будто человек пересматривает кинофильм (рис. 210).
Медленная и быстрая фазы сна сменяют друг друга несколько раз с промежутками 80-120 мин; за ночь наблюдается 4-6 таких циклов. Обычно с такими же промежутками возникают сновидения (рис. 211).
| Рис. 209. Медленный сон |
Сновидения — образные представления, нередко очень яркие, фантастические, эмоционально окрашенные, возникающие во время быстрого сна.
| Рис. 210. быстрый сон |
Сновидения бывают из-за того, что разные участки коры головного мозга тормозятся неодинаково. Некоторые из них — перевозбуждённые (вспомним доминирующие центры коры головного мозга) — могут почти не тормозиться и подчинять себе все другие мозговые центры. Во время сновидений могут активизироваться следы долговременной памяти, связанные даже со случайными впечатлениями далёкого прошлого. Именно поэтому во сне человек может решить сложную задачу или вспомнить местонахождение нужной вещи, о чём давно забыл.
Характер снов зависит от степени эмоциональности человека, а также от того, какое мышление — образное или абстрактное — у него преобладает. Из-за торможения определённых зон коры головного мозга другие зоны ночью могут стать чувствительнее, чем днём. Например, человеку, который хорошо себя чувствовал, ночью может присниться сон о какой-то болезни. Это явление вызывают слабые импульсы больного органа, которые днём заглушены множеством более сильных. Во время сна они свободно, без препятствий попадают в определённые центры головного мозга. На такие сны необходимо обращать внимание. Сновидения бывают компактными во времени: за считанные секунды человеку может присниться значительный промежуток жизни со множеством ярких событий. Материал с сайта http://worldofschool.ru
Это явление вызывают слабые импульсы больного органа, которые днём заглушены множеством более сильных. Во время сна они свободно, без препятствий попадают в определённые центры головного мозга. На такие сны необходимо обращать внимание. Сновидения бывают компактными во времени: за считанные секунды человеку может присниться значительный промежуток жизни со множеством ярких событий. Материал с сайта http://worldofschool.ru
| Рис. 211. Фазы ночного сна человека |
Сон, развивающийся у человека и животных под действием тормозных условных раздражителей, И. Павлов называл активным, противопоставляя ему пассивный сон, возникающий в случае прекращения или резкого ограничения притока сигналов от органов чувств в кору больших полушарий головного мозга. Наблюдали больного, у которого из всех органов чувств сохранились функции одного уха и одного глаза. Пока глаз мог видеть, а ухо — слышать, человек не спал, но как только врачи закрывали ему эти единственные пути общения с внешним миром, он сразу засыпал.
Пока глаз мог видеть, а ухо — слышать, человек не спал, но как только врачи закрывали ему эти единственные пути общения с внешним миром, он сразу засыпал.
Фазы сна человека и работа органов чувств человека
Фазы сна и сновидения
Сны и сновидения краткое сообщение
Как сон полезный медленный быстрый
Фазы сна чувствительность тела остается
Докажите, что работа мозга во время сна не прекращается, а по некоторым показателям — даже активизируется.
Объясните физиологические механизмы сновидений и докажите, что во снах нет ничего «таинственного».
Алкоголь в качестве снотворного приводит к нарушениям сна
Погружение в сон с помощью алкоголя не принесет ни отдыха, ни пользы здоровью, установили ученые из Великобритании, труд которых запланирован к публикации в журнале Alcoholism: Clinical & Experimental Research в апреле текущего года.
Исследователи из Лондонского и Эдинбургского центров сна изучили всю доступную литературу за последние 30 лет о влиянии алкоголя на характер сна человека. В итоге они выяснили: кажущаяся польза алкоголя в качестве снотворного, позволяющего легко и быстро заснуть, оборачивается нарушениями сна, храпом и затруднениями с дыханием.
«Наш обзор подтверждает, что алкоголь сокращает время засыпания, а в больших дозах увеличивает влияние на фазу глубокого сна. Этот эффект первой половины сна отчасти является резоном для некоторых людей, страдающих бессонницей, использовать алкоголь в качестве средства от нее», — цитируются в совместном пресс-релизе двух институтов слова директора Лондонского центра сна (London Sleep Centre) Иршаада Эбрахима (Irshaad Ebrahim).
Однако, отмечает исследователь, эффект более глубокого сна в первой половине ночи будет компенсирован нарушениями сна в оставшиеся до утра часы.
Авторы напоминают, что сон представлен двумя стадиями активности головного мозга, которые называются стадиями быстрого и медленного сна, перемежающимися друг с другом. Алкоголь увеличивает фазу медленного, глубокого сна, но укорачивает фазу быстрого сна, делая его более тревожным.
Алкоголь увеличивает фазу медленного, глубокого сна, но укорачивает фазу быстрого сна, делая его более тревожным.
«Чаще всего в быстрой фазе нам снятся сны. В это время наш мозг более активен, можно сравнить этот период с процессом дефрагментации жестких дисков в компьютере. Фаза быстрого сна также важна, потому что влияет на нашу память и укрепляет ее», — говорит Эбрахим.
Он отмечает, что недостаток этой фазы, составляющей в целом 20-25% от всего сна, может отрицательно влиять на память, способность концентрации и моторные функции.
Авторы также говорят, что сон, спровоцированный алкоголем, не принесет отдыха, и может быть связан с депрессией. «Не стоит рассчитывать, что ваш сон станет лучше с алкоголем», — предупреждают они.
Обзор, стадия I сна, стадия II сна
Автор
Селим Бенбадис, доктор медицины Профессор, директор Комплексной программы эпилепсии, отделения неврологии и нейрохирургии, Общая больница Тампа, Медицинский колледж Морсани Университета Южной Флориды
Селим Бенбадис, доктор медицины, является членом следующих медицинских обществ: Американская академия неврологии, Американская академия медицины сна, Американское общество клинической нейрофизиологии, Американское общество эпилепсии, Американская медицинская ассоциация
Раскрытие информации: Служить (d) в качестве директора, должностного лица, партнера, сотрудника, советника, консультанта или попечителя для: Альянса, Bioserenity, Ceribell, Eisai, Greenwich, LivaNova, Neurelis, Neuropace, Nexus, RSC, SK life science, Sunovion
Получил исследовательский грант от: Cerevel, LivaNova, Greenwich, SK biopharmaceuticals, Takeda.

Соавтор (ы)
Диего Антонио Риело, MD Штатный врач отделения неврологии, Memorial Hospital West, Memorial Healthcare
Диего Антонио Риело, доктор медицины, является членом следующих медицинских обществ: Американская академия неврологии
Раскрытие информации: не раскрывать.
Специальная редакционная коллегия
Франсиско Талавера, фармацевт, доктор философии Адъюнкт-профессор, Фармацевтический колледж Медицинского центра Университета Небраски; Главный редактор Medscape Drug Reference
Раскрытие информации: Получил зарплату от Medscape за работу.для: Medscape.
Норберто Альварес, доктор медицины Доцент кафедры неврологии Гарвардской медицинской школы; Консультант отделения неврологии детской больницы Бостона; Медицинский директор Центра развития Рентама
Норберто Альварес, доктор медицины, является членом следующих медицинских обществ: Американская академия неврологии, Американское общество эпилепсии, Общество детской неврологии
Раскрытие информации: не подлежит разглашению.
Главный редактор
Helmi L Lutsep, доктор медицины Профессор и заместитель председателя кафедры неврологии Медицинского факультета Орегонского университета здоровья и науки; Заместитель директора Центра инсульта OHSU
Хелми Л. Луцеп, доктор медицины, является членом следующих медицинских обществ: Американская академия неврологии, Американская ассоциация инсульта
Раскрытие информации: Medscape Neurology Редакционный консультативный совет: Комитет по рассмотрению инсульта, CREST2; Консультативный совет врачей Coherex Medical; Клиническое испытание национального лидера и руководящего комитета, Bristol Myers Squibb; Abbott Laboratories, консультативная группа.
Быстрый дельта-распад после продолжительного бодрствования отмечает фазу инерции бодрствования в NREM-сне
Эксперименты на мышах
Все мыши были самцами C57BL / 6J в возрасте 10–16 недель на момент операции. После операции мышей содержали индивидуально при постоянной температуре окружающей среды (25 ° C) и при цикле 12 часов света / 12 часов темноты (LD12: 12, флуоресцентные лампы, интенсивность 6,6 кд / м -2 , с временем Цайтгебера). (ZT) 0 и -12, обозначающие начало светлого и темного дня соответственно) и с пищей и водой, даваемыми ad libitum.Все описанные процедуры с животными были выполнены в соответствии с руководящими принципами швейцарского федерального законодательства и были предварительно одобрены ветеринарными службами кантонов Во и Берн.
(ZT) 0 и -12, обозначающие начало светлого и темного дня соответственно) и с пищей и водой, даваемыми ad libitum.Все описанные процедуры с животными были выполнены в соответствии с руководящими принципами швейцарского федерального законодательства и были предварительно одобрены ветеринарными службами кантонов Во и Берн.
ЭЭГ / ЭМГ / термисторная хирургия
Операции выполнялись в соответствии с ранее опубликованными протоколами под глубокой анестезией с использованием раствора кетамина / ксилазина [8% ксилазина (Rompun, 2%), 10% кетамина (Ketazol-100, Graeub) и 82% физиологический раствор (0,9% NaCl)] вводили перитонеально (1 мл кг -1 ) перед хирургическими вмешательствами.Температуру тела постоянно контролировали с помощью ректального термометра, и мышей помещали на регулируемую грелку (DC Temperature Control System, FHC). Вкратце, мышам имплантировали позолоченные винты из нержавеющей стали, которые были припаяны к медным проводам, прикрепленным к разъему. Электроды располагали вдоль изолатеральной (правое полушарие) оси на поверхности коры (Bregma <0,2 мм) во фронтальной (AP = +1,5 мм, ML = +1,5 мм), центральной (AP = -1,0 мм, ML = +2,5 мм) и теменной (затылочной; AP = −3.0 мм, ML = +1,5 мм), у шести мышей (рис. 4а, б). Всем другим мышам имплантировали лобно-теменные биполярные электроды, если не указано иное. Все сигналы по-разному сравнивались с дополнительным электродом, имплантированным над мозжечком. В последующих экспериментах и наборах данных использовались только лобно-теменные электроды ( n = 72). Провода ЭМГ имплантировали с двух сторон в шейно-ушные мышцы шеи, дифференциально записывая друг друга. Перед экспериментом животным давали 72 часа на восстановление после имплантации и еще 10 дней для привыкания к записывающему кабелю.Сигналы ЭЭГ / ЭМГ были получены с использованием коммерчески доступной системы (Embla; Medcare Flaga, Thornton, CO, USA). Вкратце, сигналы усиливались, фильтровались и аналого-цифровое преобразование до 2000 Гц и субдискретизация до 200 Гц для анализа.
Электроды располагали вдоль изолатеральной (правое полушарие) оси на поверхности коры (Bregma <0,2 мм) во фронтальной (AP = +1,5 мм, ML = +1,5 мм), центральной (AP = -1,0 мм, ML = +2,5 мм) и теменной (затылочной; AP = −3.0 мм, ML = +1,5 мм), у шести мышей (рис. 4а, б). Всем другим мышам имплантировали лобно-теменные биполярные электроды, если не указано иное. Все сигналы по-разному сравнивались с дополнительным электродом, имплантированным над мозжечком. В последующих экспериментах и наборах данных использовались только лобно-теменные электроды ( n = 72). Провода ЭМГ имплантировали с двух сторон в шейно-ушные мышцы шеи, дифференциально записывая друг друга. Перед экспериментом животным давали 72 часа на восстановление после имплантации и еще 10 дней для привыкания к записывающему кабелю.Сигналы ЭЭГ / ЭМГ были получены с использованием коммерчески доступной системы (Embla; Medcare Flaga, Thornton, CO, USA). Вкратце, сигналы усиливались, фильтровались и аналого-цифровое преобразование до 2000 Гц и субдискретизация до 200 Гц для анализа. Отдельную группу мышей (LFP, оптогенетические эксперименты) записывали с использованием сигнального процессора Intan RHD2000, который производил выборку всех каналов с частотой 20000 Гц. Затем сигналы подвергались субдискретизации до 200 Гц и анализировались в Matlab с использованием пользовательских сценариев (см. Ниже), а состояния сна и бодрствования были аннотированы с помощью Somnologica (Medcare Flaga, Thornton, CO, USA) с использованием идентичных критериев.В подгруппе мышей ( n = 9) термистор был вставлен между лобным и париетальным электродами для регистрации температуры поверхности коры. Термистор (серия P20AAA102M, Thermometrics, Northridge, CA) вводили через череп на поверхность правой коры (2,5 мм латеральнее средней линии, 2,5 мм позади брегмы). Постоянный измерительный ток 100 мкА подавался на термистор, и изменения напряжения регистрировались системой сбора данных ЭЭГ / ЭМГ. Напряжение было преобразовано в ° C на основе зависимости сопротивления от температуры отдельных термисторов, предоставленной производителем.
Отдельную группу мышей (LFP, оптогенетические эксперименты) записывали с использованием сигнального процессора Intan RHD2000, который производил выборку всех каналов с частотой 20000 Гц. Затем сигналы подвергались субдискретизации до 200 Гц и анализировались в Matlab с использованием пользовательских сценариев (см. Ниже), а состояния сна и бодрствования были аннотированы с помощью Somnologica (Medcare Flaga, Thornton, CO, USA) с использованием идентичных критериев.В подгруппе мышей ( n = 9) термистор был вставлен между лобным и париетальным электродами для регистрации температуры поверхности коры. Термистор (серия P20AAA102M, Thermometrics, Northridge, CA) вводили через череп на поверхность правой коры (2,5 мм латеральнее средней линии, 2,5 мм позади брегмы). Постоянный измерительный ток 100 мкА подавался на термистор, и изменения напряжения регистрировались системой сбора данных ЭЭГ / ЭМГ. Напряжение было преобразовано в ° C на основе зависимости сопротивления от температуры отдельных термисторов, предоставленной производителем.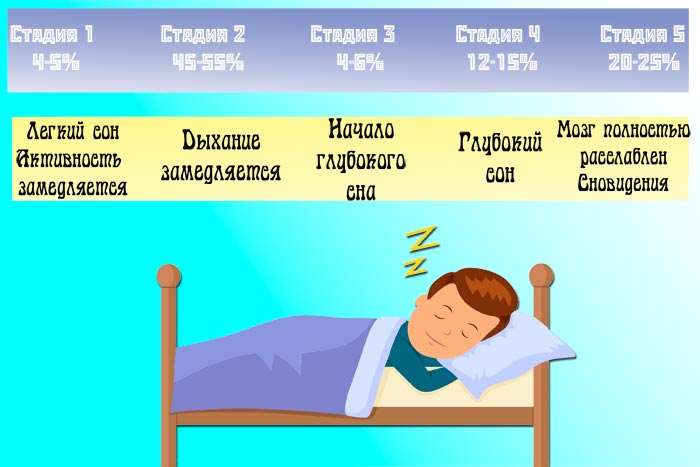
Различение состояний сна и бодрствования на основе ЭЭГ / ЭМГ
После сбора данных сигналы были отфильтрованы на предмет артефактов в линии электропередачи, возникающих в результате циклического изменения мощности переменного тока (50 Гц). Оценка состояний сна и бодрствования на основе ЭЭГ / ЭМГ была достигнута с использованием ранее опубликованных критериев 28 . Последовательные 4-секундные эпохи были классифицированы как бодрствование, сон с небыстрым движением глаз (NREM) или REM-сон с использованием визуального осмотра без знания условий записи.
Установка термоэлектрического генератора
Для контроля уровней корковой температуры во время восстановления SD у группы мышей ( n = 4) алюминиевый радиатор был помещен на череп, противоположный термистору (см. Выше и рис.5g), используя термопасту (WLP-1, S + S Regeltechnik GmbH), чтобы обеспечить эффективную передачу температуры. Термоэлектрический генератор (ТЭГ; модуль Пельтье: Laird Thermal Systems, Inc. 45850-503¨, Моррисвилл, США) затем прикрепляли с помощью стоматологического цемента (Paladur) на 2 см выше животного и прикрепляли к кабелю для записи ЭЭГ / ЭМГ. Температурные манипуляции были достигнуты с помощью настольного источника питания (RND 320-KD3005P, Distrelec, Nänikon, Швейцария) для управления входной силой тока на ТЭГ. Выходные температуры были откалиброваны до экспериментов.Тепловизионные изображения были получены с помощью инфракрасной тепловизионной камеры (модель E30, FLIR Systems, Уилсонвилл, США) и проанализированы с использованием программного обеспечения FLIR tools (версия 6.4, FLIR Systems, Уилсонвилл, США).
Имплантация тетрода LFP
Тетроды были сконструированы в лаборатории с использованием четырех жил скрученной вольфрамовой проволоки диаметром 10 мкм, которые затем были прикреплены золотыми штырями к плате интерфейса электродов. Затем они были помещены в 2 таламические и 3 корковые структуры: центральный таламус средней линии (CMT) (AP -1,7 мм, ML +1.0 мм, DV -3,8 мм, 15 °), поясная извилина (AP +1,8 мм, ML +0,2 мм, DV -1,6 мм), антеродорсальные ядра таламуса (AD) (AP -0,9 мм, ML ± 0,8 мм, DV — 3,2 мм), бочкообразной коры (AP -2,0 мм, ML +2,2 мм, DV -1,1 мм) и зрительной коры (AP -3,3 мм, ML +2,5 мм, DV -0,9 мм) и прикреплены к черепу стоматологическим акрилом. (C&B Metabond). Оптические волокна диаметром 200 мкм были помещены в CMT (AP -1,7 мм, ML +1,0 мм, DV -3,8 мм, 15 °) и закреплены тем же стоматологическим акрилом. Наконец, имплант был стабилизирован с использованием метилметакрилатного цемента, и животному дали возможность восстановиться в домашней клетке на нагревательном мате.Перед началом записи животным давали как минимум 5 дней для восстановления.
Оптогенетические эксперименты
Мышей C57Bl6 в возрасте 6 недель анестезировали изофлураном (1,0–1,5% с кислородом) и помещали в стереотаксический аппарат (модель 940, David Kopf Instruments). Инъекции AAV выполнялись с помощью шприца Hamilton объемом 10 мкл, присоединенного к инфузионному насосу (модель 1200, Harvard Apparatus), в CMT (AP –1,7 мм, ML + 1,0 мм, DV –3,8 мм, 15 °, 100 нл и выполнялись при 0,1 мкл мин -1 , а затем игла оставалась на месте на 10 мин для облегчения диффузии.Животным вводили AAV2-CaMKII-E1fa-ArchT3.0-EYFP (ArchT) для оптического подавления нейронов CMT. Все плазмиды были получены из Vector Core Facility Университета Северной Каролины. Перед началом оптогенетических экспериментов животным дали 21 день на выздоровление. Животных обрабатывали инструментами для регистрации тетрода (см. Выше: имплантация тетрода LFP) через 3–4 недели после инъекции вируса, чтобы дать достаточно времени для экспрессии опсина.
Оптические волокна, соединенные с патч-кордой, покрытой черной трубкой с разделительной трубкой (дорические линзы), были дополнительно покрыты черным лаком для уменьшения оптических утечек от лазера.Животные были приучены быть привязанными на срок до 8 часов в день до возобновления нормального эпизода сна и бодрствования на основании записей ЭЭГ от ZT4-9. Эксперименты проводились с ZT4-8, следуя SD (рис. 5b). Ингибирование достигалось зеленым лазером (532 нм; LRS-0532-GFM-00100-03, Laserglow Technologies) через 10 с после начала NREMS. Выход контролировался с помощью генератора импульсов TTL (Master-9, AMPI или PulsePal 2, Sanworks), полученного совместно со всеми записями. Во время периодов восстановления после депривации сна 10-минутное скользящее окно использовалось для расчета мощности δ1 и δ2 и относилось к тем же частотам в течение предыдущего базового дня.В отдельном эксперименте (фиг. 5c) оптогенетическое молчание выполняли в течение 10 с во время NREMS между ZT4-8 в исходных условиях (без лишения сна). Мощность δ1 и δ2 рассчитывалась в 10-секундных окнах до, во время и после ингибирования.
Протокол депривации сна
После> 10 дней восстановления после операции у мышей регистрировали ЭЭГ / ЭМГ в общей сложности 96 часов. В течение первых 48 ч животные оставались в исходных условиях, не нарушая покоя. Начиная с третьего дня, мышей подвергали 6-часовому SD, начиная с легкого начала (ZT0), с осторожным обращением 28 .Затем животным давали возможность восстановиться еще на 42 часа. Другие эксперименты, представленные в этой статье, также включали SD, начиная с различных ZT в течение 24-часового периода (рис. 3c 27 ) или различной продолжительности (2- и 4 часа, рис. 3b).
Для записи LFP и оптогенетических экспериментов животные были взяты из их домашних клеток и помещены в новые в ZT0 с чистой подстилкой, едой и водой, в дополнение к новому пластиковому объекту. 4-часовое стандартное отклонение выполняли перед тем, как мышей переводили в их исходные клетки для сбора данных между ZT4 и -9.Подробнее см. 30 .
Дыхательные записи
Для определения частоты дыхания у мышей регистрировали ЭЭГ / ЭМГ в сочетании с пьезоэлектрической системой (Signal Solutions, LLC, Лексингтон, Кентукки, США), которая использует связанные с дыханием движения для оценки состояния сна и бодрствования 76 . Вкратце, пьезоэлектрическая платформа состоит из поликарбонатной клетки и пола, покрытого пленкой поливинилидиндифторида (PVDF) (17,8 см × 17,8 см, толщиной 110 мкм; Measurement Specialties, Inc., Хэмптон, Нью-Йорк), покрытые стандартным пометом для грызунов. Вектор с 6 признаками извлекается из пьезоэлектрического сигнала для последовательных 4-х периодов, который согласовывался с состояниями сна и бодрствования, полученными на основе ЭЭГ / ЭМГ. Частота дыхания — одна из векторных характеристик.
Анализ сигналов ЭЭГ в частотной области
Спектральная плотность мощности для каждой 4-секундной эпохи ЭЭГ была сгенерирована с использованием дискретного преобразования Фурье после использования окна Хэмминга, что дало спектры плотности мощности (0–100 Гц) с разрешением по частоте 0.25 Гц. Бункеры, содержащие частоты от 49 до 51 Гц, были исключены из-за артефактов в электросети у некоторых животных. Эпохи, содержащие артефакты сигнала, были отдельно идентифицированы для включения в количественную оценку состояния, но не в спектральный анализ. Следует отметить, что частоты аннотируются на основе средней точки интервала 0,25 Гц (например, 2,0 Гц = 1,875–2,125 Гц). δ-мощность рассчитывалась для 4-секундных эпох NREMS, окруженных эпохами без артефактов одного и того же поведенческого состояния. Сводные эпизоды NREMS были определены как непрерывные последовательности продолжительностью не менее 32 секунд (8 4-секундных эпох) на основе визуального осмотра и предыдущих публикаций 7,28 .
Временной анализ спектральной мощности в NREMS
Спектральная полоса (0,25–90 Гц) была разделена на компоненты для изучения динамики во времени отдельных полос: δ (1–4 Гц), δ1 (0,75–1,75) Гц), δ2 (2,5–3,5 Гц), тета (6–9 Гц), сигма (10–15 Гц) и бета / низкая гамма (18–45 Гц). Для контроля межиндивидуальных различий абсолютная спектральная мощность каждого из этих частотных диапазонов была выражена в значениях, рассчитанных для периода в течение двух базовых дней с самым низким средним значением, соответствующим последним 4 часам светового периода (ZT8– 12).Значения были усреднены по периодам времени, содержащим равное количество 4-секундных эпох, оцененных как NREMS, называемых квантилями. Для светлых периодов было выбрано 12 квантилей, для темных периодов 6 и для светлого периода сразу после SD 25.
Временной анализ δ-динамики при переходах в NREMS
После SD, динамика суб- Полосы (δ1 и δ2) анализировались за 4-секундную эпоху при переходах от следа или REMS к NREMS, аналогично ранее опубликованным методам 7 .Переходы были разделены на четыре группы, первые три квантиля рассчитывались, начиная с восстановления стандартного отклонения и в последующие часы после точки поворота (см. Раздел ниже). Переход от бодрствования / REMS к NREMS был определен как ≥4 последовательных 4-секундных периода NREMS, которым предшествовали ≥8 баллов, оцениваемых либо как пробуждение, либо как REMS. Спектральная мощность δ1- и δ2 рассчитывалась для этих переходов, начиная с 1 мин до и 2 мин после, а затем усреднялась по этим последующим 3-минутным окнам. Значения были выражены относительно изменений в течение двух дней регистрации исходного уровня (ZT8-12), как для δ-мощности.Кривые роста до максимума для первого квантиля использовались для количественной оценки быстрого роста δ2 по сравнению с δ1 с использованием SigmaPlot (версия 12.5 SysStat 2011).
Анализ наклона медленной волны
Необработанные сигналы ЭЭГ с частотой 200 Гц были импортированы в Matlab (v. 2019b, Mathworks Inc.) в дополнение к состояниям сна и бодрствования. Затем ЭЭГ фильтровали по δ-спектру (Чебышевский тип-II 0,5–4,5 Гц, полоса пропускания 0,1 и 10 Гц, с использованием функции цифровой фильтрации нулевой фазы в Matlab, filterfilt ), аналогично другим 16,19,25 , 77 .Для обнаружения медленных волн использовался специально разработанный алгоритм Matlab, основанный на переходах через ноль и реконструкции волн, близких к другим 16,26 . Обнаружение SW NREMS было достигнуто с использованием следующих шагов: (1) были определены нулевые пересечения, (2) были определены окружающие локальные максимумы и минимумы, (3) была применена пороговая обработка для контроля амплитуды SW (см. Ниже) и ( 4) математический наклон на основе амплитуды SW и частоты (периода) был определен во всех возможных направлениях и комбинациях (см. Дополнительный рис.1). Наклоны, которые были проанализированы в рукописи, относятся к переходу через ноль к SW максимальной амплитуды, которые начинаются с отрицательного отклонения. Хотя все наклоны были рассчитаны (дополнительный рис. 1), их средние относительные изменения во времени были почти одинаковыми. Затем данные были сгруппированы и усреднены для эпизодов NREMS> 32 с. Изменения наклона, амплитуды и периода выражались в процентах от базовых значений ZT8-12 NREMS, как и для спектральной мощности.
Вышеупомянутый алгоритм обнаруживал SW в течение всего 96-часового эксперимента для всех эпизодов NREMS, а нижний и верхний пороговые значения применялись для захвата добросовестных истинных SW NREMS на основе визуального осмотра и ранее опубликованных критериев 16,25 .Верхние пороги были установлены в 6 раз больше стандартного отклонения. амплитуды всех обнаруженных сигналов. Нижний порог был установлен на уровне 95% вероятности обнаружения SW во время REMS, с идеей, что SW не будут появляться во время REMS. Распределение амплитуды см. На дополнительном рис. 1d. После обнаружения SW SW были классифицированы в соответствии с их частотой в окнах 0,25 Гц (между 0,75 Гц и 4,5 Гц) и проанализированы на предмет падения (и плотности min −1 ) и амплитуды (см. Рис. 2a, b и дополнительный рис.2а, б).
Средние формы волны во время эпизодов NREMS (рис. 2g, дополнительный рис. 2d) сначала были локализованы с использованием отфильтрованного сигнала ЭЭГ для δ1 и δ2, а затем интерполированы для соответствия 800 мс (δ1) и 300 мс (δ2) для лучшего сравнения отдельные ПВ разных частот в этих диапазонах. Затем временные коды этих SW были повторно использованы для извлечения необработанных нефильтрованных сигналов. Чтобы определить начало и смещение вложенных δ2-волн, местоположения δ1-сигналов были сохранены, а затем необработанный сигнал был отфильтрован только для δ2 (2.5–3,5 Гц) с использованием тех же характеристик фильтра Чебышева (см. Выше). Сигналы δ2, которые попадали в эти временные коды, считались вложенными, и их начало / смещение от форм сигналов δ1 вычислялось. Следует отметить, что хотя для частотного (спектрального) анализа использовались 38 мышей, одна мышь была удалена для анализа наклона SW из-за присутствия промежуточных артефактов во время определенных эпизодов NREMS, которые затрудняли обнаружение перехода через ноль и определение порога у этого животного.
Анализ физиологических переменных
Все переменные, представленные на рис.7 и дополнительный рис. 6 (за исключением частоты сердечных сокращений) были выражены в относительных значениях и сгруппированы в идентичные квантили, как δ1 и δ2 (рис. 5a). У мышей это было 25 квантилей (5 до точки поворота и 20 после). Конкретный анализ заключается в следующем: более высокие частоты (рис. 7b) вычисляются аналогично δ-power, за исключением различных частот (18–45 Гц). Мышечный тонус (рис. 7c) рассчитывается как относительные изменения средней дисперсии ЭМГ по эпизодам NREMS. Дисперсия ЭМГ рассчитывается на основе суммы квадратов расстояний от среднего, деленных на количество выборок (800).Значения выражены относительно дисперсии ЭМГ во время NREMS от исходного уровня (ZT8-12). Температура мозга (рис. 7d) представлена как разница во время квантилей NREMS от средних значений в течение 6-часового периода бодрствования (SD), предшествующего восстановлению. Дыхание (рис. 7e) описано в предыдущем разделе и выражается в абсолютной частоте.
Иерархическая кластеризация для определения разделения δ-полос
Кластерный анализ эталонной (базовой ZT8-12) мощности на 0,25 Гц, как на рис.2d, было получено на основе средних значений от 38 мышей для первых 25 квантилей NREMS во время восстановления SD (см. Основной текст). Функция Matlab , кластерограмма была использована для создания дендрограммы и тепловой карты с использованием иерархической кластеризации на основе метрики евклидова расстояния и средней связи.
Обнаружение состояний ВКЛ / ВЫКЛ и частоты возбуждения нейронов
Обнаружение состояний ВКЛ / ВЫКЛ было достигнуто с использованием сигналов LFP / ЭЭГ с полосовой фильтрацией (0,5–3 Гц) в прямом и обратном направлениях ( filterfilt , Матлаб).Отдельные состояния включения / выключения были обнаружены с использованием метода перехода через ноль этих сигналов. Начало включенного состояния было определено как переход от отрицательного к положительному и тех состояний ВКЛ / ВЫКЛ с амплитудой <1 с.о. из средних и короче 200 мс, были исключены. Чтобы извлечь активность нескольких устройств из LFP, сигналы подвергались полосовой фильтрации (600–4000 Гц, эллиптический фильтр четвертого порядка, пульсация полосы пропускания 0,1 дБ, затухание в полосе задерживания –40 дБ) и детектировались с использованием порога, в 7,5 раз превышающего медианное значение абсолютное значение отфильтрованного сигнала.Затем отдельные единицы были идентифицированы после сортировки с помощью набора инструментов WaveClus (v.3). Средняя частота возбуждения была рассчитана на основе количества всплесков, присутствующих в секунду в эпизодах NREMS, и выражена в Гц.
Эксперименты на людях
Всего в четырех исследованиях, использованных для этого анализа, приняли участие 110 здоровых молодых людей. Представленные совокупные результаты одинаково наблюдались в каждом исследовании отдельно. Все исследования на людях были одобрены независимыми институциональными наблюдательными советами и соответствовали соответствующим законам и постановлениям об исследованиях на людях и Хельсинкской декларации Всемирной медицинской ассоциации.Ни один участник не проехал более двух часовых поясов за предшествующие 3 месяца и не страдал диагностированным расстройством нервной системы или другим острым заболеванием. Кроме того, перед исследованием испытуемые прошли предварительный скрининг в лаборатории, чтобы подтвердить отсутствие каких-либо нарушений сна, а также отказаться от приема лекарств и рекреационных наркотиков. Все субъекты подписали информированное согласие и получили финансовую компенсацию за участие. Подробнее см. Предыдущие публикации 32,33,34,35 .Некоторые испытуемые были исключены из четырех исследований из-за артефактов сигнала ЭЭГ, что затрудняло временной анализ спектральных диапазонов. Для топографического анализа и анализа частоты сердечных сокращений каждый объект был удален на предмет артефактов сигнала.
Сбор данных
Непрерывная запись данных ЭЭГ, ЭОГ, ЭМГ и ЭКГ была получена во время исходного уровня и после лишения сна. Стадии сна и бодрствования визуально оценивались через 20-е эпохи (производное C3-A2) с использованием Rembrandt® Analysis Manager (версия 8; Embla Systems, Broomfield, CO, USA).Артефакты, связанные с движением и возбуждением, были визуально идентифицированы и исключены из последующего анализа. Аналоговые сигналы от каждого вывода (см. Рис. 6c), дискретизированные с частотой 256 Гц, были отфильтрованы [фильтр верхних частот (–3 дБ при 0,15–0,16 Гц) и фильтрация нижних частот (–3 дБ при 67,2 Гц)]. Спектры ЭЭГ рассчитывались так же, как и у мышей (см. Выше), с использованием Matlab для каждой 4-секундной эпохи и усреднялись в соответствии с назначенным состоянием сна и бодрствования для каждого доступного вывода. Циклы NREM / REM-сна сначала были определены в соответствии с исходными условиями и условиями восстановления, временная динамика δ-мощности в течение всех оцениваемых эпох визуально проверялась, чтобы разделить первые два эпизода, между которыми не было длительного эпизода REMS.Анализировались только первые 8 часов восстановительного сна. Мощность ЭЭГ была выражена как среднее значение за ночь за исходный период. Каждый эпизод NREMS был разделен на 10 квантилей, каждый из которых содержал равное количество 20-секундных периодов, оцененных как NREMS. Для сравнения относительных изменений на коже черепа у подгруппы людей (рис. 6c, d) использовалась средняя мощность по всем участкам для каждого поддиапазона δ во время исходного уровня. Средняя мощность электродного участка во время восстановления SD затем оценивалась в максимальном квантиле в течение δ1 и δ2 во время каждого эпизода NREMS и выражалась как отношение log2 к исходному уровню.
Динамика сердечного ритма во время NREMS была выражена как изменения RR-интервалов в процентах от индивидуального среднего исходного RR-интервала NREMS (1,07 ± 0,02 с или 57 ± 1 ударов мин. -1 ). Вариабельность сердечного ритма как показатель симпатовагального баланса оценивалась путем определения отношения LF / HF; то есть соотношение между низкочастотными (0,04–0,15 Гц) и высокочастотными компонентами (0,15–0,4 Гц) спектра сердечного ритма,
Статистический анализ
Все статистические анализы выполнялись в любой из программ Statistica 8.0 (Statsoft, inc) или с помощью встроенных функций MATLAB. Обратите внимание, что p-значения из Statistica ограничены 17 десятичными знаками (т. Е. p, = 1.0E-17 является наименьшим сообщенным значением). Распределение сна и бодрствования, спектральная мощность ЭЭГ и динамика определенных частотных диапазонов оценивались с использованием двух- или трехфакторного дисперсионного анализа с повторными измерениями (rANOVA). Статистическая значимость была принята как p <0,05, и все результаты представлены как средние значения ± стандартная ошибка среднего. Апостериорный тест Тьюки использовался для определения значимых эффектов и взаимодействий и исправлен для множественного сравнения.Сравнение двух групп было достигнуто с помощью двусторонних t-критериев Стьюдента. По возможности использовались внутрипредметные парные t -тесты. Статистическая методология дополнительно описана в разделе результатов и в подписях к рисункам. Были выполнены линейные и кусочные регрессии, а коэффициенты корреляции Пирсона статистически сравнивались с помощью t-критерия после нормализации с использованием преобразования Фишера-Z. Экспоненциальные нелинейные затухания были подогнаны к дополнительному рис. 4b, c с использованием функции fitnlm в Matlab.
Краткое изложение отчета
Дополнительная информация о дизайне исследования доступна в Резюме отчета об исследовании природы, связанном с этой статьей.
Какие стадии сна?
2. Я слышал, есть стадии сна. Кто они такие? Обычно спящие проходят пять стадий: 1, 2, 3, 4 и быстрый сон (быстрое движение глаз). Эти стадии проходят циклически от 1 до REM, затем снова начинаются со стадии 1. Полный цикл сна занимает в среднем от 90 до 110 минут.
1 этап
Стадия 1 — это легкий сон, при котором вы засыпаете и просыпаетесь, и вас легко разбудить. На этом этапе глаза медленно двигаются и мышечная активность замедляется. На этой стадии многие люди испытывают внезапные сокращения мышц, которым предшествует ощущение падения.
2 этап
На стадии 2 движение глаз прекращается, и мозговые волны замедляются, лишь изредка появляются быстрые мозговые волны.
Стадия 3 и 4 (дельта-сон или глубокий сон)
Когда человек входит в стадию 3, чрезвычайно медленные мозговые волны, называемые дельта-волнами, перемежаются с более мелкими, более быстрыми волнами.На стадии 4 мозг почти исключительно производит дельта-волны. Стадии 3 и 4 называются глубоким сном или дельта-сном, и разбудить кого-либо от них очень сложно. В глубоком сне нет движения глаз или мышечной активности. Это когда некоторые дети испытывают ночное недержание мочи, лунатизм или ночные кошмары. Глубокий сон восстанавливает организм.
Быстрый сон
Во время быстрого сна дыхание становится более частым, нерегулярным и поверхностным, глаза быстро дергаются, а мышцы конечностей временно парализуются.Мозговые волны на этой стадии увеличиваются до уровня, наблюдаемого, когда человек бодрствует. Кроме того, увеличивается частота сердечных сокращений, повышается кровяное давление, у мужчин развивается эрекция, и тело теряет часть способности регулировать свою температуру. Это время, когда происходит большинство сновидений, и, если человек просыпается во время быстрого сна, он может вспомнить сны. У большинства людей каждую ночь наблюдается от трех до пяти интервалов быстрого сна.
Медленный сон обычно наступает в первой половине ночи, быстрый сон — во второй половине.
Младенцы проводят почти 50% своего времени в фазе быстрого сна. Взрослые тратят почти половину времени на сон на стадии 2, около 20% в фазе быстрого сна, а остальные 30% распределяются между тремя другими стадиями. Пожилые люди проводят все меньше времени в фазе быстрого сна.
Регулирование и функциональные корреляты медленного сна
ВВЕДЕНИЕ
Считается, что сон важен для эффективного дневного функционирования. Сон с глубокими небыстрыми движениями глаз (NREM), также известный как медленный сон (SWS), считается наиболее восстанавливающей стадией сна и связан с качеством сна1,2 и поддержанием сна.3 Однако еще многое предстоит узнать о функции SWS и ее влиянии на другие физиологические процессы и дневное функционирование. В этом обзоре кратко описаны некоторые характеристики SWS, способы его измерения и регулирования, а также влияние SWS на склонность ко сну и поддержание сна.
ЧТО ТАКОЕ SWS?
Сон состоит из двух отдельных фаз: сна с быстрым движением глаз (REM) и сна NREM, которые можно идентифицировать с помощью полисомнографии. Эти две фазы сна имеют различные наборы связанных физиологических изменений, нейроанатомических субстратов и нейрохимических коррелятов.
У молодых людей REM-сон и NREM-сон чередуются на протяжении всего эпизода сна примерно по 80–120-минутным циклам, при этом REM-сон составляет примерно 18-25% от общего времени сна4.
Согласно классификации Rechtschaffen and Kales, 5 NREM-сон подразделяется на четыре стадии: стадии 1, 2, 3 и 4. Стадии 1 и 2 часто считаются «легким сном», тогда как стадии 3 и 4 считаются «глубоким сном», и вместе они считаются называется SWS (рис. 1). У молодых людей стадии 1 и 2 составляют примерно 4–9% и 45–60% от общего времени сна, соответственно, в то время как нормальные субъекты проводят от 10 до 25% общего времени сна в SWS.4 В новом руководстве по оценке Американской академии медицины сна различие между стадией 3 и 4 сном больше не проводится, и SWS обозначается как NREM3 или N3. 6 Определяющими характеристиками сна NREM на электроэнцефалограмме (ЭЭГ) являются : веретена сна с характерными для них «веретенообразными» изменениями амплитуды колебаний ЭЭГ 12–14 Гц; К-комплексы, длящиеся не менее 0,5 секунды и состоящие из четко очерченной отрицательной резкой волны, за которой сразу следует положительный компонент; и медленные волны или дельта-волны с их характерной медленной частотой (<2 Гц) и высокой амплитудой (> 75 мкВ).Активность веретена сна и медленные волны варьируются в пределах NREM-сна, и это изменение является основой, по которой NREM-сон подразделяется на стадии 1–4. Хотя медленные волны и веретена сна присутствуют на стадиях 2, 3 и 4, веретена более распространены на стадии 2 сна, тогда как медленные волны доминируют на ЭЭГ на стадиях 3 и 4,7. Нейрофизиологические корреляты и механизмы, лежащие в основе этих явлений ЭЭГ, были выяснены в part.8
Рисунок 1Медленный сон и медленная волновая активность (SWA) во время ночного сна у здорового 22-летнего мужчины (Dijk, неопубликованные данные).REM, быстрое движение глаз.
SWS определяется на основе визуальной оценки частоты, амплитуды и встречаемости низкочастотных колебаний в ЭЭГ <2 Гц в соответствии со стандартизованными критериями.5,6 Вариация ЭЭГ во время медленного сна также может быть определена количественно. спектральным анализом или другими методами автоматического анализа сигналов, такими как анализ амплитуды периода. Используя спектральный анализ, вариации медленных и дельта-колебаний фиксируются с помощью показателя, называемого медленной волновой активностью (SWA), который представляет плотность мощности в диапазоне 0.Диапазон 75–4,5 Гц. Вариация активности веретена во сне фиксируется с помощью плотности мощности в диапазоне 12–14 Гц или 12–15 Гц (сигма или частотная активность веретена), но также может быть определена количественно с помощью аналитических мер, определяющих фазовую активность ЭЭГ7,9
Динамика визуально идентифицируемых стадий сна и обнаруженного компьютером SWA проиллюстрирована на рисунке 1. Динамика SWA четко иллюстрирует непрерывный характер процесса NREM-сна с небольшими доказательствами ступенчатых изменений SWA при переходах стадий сна. .Неудивительно, что максимальные уровни SWA присутствуют в стадии 4 сна, который считается более глубоким, чем стадия 3. Однако разница между стадиями сна 3 и 4 незначительна, и обе можно рассматривать как дельта-сон или SWS, и они называется N3 в соответствии с новой классификацией.6 Во время стадии 1 и быстрого сна присутствует очень мало SWA, а уровни во время стадии 2 являются промежуточными между стадией 1 / REM и SWS. На рисунке 1 очень четко показано, как SWA постепенно увеличивается в течение каждого эпизода NREM, а затем быстро уменьшается незадолго до начала эпизода REM.Более того, как скорость роста, так и максимальный уровень SWA снижаются в последовательных эпизодах NREM. Динамика SWA внутри и между циклами NREM была подробно описана и может быть смоделирована с помощью компьютерного моделирования.10 Динамика SWA тесно связана с рядом других показателей ЭЭГ, которые, как считается, отражают связность в корковых сетях.
SWS и SWA также коррелируют с вариациями эндокринных и сердечно-сосудистых параметров. Например, скорость секреции гормона роста человека является максимальной во время SWS12, а вегетативная нервная система переходит от симпатической к парасимпатической доминирующей роли во время SWS.13,14 Нарушение SWS может привести к сдвигу вегетативного баланса и связанным с ним изменениям реакции на глюкозу. Фактически, всего три ночи нарушения SWS у людей могут привести к снижению способности адекватно реагировать на глюкозную нагрузку. Эти изменения отражаются в снижении чувствительности к инсулину и могут указывать на связь между нарушением SWS и повышенным риском диабета, возможно, опосредованным влиянием нарушения SWS на вегетативную нервную систему.15,16 Таким образом, SWS — это не только феномен ЭЭГ или состояние мозга, но также состояние, которое влияет на многие физиологические системы.
Циркадная регуляция SWS
Циркадная ритмичность — главный детерминант физиологии, эндокринологии и поведения человека.17 Циркадный ритм, о котором мы больше всего знаем, — это цикл сна и бодрствования, но другие основные ритмы в физиологии и эндокринологии включают основные температура тела, мелатонин в плазме и кортизол. Все эти физиологические и поведенческие ритмы управляются супрахиазматическими ядрами (SCN) гипоталамуса.Поражения этой структуры устраняют ритмичность этих переменных, и исследования поражений также продемонстрировали, что SCN участвует в определении времени сна и бодрствования у животных.18,19 Кроме того, у людей вызывает поражение в области, гомологичной SCN, или рядом с ней. значительное нарушение цикла сон-бодрствование. 20 Эти данные подтверждают роль SCN в циркадной регуляции сна. В настоящее время установлено, что циркадные ритмы генерируются петлями транскрипционно-трансляционной обратной связи основного набора «часовых» генов.21 Эти гены ритмично экспрессируются в SCN, но также и в других областях мозга, а также в периферических тканях и лейкоцитах.22 Вариации этих генов связаны с вариациями времени и структуры сна и могут приводить к ним23
Поведенческие Доказательства участия системы циркадного ритма в регуляции сна были ранее получены в экспериментах с произвольным запуском, в которых испытуемые жили в условиях, свободных от временных сигналов. В этих условиях циклы сна и бодрствования десинхронизируются не только с внешними 24-часовыми днями, но в некоторых случаях также с внутренним ритмом температуры тела.Подробный анализ полисомнографически записанного сна во время этих спонтанных проявлений внутренней десинхронии показал, что продолжительность сна и структура сна определяются взаимодействием циркадного осциллятора и осциллятора сна-бодрствования24–26
Роль циркадного водителя ритма в регуляции. NREM и REM-сон дополнительно исследовали с использованием протокола принудительной десинхронии, в котором испытуемые были запланированы на циклы сна-бодрствования, выходящие далеко за пределы циркадного диапазона, например.g., 28 или 20 часов.27,28 Циркадные кардиостимуляторы не могут следовать этим 28- или 20-часовым циклам, и, следовательно, цикл сна-бодрствования десинхронизируется с циркадными ритмами температуры тела, мелатонина, кортизола и т. д. Эпизоды сна происходят во время сна. все фазы эндогенного циркадного цикла и SWA снижается во время всех эпизодов сна, почти независимо от циркадной фазы. В этих исследованиях циркадная модуляция SWS и SWA была незначительной (рис. 2). Циркадный надир ритма SWA был обнаружен в ранние утренние часы, когда с точки зрения циркадных ритмов сонливость находится на пике.В отличие от SWA, активность веретена сна демонстрировала заметную циркадную модуляцию, при этом максимальная активность совпадала с ритмом мелатонина.27,29,30 Эти исследования подтвердили, что быстрый сон также находится под сильным циркадным контролем, причем пик его ритма приходится на 1 –2 часа после надира температуры, т. Е. Близко к привычному времени бодрствования. Эти исследования принудительной десинхронии без сомнения продемонстрировали, что, в то время как активность веретена сна и быстрый сон в значительной степени определяются циркадными часами, SWA в первую очередь определяется временем бодрствования, почти независимо от циркадной фазы.Интересно, что SWA также практически не зависит от другого цикла: менструального цикла. Активность SWA снижается во всех эпизодах сна, во всех фазах менструального цикла. Напротив, активность веретена сна значительно модулируется по фазам менструального цикла.31
Рисунок 2A : Динамика медленной активности (SWA) во время эпизодов сна здорового субъекта во время протокола принудительной десинхронии. Данные нанесены на двойной график, т.е. два последовательных 24-часовых периода нанесены рядом друг с другом и друг под другом.Сплошная линия представляет прогрессирование времени надира ритма внутренней температуры тела. Обратите внимание, что SWA снижается во всех эпизодах сна, независимо от того, когда эпизоды сна начинаются относительно надира основной температуры тела.27 B : Оценка циркадных (левая панель) и зависимых от сна (правая панель) изменений в SWA ( верхняя панель) и внутренняя температура тела (нижняя панель) у 8 здоровых субъектов, живущих в среде, свободной от временных сигналов, в течение 33–36 дней и запланированных на 28-часовой цикл отдыха и активности.27 Обратите внимание, что несмотря на наличие устойчивой зависимой от сна модуляции SWA, ее циркадная амплитуда очень мала, что указывает на то, что SWA не сильно модулируется циркадной фазой. NREM, небыстрое движение глаз. Воспроизведено с разрешения Дейка и Чейслера27.
ГОМЕОСТАТИЧЕСКОЕ РЕГУЛИРОВАНИЕ SWS
Гомеостатическое регулирование — это контроль и поддержание стабильного, постоянного состояния, включающее множество регулирующих механизмов. Исследования депривации сна, которые увеличивают давление сна, а также исследования продления сна и дневного сна, которые приводят к снижению давления сна, были использованы для изучения того, контролируется ли SWS гомеостатически.Это было установлено на раннем этапе современной эры исследований сна, в частности, благодаря обширной работе Уэбба и других с использованием визуальной оценки SWS.32 Совсем недавно этот гомеостатический процесс был исследован в исследованиях с использованием количественных методов ЭЭГ, таких как спектральный анализ. анализ амплитуды периода и методы ЭЭГ с высокой плотностью, такие как картирование мозга.33–35
В одном из таких исследований девять субъектов бодрствовали в течение 36 часов, при этом исходный сон начинался в 23:00, а восстановительный сон — в 19:00.36 Когда восстановительный сон начинается в этой циркадной фазе, а субъекты не пробуждаются исследователем, могут наблюдаться очень длительные эпизоды восстановительного сна (> 12 часов). Анализ динамики SWA во время многих циклов NREM – REM восстановительного сна показал, что SWA в NREM-сне первоначально была значительно увеличена по сравнению с исходным уровнем, а затем достигла стабильного уровня во второй части эпизода сна. экспериментов было обнаружено мало доказательств циркадного контроля SWS / SWA (рис. 3).
Рисунок 3Динамика медленной активности (SWA) и преобладающего состояния бдительности (N — сон с небыстрым движением глаз; R — сон с быстрым движением глаз; W — бодрствование) во время базового сна и восстановительного сна после 36 часов лишения сна у людей36 и 24-часовое лишение сна у крыс (Franken, неопубликованные данные). Воспроизведено с разрешения Dijk et al36 и Paul Franken.
Гомеостатическая регуляция SWS также была продемонстрирована на большом количестве видов (грызунов).У крыс SWA колеблется синхронно с циклами NREM – REM, хотя и с более короткой периодичностью, чем у людей, что отражает меньший размер мозга (рис. 3) 37. в течение дня. Восстановление после депривации сна сопровождается заметным увеличением продолжительности медленного сна и усилением SWA, особенно во время начальной части восстановительного сна, как и у людей (рис. 3). Манипуляции с системой суточного времени за счет сокращения или удлинения фотопериода оказывают преимущественное влияние на 24-часовой режим сна, но мало влияют на 24-часовой базовый уровень общего времени сна или реакцию на лишение сна.Таким образом, изменения продолжительности фотопериода и сопутствующие изменения в циркадной организации не влияют на основной процесс, регулирующий сон у дневных грызунов38 или людей 39, подтверждая надежность гомеостатического контроля SWS / SWA.
Лишение сна и изменение продолжительности фотопериода можно рассматривать как крайнее вмешательство в изучение регуляции сна. Напротив, исследования дневного сна предлагают более взвешенный подход. Несколько исследований дневного сна продемонстрировали, что в пределах физиологического диапазона продолжительности бодрствования SWS регулируется очень точно.40–42 В исследовании дневного сна с участием здоровых женщин, дневной сон планировался в разное время дня, с интервалом не менее 2 дней, при этом также поддерживался регулярный режим сна и бодрствования. Анализ SWA во время этих дремот показал постепенное увеличение SWA в течение дня бодрствования. Таким образом, тогда как во время консолидированного сна SWA снижается, SWA увеличивается во время бодрствования (рис. 4). Интересно отметить, что в этих исследованиях склонность к засыпанию диссоциировалась с течением времени SWA.Таким образом, SWA монотонно увеличивалась со временем бодрствования, тогда как склонность ко сну в дневное время демонстрировала более сложную картину с локальным максимумом во второй половине дня, за которым следовало снижение и снижение в зоне поддержания бодрствования, незадолго до привычного отхода ко сну.
Рисунок 4Медленно-волновая активность (SWA) в сне с небыстрым движением глаз (NREM) во время исходного сна и в первом NREM-сне (открытые кружки) и в первых эпизодах NREM-сна (закрытые кружки), начиная с 10 лет. 12, 14, 16, 18, 20 и 22 часа у семи здоровых женщин.33 Данные нанесены на середину циклов NREM – REM. Обратите внимание, что первое значение базового сна пересчитывается через 24 часа. Вертикальные линии показывают стандартную ошибку среднего. Экспоненциальная функция насыщения была аппроксимирована экстраполированными значениями в начале сна. Эта экстраполяция была основана на продолжительности цикла NREM – REM дневного сна и постоянной времени функции экспоненциального снижения, подобранной через средние значения SWA в циклах NREM – REM базового сна. Все значения выражены в процентах от значения медленной активности в первом цикле NREM – REM исходного ночного сна.Воспроизведено с разрешения Дейка.33
Сон в течение дня и, в частности, сон во второй половине дня или ранним вечером приводил к снижению уровня SWS или SWA в начальный период последующего ночного сна и к увеличению активность веретена сна по сравнению с субъектами, у которых не было дневного сна (Рисунок 5) .42 Это является следствием гомеостатической регуляции SWS / SWA: SWA, производимая во время сна ранним вечером, снижает ранее повышенное давление для SWA, с продолжительность между окончанием дневного сна и запланированным началом эпизода ночного сна недостаточна для восстановления этого давления для SWA.
Рисунок 5A : склонность ко сну, B : медленная волновая активность (SWA) и C : временной ход SWA и состояний бдительности (N, сон с небыстрым движением глаз; R, сон с быстрым движением глаз; W, бодрствование) базовой ночи, дневного сна и пост-дневного сна после дневного сна ранним вечером у одного субъекта. SWA выражали как процент от общего накопленного SWA в исходном сне. Воспроизведено с разрешения Werth et al.42
В более поздних исследованиях изучались местные аспекты регуляции SWA как у людей43–45, так и у грызунов.46,47 Во время исходного сна медленные волны наиболее распространены в лобных областях, а во время восстановительного сна после недосыпания лобная кора является областью мозга, в которой наблюдается наибольшее увеличение SWA по сравнению с центральной, теменной и затылочной областями. 35,43 Более сильное увеличение SWA после лишения сна в лобных областях мозга, вместе с преобладанием SWA в лобных областях мозга, даже на исходном уровне, было интерпретировано как доказательство роли SWS в этих областях. функции мозга, которые обычно связаны с лобной корой.Дальнейшие доказательства локальной регуляции SWS получены из экспериментов, в которых определенные переживания во время бодрствования, ведущие к активации в определенных областях мозга, приводили к локальному увеличению SWA в этих областях. Например, обширная соматосенсорная стимуляция правой руки перед сном привела к увеличению мощности дельты в левом полушарии, в частности над соматосенсорной корой.45 Совсем недавно было показано, что противоположная манипуляция — иммобилизация руки — приводит к местное сокращение SWA в сенсомоторной коре.48
Эти исследования подтверждают гипотезу о том, что механизм гомеостатической регуляции, а не механизм, основанный на циркадном ритме, является основным механизмом, участвующим в регуляции SWS и SWA. Более того, эти данные также предполагают, что процесс SWS или NREM-сна регулируется в ответ на локальную активацию областей мозга во время бодрствования.
ИНДИВИДУАЛЬНЫЕ РАЗЛИЧИЯ В SWS
Люди заметно различаются по продолжительности SWS и уровню SWA.В определенной степени демографические характеристики, включая пол и возраст, влияют на эти межличностные различия, и генетические факторы, по-видимому, также играют роль.
Гендерные различия в SWS / SWA
Гендерные различия в NREM-сне были отмечены для субъектов среднего и старшего возраста, при этом у женщин было больше SWS во время нормального и восстановительного сна, а также у 3-й и 4-й стадий NREM-сна. 50 Было высказано предположение, что это различие может быть связано с уменьшением SWS, которое происходит с возрастом, а не с конкретным гендерным различием.50
В исследовании, сравнивавшем 13 молодых мужчин (средний возраст 23,5 года) и 15 молодых женщин (средний возраст 21,9 года), визуальная оценка ЭЭГ показала, что мужчины и женщины имеют одинаковое количество МС и быстрого сна. Однако спектральный анализ обнаружил значительно более высокие плотности мощности во время медленного сна в широком диапазоне частот (0,25–11,0 Гц) у женщин по сравнению с мужчинами.51 Эти различия сохранялись на протяжении всего периода сна. Возникает вопрос, отражают ли эти различия нейроанатомические различия, а не гендерные различия в механизмах регуляции сна.51 Последующие исследования показали, что женщины имеют больше SWS и SWA, чем мужчины, и эти половые различия присутствуют у взрослых всех возрастов.52,53 Функциональная значимость и корреляты этих половых различий в SWS изучаются, но остаются малоизученными. 54,55 Понимание различий в SWS между мужчинами и женщинами представляет интерес, потому что многие женщины жалуются на проблемы со сном, такие как бессонница. Фактически, последние данные показывают, что влияние бессонницы на SWA различается у мужчин и женщин.56 Удивительно, но у женщин с первичной бессонницей была большая мощность низких частот (3-5 Гц) во всех периодах NREM, а также большая мощность высоких частот (16-44 Гц) в первые 3 периода NREM по сравнению с контрольной группой (те кто хорошо спал). Напротив, мужчины с первичной бессонницей не отличались от контрольной группы по этим параметрам.
Возрастное снижение SWS / SWA и склонности к дневному сну
SWS и SWA демонстрируют серьезные изменения на протяжении жизни3,57,58, и старение действительно является очень сильным предиктором индивидуальных различий.Фактически, по сравнению с большим количеством других параметров сна, размер влияния старения на SWS является одним из самых больших.4 SWS достигает пика в препубертатный период59 и затем снижается.58,60 Многие из этих исследований также показали. Показано, что количество ночных пробуждений увеличивается с возрастом. Следует подчеркнуть, что данные возрастные изменения МС и непрерывности сна наблюдаются даже у здоровых людей без жалоб на сон и нарушений сна. Возникает вопрос, приводит ли это снижение качества ночного сна с возрастом к негативным последствиям для дневного функционирования, т.е.г., повышение склонности ко сну. Удивительно, но доказательств этому появилось очень мало. Фактически, обзоры влияния возраста на склонность ко сну, оцененные с помощью теста множественной задержки сна (MSLT), предполагают, что склонность ко сну в дневное время снижается с возрастом.61 Недавно было показано, что пожилые люди имеют более низкую склонность ко сну в дневное время, чем молодые люди. и что это сокращение наблюдается даже при контроле за вариацией привычной продолжительности сна.53,57,62 Это возрастное снижение склонности ко сну в дневное время уже наблюдается у людей среднего возраста и происходит при наличии статистически значимых сокращений общей продолжительности сна. время, SWS и SWA.57 Связанное с возрастом снижение SWS и склонности ко сну наблюдается во всех циркадных фазах.63,64 Таким образом, в целом, пожилые люди и мужчины в частности имеют гораздо меньше SWS / SWA, чем молодые люди, и продолжительность сна, в частности продолжительность медленного сна 65 — гораздо хуже у пожилых людей.
Гены и индивидуальные различия
Люди также заметно различаются в отношении SWS и SWA даже в узком возрастном диапазоне и в пределах одного пола. Эти различия устойчивы к манипуляциям с переменными состояния, например.g., недосыпание и может рассматриваться как подобный признаку.66 Фактически, изменения в ряде генов, включая те, которые кодируют аденозиндезаминазу 67, и ген циркадных часов, кодирующих белок PERIOD3 (PER3) 68, теперь были показаны. для прогнозирования индивидуальных различий в SWS. Было показано, что у грызунов другие гены часов, которые экспрессируются не только в SCN, но и в других областях мозга, влияют на SWS.23 Таким образом, в отличие от независимых отношений на поведенческом и нейроанатомическом уровнях, SWS / SWA и циркадные ритмы. ритмичность, по-видимому, связана на молекулярном уровне.Остается установить, связаны ли эти индивидуальные различия в SWS и их генетических предикторах со сном и циркадными расстройствами. Было показано, что полиморфизм PER3, который предсказывает различия в SWS, связан с синдромом задержки фазы сна.69
ФУНКЦИОНАЛЬНЫЕ КОРРЕЛЯЦИИ SWS
Большие межиндивидуальные различия в SWS / SWA, глубокие возрастные изменения в SWS / SWA и точное гомеостатическое регулирование SWS / SWA поднимают вопрос о функциональных коррелятах SWS / SWA.Могут ли различия в SWS / SWA быть связаны с различиями в качестве сна и дневной активности?
Влияние снижения ночного SWS посредством дневного сна на последующую склонность ко сну
Изменяя давление сна, используя либо исследования дневного сна для снижения давления сна, либо исследования депривации сна для повышения давления сна, можно повлиять на склонность ко сну в ночное время. Например, в исследовании с участием девяти здоровых субъектов дневной сон ранним вечером приводил к заметному увеличению латентности начала сна и сокращению латентности быстрого сна во время последующего ночного сна.Таким образом, латентность (среднее значение ± стандартная ошибка) до стадии 1 увеличилась с 8,8 (± 1,6) минут до 35,6 (± 8,8) минут, а латентность до стадии 2 с 12,8 (± 2,0) минут до 45,5 (± 8,8 минут), в то время как латентность быстрого сна была уменьшена с 76,5 (± 4,3) минут до 57,3 (± 7,8) минут (Рисунок 5) .42 Эти эффекты дневного сна настолько существенны и надежны, что послеродовой ночной сон был использован в качестве модели. для бессонницы во сне — в ряде исследований.70,71
Меж- и внутрииндивидуальные различия в SWS и непрерывности сна
SWS и непрерывность сна ухудшаются на протяжении всей жизни.4 Депривация сна усиливает МС и снижает возбуждение, в частности, микровозбуждение.72 Взаимосвязь между МС и непрерывностью сна была дополнительно исследована на большой группе здоровых субъектов, состоящей из молодых (20–30 лет; N = 41), среднего возраста. люди пожилого возраста (40–55 лет; N = 31) и пожилые (66–83 года; N = 31). Было замечено, что SWA и SWS отрицательно коррелировали с количеством пробуждений. Эта корреляция сохранялась даже при учете возраста.3 В целом эти данные предполагают, что индивидуальные вариации в SWS / SWA и непрерывности сна взаимосвязаны.Эта взаимосвязь также наблюдалась в исследованиях на животных; количество кратковременных пробуждений обратно коррелирует с SWA у крыс, 73 морских свинок, 74 и мышей.75
SWS Нарушение дневного функционирования
Для исследования функционального значения SWS необходимы экспериментальные подходы. Один из таких подходов основан на нарушении и уменьшении SWS посредством акустической стимуляции (или умеренной электрической стимуляции в ранних исследованиях). Такие исследования продемонстрировали, что достигнутое количество SWS может быть существенно уменьшено без значительных изменений во времени NREM и REM.15,76,77 Другими словами, вмешательство только уменьшает SWS и, фактически, в первую очередь уменьшает сон стадии 4, хотя акустическая стимуляция приведет к пробуждению и, возможно, к кратковременному пробуждению. Эти эксперименты также продемонстрировали, что депривация SWS приводит к восстановлению SWS во время ненарушенного последующего сна, который может происходить либо в том же эпизоде сна, либо во время следующего эпизода сна, в ситуациях, когда нарушение SWS длилось всю ночь.78,79 Например, В исследовании нарушения сна с участием девяти здоровых молодых мужчин испытуемых лишали МСВ с помощью акустических стимулов (избегая пробуждения) в течение первых 3 часов третьей ночи.Частота дельта- и тета-активности уменьшалась во время депривации SWS. В часы сна после депривации SWS как плотность мощности, так и количество SWS увеличивались по сравнению с базовой ночью. Это исследование демонстрирует точную гомеостатическую регуляцию SWS во время эпизода сна (рис. 6) .78
Рис. 6Влияние повторного нарушения сна на медленную волновую активность (SWA) у девяти здоровых субъектов. A : Динамика SWA во время исходного сна (вверху) и во время эпизода сна, во время которого SWA подавлялась акустической стимуляцией (внизу).33 B : Накопление энергии электроэнцефалограммы (ЭЭГ) во время исходного уровня (сплошная линия) и эпизода сна, во время которого SWA подавлялась акустической стимуляцией (пунктирная линия). Сплошные вертикальные линии указывают стандартные ошибки средних значений. Пунктирная вертикальная линия указывает конец депривации медленного сна в течение экспериментальной ночи. Значения выражены в процентах от энергии, накопленной в течение первых 7 часов исходной ночи. 78 NREM, небыстрое движение глаз; REM, быстрое движение глаз.Воспроизведено с разрешения Dijk33 и Dijk et al.78
Аналогичным образом, после 2 ночей нарушенного сна с подавлением акустической стимуляции SWS у 10 здоровых субъектов, доля SWS увеличивалась в ночь восстановления. Похоже, что дефицит SWS в первую ночь был перенесен на ночь восстановления.79 Эти результаты подтверждают гипотезу о том, что количество SWS более сильно связано с количеством SWS и SWA, которое было получено во время предыдущих периодов сна, чем к общему времени сна, что снова свидетельствует о запоминании SWS в соответствии с его гомеостатической регуляцией.Эти данные также подразумевают, что депривация SWS приводит к дефициту SWS, который переносится на период бодрствования, что позволяет предположить, что этот подход может быть использован для исследования влияния дефицита SWS на функцию бодрствования. Был проведен ряд таких исследований, и результаты оказались неоднозначными. В раннем эксперименте Джонсон и его коллеги сравнили 3 ночи депривации 4 стадии у 7 молодых людей и 3 ночи депривации REM у 7 молодых людей. В то время как обе процедуры депривации привели к уменьшению ряда показателей, в том числе производительности подсчета, существенных различий между двумя режимами депривации не было обнаружено, и конкретный вклад 4-й стадии сна в дневную функцию не был выявлен.80 В более поздних экспериментах сообщалось о влиянии депривации SWS на работоспособность и склонность ко сну в дневное время, но эти эффекты были отнесены к негативным эффектам вмешательства на непрерывность сна, а не конкретно к SWS.81,82 Ввиду тесной взаимосвязи между SWS и непрерывность сна, однако, остается спорным, можно ли разделить эффекты SWS и непрерывности сна. Во многих из этих исследований использовались только небольшие размеры выборки и не использовался спектральный анализ для количественной оценки эффектов депривации SWS.
В одном недавнем эксперименте здоровые испытуемые (N = 58) были лишены SWS в течение 2 ночей с использованием акустической стимуляции при появлении дельта-волн в ЭЭГ во сне. Измерение склонности к дневному сну с помощью MSLT продемонстрировало значительное увеличение склонности к дневному сну после обеих ночей депривации по сравнению с контрольной группой, которая не была лишена SWS (N = 59) .83 В другом недавнем эксперименте депривация SWS использовалась для исследования его вклад в зависящее от сна улучшение задачи обучения восприятию у 20 молодых людей.Улучшение производительности в задаче распознавания текстуры коррелировало с SWA, когда контрольные и экспериментальные условия были объединены у тех 16 человек, которые соответствовали определенному исходному критерию производительности.84 Аналогичная корреляция между локальной SWA и улучшением производительности в задаче на зрительно-моторную координацию сообщалось с использованием ЭЭГ высокой плотности.44
В целом эти исследования показывают, что склонность ко сну в дневное время увеличивается за счет увеличения давления на SWS из-за нарушения SWA в ночное время, в то время как склонность к ночному сну и SWA снижаются после вечернего сна во время что большая часть давления для SWA рассеивается.Эти исследования, вместе с наблюдением, что SWS и SWA связаны с непрерывностью сна (оцениваемой по количеству пробуждений), предполагают, что усиление SWS и SWA может привести к улучшению непрерывности сна3, а также к снижению склонности ко сну в дневное время. и может способствовать дневной функции.
ГИПНОТИКИ
Большинство снотворных, в частности бензодиазепины и Z-препараты, такие как золпидем и зопиклон, уменьшают SWS и SWA.85 Однако ряд соединений, обладающих снотворными свойствами, может усиливать SWS; к ним относятся этанол, 86 гамма-гидроксибутират / оксибат натрия, 87 тиагабин, 88 габоксадол, 89 и антагонисты рецепторов 5HT 2 .70 Например, при прямом сравнении сегансерина, антагониста 5HT 2 , и темазепама на стадиях сна человека и спектрах мощности ЭЭГ в течение ночи после дневного сна ранним вечером было обнаружено, что сегансерин вызывает увеличение SWS и увеличение плотности мощности на дельта- и тета-частотах во время медленного сна. Напротив, темазепам вызывал снижение плотности мощности на частотах дельта и тета.70 Таким образом, антагонизм 5HT 2 увеличивал SWA, а бензодиазепины оказывали противоположный эффект.
Кроме того, эпливансерин, антагонист серотониновых рецепторов 2А (ASTAR), при введении здоровым субъектам утром или вечером удваивал продолжительность SWS и приводил к соответствующему снижению фазы сна 2 по сравнению с плацебо. Анализ эффектов эпливансерина на ЭЭГ сна показал, что SWA увеличилась, а частотная активность веретена снизилась.91 Эти данные показывают, что антагонизм рецепторов 5HT 2 приводит к усилению SWS и SWA во время сна, что может привести к улучшение структуры сна и может принести пользу пациентам с первичной бессонницей.
ВЫВОДЫ
SWS точно регулируется и компенсируется. Преобладание SWS в лобных областях, связанных с более высокой функцией мозга, или в областях, которые были очень активными во время бодрствования, подчеркивает значительную роль SWS. Отрицательная корреляция между SWA и SWS и показателями непрерывности сна у животных и людей позволяет предположить, что SWS способствует непрерывности сна. Экспериментальное нарушение SWS увеличивает неглубокий сон и фрагментацию сна, увеличивает склонность ко сну в дневное время и может нарушать дневную функцию.Эти данные служат обоснованием для проверки гипотезы о том, что фармакологическое усиление SWS может привести к улучшению поддержания сна и дневной функции у пациентов с первичной бессонницей или не восстанавливающим сном.
ЗАЯВЛЕНИЕ О РАСКРЫТИИ ИНФОРМАЦИИ
Профессор Дейк получил исследовательскую поддержку от AFOSR, BBSRC, GlaxoSmithKline, H Lundbeck A / S, Merck & Co Inc., Philips Lighting, Organon, Takeda и Wellcome Trust. Профессор Дейк работал консультантом в компаниях Actelion, Cephalon, GlaxoSmithKline, H Lundbeck A / S, Merck & Co Inc., Pfizer Inc., Philips Lighting, Sanofi-Aventis и Takeda.
СПИСОК ЛИТЕРАТУРЫ
1 Капот MHS Восстановление сна как функция периодического пробуждения, движения или электроэнцефалографических изменений Сон 19871036473, 3659734
2 Акерстедт THume KMinors Sleepological Sleephouse 9 Характеристики времени сна и физиологического режима сна JG Res 199762219, 9493521
3 Dijk DJGroeger JDeacon SStanley NA Связь между индивидуальными различиями в медленноволновом сне, медленноволновой активности и непрерывности сна у мужчин и женщин молодого, среднего и старшего возраста Eur Neuropsychopharmacol 200616S538 4000367
67 90 Охайон М.М. Карскадон М.А. Guilleminault CVitiello MV Мета-анализ количественных параметров сна от детства до старости у здоровых людей: разработка нормативных значений сна на протяжении всей жизни человека Сон 200427125573, 155867795 Rechtschaffen AKales A A методы и система оценки стадий сна людей 1968 Вашингтон, Служба здравоохранения округа Колумбия, Типография правительства США
6 Iber CAncoli-Israel SChesson AQuan SF Руководство AASM по оценке сна и связанных с ним событий: правила, терминология и технические характеристики 2007 Вестчестер, штат Иллинойс, Американская академия медицины сна
7 Дейк Д.Хайес BCzeisler CДинамика электроэнцефалографических веретен сна и медленноволновой активности у мужчин: эффект депривации сна Brain Res 19936261909, 8281430
WCBrain Электрическая активность и сенсорная обработка во время бодрствования и сна Принципы и практика медицины сна 2005PhiladelphiaW.B. Saunders Co.1051249 Knoblauch VMartens WWirz-Justice AKrauchi KCajochen C Региональные различия в циркадной модуляции характеристик веретена сна человека Eur J Neurosci 20031815563, 12859349
DPD- человека гомеостаз сна на основе медленноволновой активности ЭЭГ: количественное сравнение данных и моделирования Brain Res Bull 19933197113, 8453498
11 Esser SKHill SLTononi G Гомеостаз сна и корковая синхронизация: I.Моделирование влияния синаптической силы на медленные волны сна Сон 200730161730, 18246972
12 Ван Каутер Элатта Ф. Недельчева А. и др. Взаимные взаимодействия между осью гормона роста и сном Горм роста IGF Res 200414Suppl AS5
13 Brandenberger GEhrhart JPiquard FSimon C Обратная связь между ультрадианными колебаниями дельта-волновой активности и вариабельностью сердечного ритма во время сна циркадная фаза Am J Physiol Heart Circ Physiol 2008295h3156h3163, 18835917
15 Tasali ELeproult REhrmann DAVan Cauter ES Медленноволновой сон и риск диабета 2 типа у людей Proc Natl Acad Sci10 0003 16 Дейк Д.Д. Медленный сон, диабет, а и симпатическая нервная система Proc Natl Acad Sci USA 200810511078, 18212114
17 Dijk DJvon Schantz MTiming и объединение человеческого сна, бодрствования и производительности симфонией осцилляторов J Biol Rhythms
18 Эдгар DMDement WCFuller CA Влияние поражений SCN на сон у беличьих обезьян: доказательства противостоящих процессов в регуляции сна и бодрствования J Neurosci 199313106579, 8441003
19 Mistlberger REC Циркадная регуляция сна у млекопитающих Brain Res Brain Res Rev 20054942954, 16269313
20 Cohen RAAlbers HE Нарушение циркадной и когнитивной регуляции у человека после дискретного поражения гипоталамуса: тематическое исследование Neurology 1991417269, 2027490
- циркадный порядок и расстройство у млекопитающих r: значение для физиологии и болезни Nat Rev Genet 2008976475, 18802415
22 Archer SNViola AUKyriakopoulou Vvon Schantz MDijk DJ Межличностные различия в привычном времени сна и увлеченной фазе эндогенных циркадных ритмов в BMALER1 человека и PERRNA человека. лейкоциты Сон 20083160817, 18517031
23 Franken PDijk DJA нециркадная роль часовых генов в гомеостазе сна Eur J Neurosci 2009In press
MC36725 Zulley JWever RAschoff J Зависимость начала и продолжительности сна от циркадного ритма ректальной температуры Pflugers Arch 19813913 Dawn, 7312563 9000an SB DGBorbely AATiming человеческого сна: процесс восстановления ga с помощью циркадного кардиостимулятора Am J Physiol 1984246R161R183, 6696142
27 Dijk DJCzeisler CAC Влияние циркадного кардиостимулятора и гомеостата сна на склонность ко сну, структуру сна, электроэнцефалографические медленные волны и активность веретена J у людей 199515352638, 7751928
28 Wyatt JKRitz-De Cecco ACzeisler CADijk DJ Циркадные ритмы температуры и мелатонина, сон и нейроповеденческая функция у людей, живущих в течение 20-часового дня Am J Physiol 1999277R111652R1163 ECzeisler CADijk DJ Аттенуированная амплитуда циркадной и зависимой от сна модуляции электроэнцефалографических характеристик веретена сна у пожилых людей Neurosci Lett 19992602932, 10027692
30 Dijk DJShanahan TLDuffy JFRonda JMC движение глаз сон с фазой циркадного ритма мелатонина у людей J Physiol 19975058518, 9457658
31 Driver HSDijk DJWerth EBiedermann KBorbely AAS Сон и электроэнцефалограмма сна во время менструального цикла у молодых здоровых женщин
32 Webb WBAgnew HWStage 4 сон: влияние переменных динамики времени Science 197117413546, 5135723
33 Dijk DJEEG медленные волны и шпиндели сна: окна на спящем мозгу Behav Brain Res 7537302
34 Borbely AAAchermann PSГомеостаз сна и модели регуляции сна J Biol Rhythms 19991455768, 10643753
35 Finelli LABorbely AAAchermann P Функциональная топография электроэнцефалограммы сна человека без REM
0, 9000sc2000 9000sc2000 9000sc4000 DJBrunner DP Borbely A.A. Измерение плотности мощности ЭЭГ во время длительного сна у человека Am J Physiol 19R650R661, 231671237 Franken PTobler IBorbely AA Продолжительный фотопериод у лабораторных крыс: глубокое влияние на суточный сон, но не на гомеостаз сна Am J Physiol 1995269R691R701, 7573572
38 Dijk DJDaan SS Спектральный анализ ЭЭГ сна у дневных грызунов: Eutamias sibiricus J Comp Physiol 198916520515, 2746549
ar во время монофазного и двухфазного сна человека Neurosci Lett 2001298836, 1116328340 Брюнет DNish DMacLean AWCoulter MKnowles JB Временной ход «процесса S»: сравнение визуально оцениваемых медленных волн сна и спектрального анализа мощности
7 2458235
41 Dijk DJBeersma DGDaan Плотность мощности SEEG во время дневного сна: отражение песочных часов, измеряющих продолжительность предыдущего бодрствования J Biol Rhythms 1987220719, 2979661
42 Werth EDijk DJAchermann PBorbely AADynamics после раннего вечернего сна EEG данные и моделирование Am J Physiol 1996271R501R510, 8853369
43 Cajochen CFoy RDijk DJ Фронтальное преобладание относительного увеличения дельты сна и тета-активности ЭЭГ у людей после потери сна Sleep Res Online 19992659, 11382 Huber RGhilardi MFMassimini MTononi GLокальный сон и обучение Nature 20044307881, 15184907
45 Kattler HDijk DJBorbely AA Эффект односторонней соматосенсорной стимуляции перед сном на сон на сон
VVTobler I Региональные различия в муфтах NREM p медленноволновая активность у мышей с врожденной каллозальной дисгенезией J Sleep Res 200514299304, 1612010547 Вязовский В.В., Велкер Е.Фричи Дж.М.Тоблер I Региональный паттерн метаболической активации отражается на ЭЭГ сна 9000 в сочетании с односторонней депривацией сна у мышей. Eur J Neurosci 200420136370, 15341608
48 Huber RGhilardi MFMassimini M, et al.Иммобилизация руки вызывает кортикальные пластические изменения и местно снижает медленную активность сна Nat Neurosci 200676, 16936722
49 Reynolds CFKupfer DJHoch CC и др. Депривация сна у здоровых пожилых мужчин и женщин: влияние на настроение и сон во время восстановления Сон 19869492501, 3809863
50 Webb WBS Сон у пожилых людей: структура сна мужчин и женщин в возрасте от 50 до 60 лет J Gerontol 1982375816, 7096931
51 Dijk DJBeersma ЭЭГ сна молодых людей: визуальная оценка и спектральный анализ Сон 19807, 2595173
52 Carrier JLand SBuysse DJKupfer DJMonk TH Влияние возраста и пола на спектральную плотность мощности ЭЭГ сна в средние годы жизни (возраст 20-60 лет) Психофизиология 20013823242, 11347869
53 Dijk DJ Сон стареющих женщин и мужчин: назад к основам cs Sleep 200629123, 16453973
54 Latta FLeproult RTasali EHofmann EVan Cauter ESex Различия в дельта- и альфа-активности ЭЭГ у здоровых пожилых людей Sleep 200528152534, 16408411
- Latta Eeplt al.Половые различия в ночной секреции гормона роста и пролактина у здоровых пожилых людей: взаимосвязь с переменными ЭЭГ сна Сон 200528151924, 16408410
56 Buysse DJGermain AHall ML и др. Спектральный анализ ЭЭГ при первичной бессоннице: эффекты периода NREM и пол различия Sleep 200831167382, 190
57 Dijk DJGroeger JDeacon SStanley Связанное с NAge снижение склонности ко сну в дневное время J Sleep Res 200615Suppl.1183, 16704574
58 Landolt HPDijk DJAchermann PBorbely AA Влияние возраста на ЭЭГ сна: медленноволновая активность и частотная активность веретена у мужчин молодого и среднего возраста анализ электроэнцефалограммы сна в подростковом возрасте Сон 20042777483, 15283014
60 Van Cauter ELeproult RPlat Изменения медленноволнового сна и быстрого сна, связанные с изменениями медленноволнового сна и быстрого сна, а также взаимосвязь с уровнями гормона роста и кортизола у здоровых мужчин JAMA 20002848618,
61 Arand DBonnet MHurwitz T, et al.Клиническое использование MSLT и MWT Sleep 20052812344, 15700728
62 Klerman EBDijk DJ Снижение максимальной способности к засыпанию, связанное с возрастом — последствия для бессонницы Curr Biol 200818111823, 18656358 7 7 THCarrier JBegley AC Циркадные паттерны сна, сонливости и работоспособности у пожилых и молодых людей Сон 200528136576, 16335481
64 Дийк Д.Д. , 10841208
65 Dijk DJDuffy JFCzeisler Увеличение пробуждений, связанное с возрастом: нарушение консолидации медленного сна на всех циркадных фазах молодежь J Sleep Res 20071617080, 17542947
67 Retey JVAdam MHonegger E, et al.Функциональная генетическая вариация аденозиндезаминазы влияет на продолжительность и интенсивность глубокого сна у людей Proc Natl Acad Sci USA 20051021567681, 16221767
68 Виола AUArcher SNJames LM и др. Полиморфизм Per3 предсказывает структуру сна и способность бодрствования Biol 2007176138, 17346965
69 Archer SNRobilliard DLSkene DJ, et al. Полиморфизм длины гена циркадных часов Per3 связан с синдромом отложенной фазы сна и крайним дневным предпочтением Sleep 2003264135, 128413 703
9037k DJBeersma DGDaan SVan den Hoofdakker RHE Влияние сегансерина, антагониста 5-HT2, и темазепама на стадии сна человека и спектры мощности ЭЭГ Eur J Pharmacol 198917120718, 257600071 Mathias SSteiger AL качество сна после дремоты Психофармакология (Berl) 2001157299304, 11605086
72 Sforza EChapotot FPigeau RPaul PNBuguet A Влияние депривации сна на спонтанные возбуждения у людей Sleep 200427106875, 15532200
73 Franken PDijk DJTobler IBorbely EAS Spectra, депривация температурных состояний, эмоциональная депривация состояния сна Am J Physiol 19R198R208, 1858947
74 Tobler IFranken PJaggi K Состояния бдительности, спектры ЭЭГ и температура коры головного мозга у морских свинок Am J Physiol 1993264R1125R113673, 83229265 Регуляция сна Tobler 932292 нормальные мыши и мыши с дефицитом прионного белка J Neurosci 199717186979,
45
76 Agnew HWJWebb WBWilliams RL Эффекты депривации сна на четвертой стадии Electroencephalogr Clin Neurophysiol 196417683000 196417683 Мощность ЭЭГ Плотность сна и продолжительность спонтанного сна Электроэнцефалор Clin Neurophysiol 19897231220, 2467797
78 Дейк DJBeersma DGDaan SBloem GMVan den Hoofdakker RH Количественный анализ влияния медленной депривации мощности сна во время последующих 3 часов ЭЭГ на первые 3 часа ЭЭГ. Arch Psychiatry Neurol Sci 19872363238, 3678290
79 Ferrara MDe Gennaro LBertini MS Избирательная депривация медленноволнового сна (SWS) и восстановление SWS: нужно ли нам фиксированное количество SWS за ночь? Sleep Res Online 19992159, 11382878
80 Johnson LC Naitoh PMoses JMLubin A Взаимодействие депривации REM и депривации стадии 4 с полной потерей сна: эксперимент 2 Психофизиология 19741114759, 43HP34
03
19741114759, 43HPO382 Уолш Дж. К. Хартман PGSchweitzer PKS Медленноволновая депривация сна и функция пробуждения J Sleep Res 199431625, 1060 SS7104
депривация связана с повышенной склонностью ко сну в дневное время у мужчин и женщин молодого, среднего и старшего возраста. J Sleep Res 200615Suppl.1250, 16684 Aeschbach DCutler AJRonda JMA Роль гомеостаза сна с небыстрыми движениями глаз в перцептивном обучении J Neurosci 200828276672, 18337406
85 Lancel MRole рецепторов в начальной стадии регуляции GABAA реакции сна на периферически вводимые модуляторы и агонисты Сон 1999223342, 9989364
86 Дейк DJBrunner DPAeschbach DTobler IBorbely AAT Влияние этанола на спектр мощности ЭЭГ человека во сне отличается от такового у бензодиазепиновых рецепторов 967300030003000 1300030002 9675272 272
87 Pardi DBlack Jgamma-гидроксибутират / оксибат натрия: нейробиология и влияние на сон и бодрствование Препараты для ЦНС 2006209931018, 17140279
88 Mathias SWetter TCSteiger ALancelabake M Старение 20012224753, 11182474
89 Walsh JK Дикон SDijk DJLundahl J Селективный внесинаптический агонист GABAA, габоксадол, улучшает традиционные меры гипнотической эффективности и усиливает медленную волновую активность в модели временной бессонницы 52 9602 9602 9602 5
AMartinez JMPinquier JL Сон и психомоторные показатели у здоровых субъектов после утреннего или вечернего приема эпливансерина, нового соединения для сна J Sleep Res 200817Suppl.1Abstract No. P35991 Landolt HPMeier VBurgess HJ, et al. Рецепторы серотонина-2 и человеческий сон: влияние селективного антагониста на спектры мощности ЭЭГ Нейропсихофармакология 19992145566, 10457543
сна 909
24 Cheisler CA Weitzman E. и организация зависят от его циркадной фазы Science 198021012647, 7434029
03
нарушение и депривация медленного сна Physiol Behav 1986379158, 3786485Если вы считаете, что контент, доступный через Веб-сайт (как определено в наших Условиях обслуживания), нарушает или другие ваши авторские права, сообщите нам, отправив письменное уведомление («Уведомление о нарушении»), содержащее в информацию, описанную ниже, назначенному ниже агенту.Если репетиторы университета предпримут действия в ответ на ан Уведомление о нарушении, оно предпримет добросовестную попытку связаться со стороной, которая предоставила такой контент средствами самого последнего адреса электронной почты, если таковой имеется, предоставленного такой стороной Varsity Tutors.
Ваше Уведомление о нарушении прав может быть отправлено стороне, предоставившей доступ к контенту, или третьим лицам, таким как в качестве ChillingEffects.org.
Обратите внимание, что вы будете нести ответственность за ущерб (включая расходы и гонорары адвокатам), если вы существенно искажать информацию о том, что продукт или действие нарушает ваши авторские права.Таким образом, если вы не уверены, что контент находится на Веб-сайте или по ссылке с него нарушает ваши авторские права, вам следует сначала обратиться к юристу.
Чтобы отправить уведомление, выполните следующие действия:
Вы должны включить следующее:
Физическая или электронная подпись правообладателя или лица, уполномоченного действовать от их имени; Идентификация авторских прав, которые, как утверждается, были нарушены; Описание характера и точного местонахождения контента, который, по вашему мнению, нарушает ваши авторские права, в \ достаточно подробностей, чтобы позволить репетиторам университетских школ найти и точно идентифицировать этот контент; например нам требуется а ссылка на конкретный вопрос (а не только на название вопроса), который содержит содержание и описание к какой конкретной части вопроса — изображению, ссылке, тексту и т. д. — относится ваша жалоба; Ваше имя, адрес, номер телефона и адрес электронной почты; а также Ваше заявление: (а) вы добросовестно считаете, что использование контента, который, по вашему мнению, нарушает ваши авторские права не разрешены законом, владельцем авторских прав или его агентом; (б) что все информация, содержащаяся в вашем Уведомлении о нарушении, является точной, и (c) под страхом наказания за лжесвидетельство, что вы либо владелец авторских прав, либо лицо, уполномоченное действовать от их имени.
Отправьте жалобу нашему уполномоченному агенту по адресу:
Чарльз Кон
Varsity Tutors LLC
101 S. Hanley Rd, Suite 300
St. Louis, MO 63105
Или заполните форму ниже:
Пять этапов, через которые проходит ваше тело во время сна
Циклы сна: наш мозг и тело во сне
На протяжении большей части истории человечества считалось, что мозг отключается во время сна, но теперь ученые знают, что он довольно активен.Эксперты делят сон на две основные фазы: медленный сон и быстрый сон. REM означает быстрое движение глаз, потому что на этом этапе глаза быстро перемещаются во всех направлениях. Напротив, во время медленного сна движение глаз очень мало.
Каждая из пяти стадий сна характеризуется определенными мозговыми волнами. Этапы проходят несколько раз в течение ночи. «Это почти как если бы это была эта прекрасная симфония, и вы слышите оркестр, играющий на разных инструментах или играющий в разное время через некоторое повторение», — сказал д-р.Ана Кригер, которая специализируется на медицине сна в Weill Cornell Medicine в Нью-Йорке.
БЫСТРЫЙ СОН
ЭТАП 1
Стадия 1 сна — это переходная стадия между бодрствованием и сном. На этом этапе и ум, и тело начинают замедляться. Мышцы расслабляются, и мозг начинает производить альфа- и тета-волны более низкой частоты, которые помогают нам успокоиться. Мы можем испытывать мышечные спазмы или ощущение падения, но при этом легко просыпаемся.Этап 1 обычно длится менее 5 минут.
ЭТАП 2
На стадии 2 мозговая волна продолжает замедляться со вспышками быстрой мозговой активности, называемыми веретенами сна и K-комплексами. Считается, что эти всплески играют роль в консолидации долговременной памяти и сенсорной обработке, а также защищают мозг от пробуждения ото сна. Снижается температура тела и замедляется пульс. Лучше всего просыпаться после сна второй стадии, прежде чем переходить к более глубоким стадиям, что происходит примерно через 20-30 минут после засыпания.
ЭТАПЫ 3 и 4
Стадия 3 и 4 сна, наиболее восстановительные стадии, известны как глубокий сон. Нам нужно от полутора до двух часов глубокого сна в сутки. На стадии 3 очень медленные мозговые волны, называемые дельта-волнами, чередуются с более мелкими и более быстрыми волнами. На стадии 4 мозг почти исключительно производит дельта-волны. Дельта-волны способствуют развитию иммунной функции и помогают нам чувствовать себя восстановленными. Во время глубокого сна тело восстанавливает мышцы и ткани, стимулирует рост и повышает иммунную функцию.На этих этапах вы не почувствуете движения глаз или мышечной активности. На стадиях 3 и 4 возникают разговоры во сне и ночные кошмары. Лунатизм также проявляется в глубоком сне. Лунатизм может быть опасным, потому что он может случайно пораниться, упав или коснувшись острого предмета.
БЫСТРЫЙ СОН
Быстрый сон, опять же, означает быстрое движение глаз, потому что глаза быстро бегают во все стороны. Именно на этой стадии происходит большая часть сновидений.Мозг становится более активным, частота сердечных сокращений и артериальное давление повышаются, а дыхание становится частым и нерегулярным. Быстрый сон также играет роль в обучении; во время быстрого сна мозг объединяет и обрабатывает информацию, которая сохраняется в долговременной памяти.
По данным Американской ассоциации сна, взрослые тратят около 50% общего времени сна на стадии 2, около 20% — на фазу быстрого сна, а оставшиеся 30% — на других стадиях.
Первый цикл сна за ночь занимает около полутора часов, а остальные циклы в среднем от часа сорока минут до двух часов.С наступлением ночи периоды быстрого сна удлиняются, а периоды глубокого сна — короче. Взрослым необходимо спать от четырех до пяти циклов за ночь, что составляет от 6 до 9 часов сна.
Пока все это происходит, недавно открытый процесс, называемый глимфатической системой, на самом деле очищает мозг от токсинов. Ученые все еще работают над более полным пониманием того, что происходит в нашем мозгу и теле, когда мы спим. «Мы просто царапаем поверхность …. Чем больше мы понимаем, тем больше мы знаем, что на самом деле мы не знаем всех деталей », — сказал Кригер.
Чтобы узнать больше о важности сна и о том, как мы все можем получить от него больше, посмотрите полный выпуск нашей серии о сне на YourHealthExplored.com.
возрастных изменений медленного сна и быстрого сна и взаимосвязь с уровнями гормона роста и кортизола у здоровых мужчин | Медицина сна | JAMA
Контекст У молодых людей сон влияет на регуляцию гормона роста (GH) и кортизол.Взаимосвязь между снижением качества сна у пожилых людей и возрастные изменения в регуляции гормона роста и кортизола неизвестны.
Цель Определить хронологию возрастных изменений продолжительности сна и качество (стадии сна) у здоровых мужчин и сопутствующие изменения возникают на уровне гормона роста и кортизола.
Дизайн и установка Данные, полученные в результате серии исследований, проведенных в период с 1985 по 1999 гг. в 4 лабораториях.
Субъекты Всего 149 здоровых мужчин в возрасте от 16 до 83 лет со средним (SD) телом индекс массы 24,1 (2,3) кг / м 2 , без жалоб на сон и анамнеза эндокринных, психических расстройств или нарушений сна.
Основные показатели результатов Круглосуточные профили уровней гормона роста и кортизола в плазме и полиграфические записи сна.
Результаты Средний (SEM) процент глубокого медленного сна снизился с 18,9% (1.3%) в раннем взрослом возрасте (возраст 16-25 лет) до 3,4% (1,0%) в среднем возрасте (возраст 36-50 лет) и был заменен более легким сном (1 и 2 этапы) без значительное увеличение фрагментации сна или уменьшение быстрого движения глаз (БДГ) сон. Переход от среднего возраста к позднему (возраст 71-83 года) включал нет дальнейшего значительного уменьшения медленноволнового сна, но увеличение времени бодрствование продолжительностью 28 минут за декаду за счет уменьшения как легкой, не относящейся к быстрой фазе сна сон (-24 минуты на декаду; P <.001) и Быстрый сон (-10 минут на декаду; P <0,001). Снижение медленного сна с раннего взрослого до среднего возраста происходило параллельно. за счет значительного снижения секреции GH (-372 мкг за десятилетие; P <0,001). От среднего до позднего возраста, дальнейшая секреция GH снижались более медленными темпами (-43 мкг за десятилетие; P <0,02). Независимо от возраста, количество секреции GH было значительным. связаны с медленным сном ( P <0,001). Увеличение возраст был связан с повышением вечернего уровня кортизола (+19.3 нмоль / л за десятилетие; P <.001), которые стали значимыми только после 50 лет, когда сон стал более фрагментированным, а быстрый сон уменьшился. Тенденция к ассоциации между меньшим количеством быстрого сна и более насыщенным вечером Концентрация кортизола не зависела от возраста ( P <0,10).
Выводы У мужчин возрастные изменения медленного сна и быстрого сна происходят с заметно различаются по хронологии и связаны с определенными гормональными переделки.В будущих исследованиях следует оценить, есть ли стратегии по улучшению качество сна может иметь положительные гормональные эффекты.
Пониженное субъективное качество сна — одна из наиболее распространенных проблем со здоровьем. жалобы пожилых людей. 1 Самый стабильный изменения, связанные с нормальным старением, включают увеличение количества и продолжительности пробуждений и уменьшения количества глубокого медленного сна (т.е. 3 и 4 сна с медленным движением глаз (не-REM)). 2 -4 Быстрый сон, по-видимому, относительно лучше сохраняется во время старения. 3 -7 Возраст, в котором появляются изменения в количестве и распределении стадий сна. неясно, потому что большинство исследований основано на сравнении молодых и пожилых людей. Несколько исследователей заметили, что есть заметное уменьшение сна в раннем взрослом возрасте у мужчин, но не у женщин. 8 -11
Сон является основным модулятором эндокринной функции, особенно гипофизарно-зависимой. гормональный выброс.Секреция гормона роста (GH) стимулируется во время сна а у мужчин от 60% до 70% суточной секреции GH происходит во время раннего сна, в ассоциация со сном SW. 12 Уменьшает ли в SW-снах способствуют известному снижению секреции GH в нормальном состоянии. старение не известно. 13 -15
В отличие от повышенной активности оси GH во время сна, ось гипоталамус-гипофиз-надпочечники (HPA) резко ингибируется на ранних этапах SW спать. 16 -20 Более того, даже частичное недосыпание приводит к повышению уровня кортизола. выравнивает на следующий вечер. 21 Таким образом, оба уменьшение сна и недосыпание в результате повышенной фрагментации сна может способствовать повышению уровня кортизола. Повышение вечернего кортизола уровни — признак старения 14 , 15,22 считается, что это отражает нарушение контроля отрицательной обратной связи оси HPA и может лежать в основе совокупности метаболических и когнитивных переделки. 23 -25
Настоящее исследование определяет хронологию возрастных изменений сна. продолжительность и качество (то есть количество стадий сна), секреция гормона роста и кортизол уровней у здоровых мужчин и исследует, является ли снижение качества сна связаны с изменениями уровня гормона роста и кортизола.
Представлены данные 149 здоровых мужчин в возрасте от 16 до 83 лет.Средний (SD) индекс массы тела (ИМТ) испытуемых составил 24,1 (2,3) кг / м 2 . Все испытуемые имели нормальный вес (ИМТ 18-28 кг / м 2 ).
Данные были собраны в период с 1985 по 1999 гг. В ходе серии исследований. из нашей группы (109 из 149 индивидуальных наборов данных 14 , 26 -32 ) и 3 другие лаборатории, использующие аналогичные процедуры анализа, процедуры регистрации, или оба (Питтсбургский университет, Питтсбург, Пенсильвания, 14 предметов 33 , 34 ; Калифорнийский университет в Лос-Анджелесе, 8 предметов 35 , 36 ; и Государственный университет Пенсильвании, Херши, 18 предметов ( 37 ).За исключением данных от 29 субъектов, изученных в нашей лаборатории, все остальные данные были включены в ранее опубликованные отчеты, в которых не учитывалась хронология возрастных эффектов. Необработанные данные от всех 149 субъектов были отправлены в новый анализ, предназначенный для количественного определения сна и гормональных параметров совершеннолетие.
Испытуемые были платными добровольцами, у которых не было жалоб на сон, не принимали какие-либо лекарства, находились в хорошем состоянии здоровья на основании медицинского осмотра и имели отсутствие в анамнезе эндокринных, психических расстройств или нарушений сна.Вахтовые рабочие, подданные недавно путешествовали по часовым поясам, и конкурентоспособные спортсмены Исключенный. Юные испытуемые достигли пятой стадии половой зрелости по Таннеру. и полный рост. Старшие мужчины были самодостаточны и не обладали познавательными способностями. обесценение. Все субъекты дали письменное информированное согласие. Протокол эксперимента был одобрен наблюдательным советом по этике в каждом учреждении.
Перед исследованием испытуемые проводили от 1 до 3 ночей привыкания в лаборатория сна.В исследованиях, включая гормональные измерения (133 из 149 исследований), катетер вводили в вену предплечья и брали пробы крови. собираются с 15–30-минутными интервалами в течение 24–25 часов. В течение ночи, катетер был подсоединен к трубке, идущей в соседнюю комнату, чтобы избежать тревожить тему. Получены ночные полиграфические записи сна. Испытуемые оставались лежать в постели в темноте не менее 8 часов. Дневное время дремать не разрешалось.
Профили сна, кортизола и GH были получены у 132, 124 и 114 человек. предметы соответственно.Сопутствующие профили сна, кортизола и GH были получено по 94 предметам.
Запись и анализ сна
Полиграфические записи сна визуально оценивались через 20 или 30 секунд. интервалы в стадиях бодрствования, 1, 2, 3 и 4 и REM с использованием стандартизованных критериев. 38 Начало сна и утреннее пробуждение определялись соответственно, как время появления первого и последнего интервала, оцениваемого как этап 2, 3, 4 или REM.Период сна был определен как интервал, разделяющий сон начало с утреннего пробуждения. Общее время сна рассчитывалось как время сна. период минус общая продолжительность пробуждений. Общая продолжительность каждого стадия выражалась в минутах, а также в процентах от периода сна. Медленноволновый сон определялся как сумма стадий 3 и 4.
Во всех исследованиях уровень кортизола измерялся стандартным радиоиммуноанализом. (РИА).Предел чувствительности составил в среднем 27,6 нмоль / л, а коэффициенты внутри анализа вариации (CV) составляли от 5% до 10%.
В 89 профилях концентрации GH были измерены с помощью RIA с чувствительностью 0,4 мкг / л. 39 Диапазон CV внутри анализа от 5% до 9%. CV между исследованиями в среднем составлял 15%. В 25 исследованиях концентрации GH были измерены хемилюминесцентным методом (8 исследований: Nichols Institute Диагностика, Сан-Хуан-Капистрано, Калифорния; и 17 исследований: диагностический продукт Corporation, Лос-Анджелес, Калифорния) с пределом чувствительности 0.002 до 0,003 мкг / л, CV внутри анализа в диапазоне от 4,8% до 9,9% и межисследовательский CV менее 8%. Исходные, т.е. нелегальные, концентрации GH менее 1 мкг / л по хемилюминесценции соответствуют концентрациям меньше, чем предел чувствительности (0,4 мкг / л) по РИА. Оценка пульсирующего GH секреция выше исходного уровня, полученного из профилей GH, измеренных с помощью хемилюминесценция существенно не отличается от измеренных методом РИА. 40
Анализ индивидуальных профилей кортизола
Циркадные колебания кортизола в плазме были количественно определены с использованием наиболее подходящего кривая на основе расчетов периодограммы. 41 Акрофаза и надир определялись, соответственно, как время появления максимума и минимума наиболее подходящей кривой. Значение акрофазы или надир определялся как уровень, достигаемый кривой наилучшего соответствия в акрофазе. или надир.
Анализ индивидуальных профилей GH
Значительные импульсы секреции GH выше исходного уровня были идентифицированы с использованием компьютеризированного алгоритма. 42 Порог значимости был установлен в 2 раза выше CV внутри анализа. Для каждый значимый импульс, количество GH, секретируемого выше базового уровня, было оценивается математической деконволюцией на основе 1-секционной модели для Клиренс GH и переменный индивидуальный период полураспада. 43 Было определено общее количество GH, секретируемого за данный интервал времени. суммируя количества, выделяемые в каждом из импульсов, происходящих во время этого интервал времени.
Каждый из параметров, используемых для количественной оценки сна, секреции GH и 24-часовой профиль кортизола использовался в 2 анализах. Во-первых, параметр был рассматривается как зависимая переменная в дисперсионном анализе (ANOVA), включая возраст, ИМТ и взаимодействие возраст × ИМТ в качестве независимых переменных. Когда возраст взаимодействия × ИМТ был P > 0,05 на основе сумма квадратов типа III, анализ повторяли только с возрастом и ИМТ. как независимые переменные.Чтобы убедиться, что значимые взаимодействия не отражают воздействие одного предмета, расчеты повторяли после исключение данных из наиболее удаленного объекта для переменных в анализе. Взаимодействие сохранялось в анализе только в том случае, если статистическая значимость не зависел критически ни от одного предмета. Во-вторых, данные были сгруппированы по возрастным группам (16-25 лет — 42 человека; 26-35 лет — 28 человек; в возрасте 36-50 лет: 26 человек; в возрасте 51-60 лет — 23 человека; в возрасте 61-70 лет, 18 предметов; 71-83 года — 12 человек).Для каждого параметра простой линейные регрессии с возрастом в качестве независимой переменной рассчитывались отдельно для субъектов от раннего взросления (в возрасте 16-25 лет) до 43 лет, т. е. средняя точка среднего возраста (в возрасте 36-50 лет), а для субъектов среднего возраста до позднего возраста (в возрасте 44-83 года). Если не указано иное, все значения группы выражаются как среднее (SEM).
В соответствии с предыдущими отчетами, 3 -5 период сна не зависел от возраста.Напротив, всего время сна заметно сокращается с возрастом ( P <0,001), но значительное сокращение общего времени сна произошло только после среднего возраста. С среднего возраста до восьмого десятилетия общее время сна в среднем сокращалось. на 27 минут за декаду (Таблица 1).
Старение по-разному влияло на параметры сна (рис. 1). От раннего взрослого до среднего возраста (возраст от 16-25 до 36-50 лет), процент сна SW снизился с 18.От 9% (1,3%) до 3,4% (1,0%), и это уменьшение глубокого медленного сна было компенсировано увеличением светового медленный сон (т.е. стадии 1 и 2) с 51,2% (1,4%) до 67,3% (1,6%) без значительное изменение времени, проведенного без сна. Изменений в фазе быстрого сна не было. от раннего взрослого до среднего возраста. Увеличение времени бодрствования и уменьшение REM сон стал значительным, начиная с среднего возраста, а стадии 1 и 2 уменьшились с 60,5% (2,5%) для субъектов в возрасте от 51 до 60 лет до 50,6% (4.6%) для субъектов старше 70 лет. Изменения в режиме сна после 50 лет не были значительными.
Влияние ИМТ и взаимодействия возраст × ИМТ было значительным. для стадий 1 и 2 и сна SW, но не для других параметров сна (рисунок 1). У субъектов молодого и среднего возраста (в возрасте 16-43 года), но не у пожилых людей (в возрасте 44-83 года), более высокий ИМТ был связан с более поверхностным, медленным сном (меньше сна, больше стадий 1 и 2 сна).
Средние 24-часовые профили GH от 8 пожилых мужчин и 8 молодых мужчин, которые были сопоставлены для ИМТ иллюстрируют временное совпадение основного импульса GH с ранним сон и заметное снижение уровня гормона роста в пожилом возрасте (рис. 2).
Влияние возраста на секрецию GH в течение 24-часового цикла, время бодрствования, и сон, как показано на рисунке 3. Были очевидны значительные эффекты возраста, не зависящие от ИМТ. С юных лет к среднему возрасту секреция GH снизилась почти на 75%. Дальнейшее меньшее уменьшение произошло в период между средним и поздним взрослым возрастом (Таблица 1).
Значительная отрицательная связь ИМТ с 24-часовой секрецией GH и секреция GH во время бодрствования, но не во время сна, была обнаружена независимо возраста.
Средние суточные профили кортизола у молодых и пожилых мужчин показаны в нижние панели рисунка 2. В обоих группы, уровни кортизола показывают подъем рано утром, уровни снижаются в дневное время и в период ночного покоя. Различия в возрасте в основном проявляются вечером и в начале ночи.
На рисунке 4 показаны изменения. в 24-часовом среднем уровне кортизола, утренней акрофазе и вечернем низком уровне совершеннолетие.Умеренное влияние старения на средний 24-часовой уровень кортизола было незначительным. обнаружен. Старение ассоциировалось с повышением вечернего надира, но утренние максимальные значения оставались стабильными во всех возрастных диапазонах. Увеличивается вечерний уровень кортизола стал очевидным после достижения среднего возраста (таблица 1).
Не было влияния ИМТ или взаимодействия ИМТ × возраст на любой из параметров, характеризующих суточный профиль кортизола.
Связь между сном и гормональными изменениями
Связанное с возрастом снижение секреции гормона роста и быстрого сна происходило аналогичным образом. хронология, при этом большинство декрементов приходится на молодую взрослую жизнь, тогда как возрастное повышение вечернего кортизола не происходило до пятая декада, когда уменьшается фаза быстрого сна и увеличивается количество бодрствования время стало очевидным (таблица 1).Таким образом, мы стремились определить, действительно ли сопутствующее изменение сна и его Взаимодействие с возрастом способствовало гормональным изменениям.
Анализ вариации секреции GH во время сна в зависимости от возраста, SW-сон и их взаимодействие показали, что SW-сон ( P <0,001) и взаимосвязь возраст × SW-сон ( P = 0,008) составляли большую часть дисперсии, и это эффекты возраста сами по себе были незначительными ( P >.50). Похожий результаты были получены при анализе общей 24-часовой секреции GH (возраст, P > 0,50; SW сон, <,001; возраст × SW сна, P = 0,003). В молодом и среднем возрасте у испытуемых, но не у пожилых мужчин, повышенное количество СН-сна было связано с более высоким уровнем секреции GH. На левой панели рисунка 5 сравниваются уровни гормона роста во время сна у субъектов, у которых был большой количества сна SW и у субъектов, соответствующих возрасту и ИМТ, у которых было небольшое количество SW сна.
Разница уровней кортизола в вечернее время была проанализирована в зависимости от возраст, быстрый сон, время бодрствования и их взаимодействие. Вклад всех взаимодействия и время бодрствования не были значительными. Возраст ( P <0,001) и, в меньшей степени, фаза быстрого сна ( P <0,10) были отрицательно связаны с вечерними концентрациями кортизола. Чтобы проиллюстрировать обратную зависимость между количеством быстрого сна и вечера уровни кортизола, правые панели рисунка 5 показан средний надир кортизола у субъектов с большим количеством Быстрый сон и у субъектов, соответствующих возрасту и ИМТ, с небольшим количеством быстрого сна.
Настоящий анализ показывает, что у здоровых мужчин старение влияет на SW-сон и выброс гормона роста с аналогичной хронологией, характеризующейся значительным снижением от раннего взрослого до среднего возраста. Напротив, влияние возраста на быстрый сон, фрагментация сна, и функция HPA становится очевидной позже в жизни. Анализ также предполагает, что возрастные изменения в соматотропная и кортикотропная оси могут частично отражать снижение сна качественный.
Человеческий сон находится под двойным контролем циркадной ритмичности и гомеостатический процесс, связывающий глубину сна с продолжительностью предшествующего бодрствование. 44 Этот гомеостатический процесс включает предполагаемый нервный фактор сна, который увеличивается во время бодрствования и экспоненциально затухает во сне. Медленный сон в первую очередь контролируется гомеостатическим процессом. Циркадная ритмичность — это колебание с периодом около 24 часов. с помощью кардиостимулятора, расположенного в супрахиазматическом ядре гипоталамуса.Циркадный ритмичность играет важную роль в выборе времени сна, консолидации сна, и распределение быстрого сна. 45 Настоящее данные показывают, что изменение гомеостаза сна и бодрствования является ранним биологическим маркер старения у взрослых мужчин. Напротив, компоненты сна, находящиеся ниже контроль циркадного кардиостимулятора относительно хорошо сохранился до позднего возраста.
Хронология старения секреции GH в значительной степени следует модели похож на сон SW.Таким образом, у мужчин возникает так называемая «соматопауза». в раннем зрелом возрасте, в возрасте от 25 до 35 лет, возрастной диапазон, соответствующий к продолжительности жизни человека до развития современной цивилизации и практически завершится к концу четвертого десятилетия. Наши анализы также указывают на то, что снижение количества сна, вызванного SW, независимо от возраста, является частично отвечает за снижение секреции GH в среднем и позднем возрасте. Что это коррелятивное свидетельство отражает общий механизм, лежащий в основе СН-сна. поддерживается генерация и выпуск GH, а не косвенная связь 2 исследованиями, которые показали, что фармакологическое улучшение сна приводит к увеличению выпуска GH. 46 , 47 Также подтверждается причинно-следственная связь между ухудшением качества сна и снижение секреции ночного гормона роста — это исследования у пациентов с апноэ во сне, показывающие заметное увеличение выброса GH после лечения с положительным давлением в дыхательных путях. 48 , 49 Обратное взаимодействие между сном и GH, то есть пагубное влияние пониженной соматотропной функции на сон, также возможно, поскольку исследования как в нормальных, так и в патологических состояниях показали, что фактор высвобождения GH и GH влияют на качество сна. 12 , 50 В настоящем исследовании пациентов, не страдающих ожирением, мужчин, обнаружение отрицательного влияния ИМТ на секрецию гормона роста во время бодрствования и количество сна SW согласуется с гипотезой о том, что ингибирование ось GH может отрицательно влиять на регуляцию сна.
Хотя клинические последствия пониженного сна во время сна до сих пор неясны, относительный дефицит гормона роста у пожилых людей связан с повышенным содержанием жира тканевое и абдоминальное ожирение, снижение мышечной массы и силы, а также работоспособность. 51 -53 В настоящее время многочисленные испытания изучают клиническую полезность и безопасность. заместительной терапии рекомбинантным GH, другими гормонами оси GH, и синтетические стимуляторы секреции GH у пожилых людей без патологического дефицита GH. Хотя преимущества таких вмешательств все еще не доказаны, настоящие результаты предлагают, чтобы они были нацелены на более молодой возрастной диапазон, чем предполагается в настоящее время, т. е. люди в раннем среднем возрасте, а не лица старше 65 лет, когда периферические ткани постоянно подвергались воздействию очень низких уровней гормона роста. минимум 2 десятилетия.Кроме того, поскольку фармакологическое усиление Было показано, что сон SW у молодых людей приводит к одновременному и пропорциональному увеличение выпуска GH 46 , 47 и текущие исследования в нашей лаборатории показывают, что аналогичные эффекты могут быть получены у пожилых людей препараты, которые надежно стимулируют SW-сон, могут представлять новый класс секретарей GH.
Настоящие данные демонстрируют, что количество быстрого сна сокращается. примерно на 50% в пожилом возрасте по сравнению с молодостью.Однако уменьшенные суммы фазы быстрого сна и значительной фрагментации сна не происходит до возраст 50 лет. Влияние старения на уровень кортизола следует той же хронологии. Старение было связано с повышением уровня кортизола в вечернее время, что отражает нарушение способности достигать вечернего покоя после утренней стимуляции. Исследования как на животных, так и на людях показали, что вредные эффекты гиперактивности HPA более выражены во время впадины ритм, чем во время пика. 25 , 54 Таким образом, небольшое повышение уровня кортизола вечером может способствовать развитию центральных и периферических нарушений, связанных с избытком глюкокортикоидов, такие как дефицит памяти и инсулинорезистентность, 24 , 25 и еще больше способствуют фрагментации сна. Действительно, повышенный уровень кортизола может способствовать пробуждению. 55 , 56
Повышенный уровень кортизола вечером в пожилом возрасте, вероятно, отражает нарушение отрицательной обратной связи контроля оси HPA при старении.Наши анализы предполагают что существует связь между этим изменением функции HPA и уменьшением количество быстрого сна, не зависящее от возраста. Данные в целом поддерживают концепция о том, что снижение качества сна способствует аллостатической нагрузке, т.е. износ в результате чрезмерной активности систем, чувствительных к стрессу. 57
Настоящее исследование посвящено влиянию старения на отношения между сном и соматотопической и кортикотропной осями у мужчин, потому что преобладающая секреция GH происходит во время сна у мужчин, но не у женщин 11 и поскольку есть данные, позволяющие предположить, что заметное уменьшение сна в раннем взрослом возрасте происходит у мужчин, но не у женщин. 8 -11 Ли выводы, аналогичные тем, которые получены для мужчин, и для женщин, потребуют отдельная оценка как половых различий в качестве сна, так и 24-часового профили секреции гормона роста и кортизола у молодых и пожилые люди. 11 , 12,22
В заключение, у здоровых мужчин отчетливые изменения качества сна которые характеризуют переходы от раннего зрелого возраста к среднему возрасту, на с одной стороны, и от среднего возраста до старости, с другой стороны, все они связаны со специфическими изменениями в гормональной системе, которые важны для метаболизма регулирование.Стратегии предотвращения или ограничения снижения качества сна у Следовательно, средний и поздний период жизни может представлять собой косвенную форму гормонального терапия с возможными благотворными последствиями для здоровья.
1.Принц ПН. Нарушения сна и сна у пожилых людей. J Clin Neurophysiol. 1995; 12: 139-146.Google Scholar2, Файнберг И., Кореско Р.Л., Хеллер Н. Модели сна на ЭЭГ как функция нормального и патологического старения в организме человека. человек. Psychiatry Res. 1967; 5: 107-144.Google Scholar3. Бенка Р.М., Обермейер WH, Thisted RA, Gillin JC. Сон и психические расстройства. Arch Gen Psychiatry. , 1992; 49: 651-668.Google Scholar4.Bliwise DL. Нормальное старение. В: Kryger MH, Roth T, Dement WC, eds. Принципы и Практика медицины сна . Филадельфия, Пенсильвания: У. Б. Сондерс; 1994: 26-39.
5.Файнберг И. Функциональные последствия изменений физиологии сна с возрастом. В: Терри Р.Д., Гершон С., ред. Нейробиология старения . Нью-Йорк, Нью-Йорк: Raven Press; 1976: 23-41.
6. Элерс К.Л., Купфер Д.И. Влияние возраста на параметры дельта- и быстрого сна. Электроэнцефалогер Клин Нейрофизиол. 1989; 72: 118-125.Google Scholar7.Landolt HP, Dijk DJ, Achermann P, Borbely AA. Влияние возраста на ЭЭГ сна: медленноволновая активность и частота веретена у мужчин молодого и среднего возраста. Brain Res. 1996; 738: 205-212.Google Scholar8.Webb WB. Сон у пожилых людей: структура сна мужчин от 50 до 60 лет и женщины. J Gerontol. 1982; 37: 581-586.Google Scholar9.Dijk DJ, Beersma DG, Bloem GM. Половые различия в ЭЭГ сна молодых людей: визуальная оценка и спектральный анализ. Сон. 1989; 12: 500-507.Google Scholar 10. Муртазаев М.С., Кемп Б., Звиндерман А.Х., Кампхуйзен Х.А. Возраст и пол влияют на различные характеристики медленных волн в ЭЭГ сна. Сон. 1995; 18: 557-564.Google Scholar11.Ehlers CL, Kupfer DJ. Медленноволновой сон: по-разному ли стареют молодые взрослые мужчины и женщины? J Sleep Res. 1997; 6: 211-215.Google Scholar 12. Ван Каутер Э, Плат Л., Копински Г. Взаимосвязь между сном и соматотропной осью. Сон. 1998; 21: 553-566.Google Scholar13.Ho KY, Evans WS, Blizzard RM. и другие. Влияние пола и возраста на 24-часовой профиль секреции гормона роста у человека: важность эндогенных концентраций эстрадиола. J Clin Endocrinol Metab. 1987; 64: 51-58.Google Scholar14. Van Coevorden A, Mockel J, Laurent E. и другие. Нейроэндокринные ритмы и сон у пожилых мужчин. Am J Physiol. , 1991; 260: E651-E661.Google Scholar, 15.Kern W., Dodt C, Born J, Fehm HL. Изменения секреции кортизола и гормона роста во время ночного сна в процессе старения. J Gerontol A Biol Sci Med Sci. 1996; 51A: M3-M9.Google Scholar 16. Weitzman ED, Zimmerman JC, Cheisler CA, Ronda JM. У нормального человека секреция кортизола подавляется во время сна. J Clin Endocrinol Metab. 1983; 56: 352-358. Google Scholar, 17, Spath-Schwalbe E, Uthgenannt D, Voget G, Kern W., Born J, Fehm HL.Адренокортикотропин и кортизол, индуцированные кортикотропин-рилизинг-гормоном секреция зависит от сна и бодрствования. J Clin Endocrinol Metab. , 1993; 77: 1170-1173.Google Scholar. 18.Spath-Schwalbe E, Uthgenannt D, Korting N, Fehm HL, Born J. Сон и бодрствование влияют на реакцию гипофизарно-надпочечников. ось к аргинину вазопрессину у людей. Нейроэндокринология. , 1994; 60: 544-548.Google Scholar, 19, Gronfier C, Luthringer R, Follenius M. и другие. Временные отношения между пульсирующей секрецией кортизола и электроэнцефалографическими данными. активность во сне у человека. Электроэнцефалогер Клин Нейрофизиол. 1997; 103: 405-408. Google Scholar20. Бервольф С., Струве К., Маршалл Л., Фем Х.Л. Медленный сон вызывает угнетение гипофизарно-надпочечниковой секреции в люди. J Нейроэндокринол. 1997; 9: 479-484.Google Scholar 21. Лепроулт Р., Копински Г., Бакстон О., Ван Каутер Э. Потеря сна приводит к повышению уровня кортизола на следующий вечер. Сон. 1997; 20: 865-870.Google Scholar 22. Ван Каутер Э, Лепроулт Р., Купфер Д. Влияние пола и возраста на уровни и циркадную ритмичность плазменный кортизол. J Clin Endocrinol Metab. 1996; 81: 2468-2473. Google Scholar23, Симан Т.Э., Роббинс Р.Дж. Старение и ответ гипоталамо-гипофизарно-надпочечников на вызов у людей. Endocr Rev. 1994; 15: 233-260.Google Scholar24. McEwen BS, Sapolsky RM. Стресс и когнитивные функции. Curr Opin Neurobiol. 1995; 5: 205-216.Google Scholar25, Даллман М.Ф., Страк А.Л., Акана С.Ф. и другие. Пир и голод: решающая роль глюкокортикоидов с инсулином в суточный поток энергии. Фронт нейроэндокринол. 1993; 14: 303-347. Google Scholar, 26. Линковски П., Мендлевич Дж., Леклерк Р. и другие. 24-часовой профиль адренокортикотропина и кортизола при большом депрессивном состоянии. болезнь. J Clin Endocrinol Metab. 1985; 61: 429-438.Google Scholar 27. Копински Г., Ван Ондерберген А., Л’Эрмит-Балерио М. и другие. Эффекты бензодиазепинтриазолама короткого действия, принимаемого перед сном, на циркадный ритм и гормональный фон, связанный со сном, у нормальных мужчин. Сон. 1990; 13: 232-244.Google Scholar 28.Ван Каутер Э., Блэкман Дж. Д., Роланд Д., Спайр Дж. П., Рефетов С., Полонский К. С.. Модуляция регуляции глюкозы и секреции инсулина циркадными ритмами. ритмичность и сон. J Clin Invest. 1991; 88: 934-942.Google Scholar 29. Фрэнк С., Роланд, округ Колумбия, Стурис Дж. и другие. Влияние старения на регуляцию глюкозы во время бодрствования и сна. Am J Physiol. , 1995; 269: E1006-E1016.Google Scholar 30, Бистон П., Ван Каутер Э., Офек Г., Линковски П., Полонский К.С., Дегауте Дж. Суточные колебания сердечно-сосудистой функции и регуляции глюкозы у нормотензивных людей. Гипертония. 1996; 28: 863-871.Google Scholar 31. Копински Г., Лепроулт Р., Ван Ондерберген А. и другие. Длительное пероральное лечение МК-677, нового средства, увеличивающего секрецию гормона роста, улучшает качество сна у человека. Нейроэндокринология. 1997; 66: 278-286.Google Scholar 32. Линковски П., Шпигель К., Керкхофс М. и другие. Генетические факторы и влияние окружающей среды на секрецию пролактина во время бодрствования и во сне. Am J Physiol. 1998; 274: E909-E919. Google Scholar 33.Джарретт DJ, Greenhouse JB.Циркадный ритм секреции кортизола в амбулаторных условиях не нарушается. с большим депрессивным расстройством. В: Программа Первого собрания Общества исследований в области биологических Ритмы; 11-14 мая 1988 г .; Чарльстон, Южная Каролина. Abstract 97.
34. Джарретт ДБ, Поллок Б., Мевальд Дж. М., Купфер Д. Д.. Острые эффекты кломипрамина внутривенно на гормон сна секреция у амбулаторных пациентов с депрессией и здоровых контрольных субъектов. Biol Psychiatry. 1991; 29: 3-14.Google Scholar 35.Rubin RT, Poland RE, Lesser IM, Winston RA, Blodgett AL.Нейроэндокринные аспекты первичной эндогенной депрессии, I: кортизол секреторная динамика у пациентов и контрольной группы. Arch Gen Psychiatry. 1987; 44: 328-336. Google Scholar, 36. Рубин Р.Т., Польша RE, Lesser IM. Нейроэндокринные аспекты первичной эндогенной депрессии, X: рост сыворотки гормональные меры у пациентов и подобранных контрольных субъектов. Biol Psychiatry. 1990; 27: 1065-1082.Google Scholar 37.Вгонцас А.Н., Папниколау Д.А., Бикслер Э.О. и другие. Апноэ во сне, дневная сонливость и утомляемость: отношение к висцеральным ожирение, инсулинорезистентность и гиперцитокинемия. J Clin Endocrinol Metab. 2000; 85: 1151-1158. Google Scholar 38.Rechtschaffen A, Kales A. Руководство по стандартизированной терминологии, методикам и система оценки стадий сна людей . Лос-Анджелес, Калифорния: Информационная служба мозга UCLA / Исследования мозга Институт; 1968.
39. Вирасоро Э., Копински Г., Бруно О.Д., Леклерк Р. Радиоиммуноанализ гормона роста человека с использованием разделения древесный уголь и декстран. процедура. Clin Chim Acta. 1971; 31: 294-297.Google Scholar40.L’Hermite-Balériaux M, Copinschi G, Van Cauter E. Анализы гормона роста: сравнение ранних и последних поколений. Clin Chem. 1996; 42: 1789-1795.Google Scholar 41. Ван Каутер Э. Метод характеристики 24-часовой временной изменчивости компонентов крови. Am J Physiol. 1979; 237: E255-E264.Google Scholar42.Van Cauter E. Оценка ложноположительных и ложноотрицательных ошибок при анализе гормональная пульсация. Am J Physiol. 1988; 254: E786-E794.Google Scholar43.Van Cauter E, Kerkhofs M, Caufriez A, Van Onderbergen A, Thorner MO, Copinschi G. Количественная оценка секреции GH у нормального человека: воспроизводимость и отношение ко сну и времени суток. J Clin Endocrinol Metab. , 1992; 74: 1441-1450. Google Scholar, 44, Борбели А.А. Процессы, лежащие в основе регуляции сна. Horm Res. 1998; 49: 114-117.Google Scholar 45.Dijk DJ, Cheisler CA. Вклад циркадного кардиостимулятора и гомеостата сна в склонность ко сну, структура сна, электроэнцефалографические медленные волны и активность веретена сна у человека. J Neurosci. 1995; 15: 3526-3538.Google Scholar 46. Ван Каутер Э, Плат Л., Шарф М. и другие. Одновременная стимуляция медленного сна и секреции гормона роста γ-гидроксибутиратом у нормальных молодых мужчин. J Clin Invest. 1997; 100: 745-753. Google Scholar, 47. Gronfier C, Luthringer R, Follenius M. и другие. Количественная оценка взаимосвязи между гормоном роста секреция и электроэнцефалографическая активность дельта-волн во время нормального сна и после обогащения дельта-волнами. Сон. 1996; 19: 817-824.Google Scholar 48.Saini J, Krieger J, Brandenberger G, Wittersheim G, Simon C, Follenius M. Непрерывное лечение положительным давлением в дыхательных путях: влияние на гормон роста, профили инсулина и глюкозы у пациентов с обструктивным апноэ во сне. Horm Metab Res. 1993; 25: 375-381.Google Scholar, 49. Купер Б.Г., Уайт Дж. Э., Эшворт Л.А., Альберти К.Г., Гибсон Г.Дж.. Гормональные и метаболические профили у лиц с нарушением сна синдром апноэ и эффекты постоянного положительного давления в дыхательных путях через нос (CPAP) лечение. Сон. 1995; 18: 172-179. Google Scholar 50. Aström C. Взаимодействие между сном и гормоном роста, оцененное с помощью ручной полисомнографии. и автоматический спектральный анализ мощности. Acta Neurol Scand. 1995; 92: 281-296. Google Scholar, 51. Cuneo RC, Salomon F, McGauley GA, Sonksen PH. Синдром дефицита гормона роста у взрослых. Clin Endocrinol. 1992; 37: 387-397.Google Scholar52.Corpas E, Harman SM, Blackman MR. Гормон роста человека и старение человека. Endocr Rev. 1993; 14: 20-39. Google Scholar. 53. Розен Т., Ханссон Т., Гранхед Х., Сцук Дж., Бенгтссон Б.А. Снижение содержания минералов в костях у взрослых пациентов с гормоном роста дефицит. Acta Endocrinol. 1993; 129: 201-206.Google Scholar 54. Plat L, Féry F, L’Hermite-Balériaux M, Mockel J, Van Cauter E. Метаболические эффекты кратковременных физиологических подъемов плазмы кортизол более выражен вечером, чем утром. J Clin Endocrinol Metab. 1999; 84: 3082-3092.Google Scholar 55.Holsboer F, von Bardelein U, Steiger A. Влияние внутривенного введения кортикотропин-рилизинг-гормона на связанные со сном выброс гормона роста и ЭЭГ сна у человека. Нейроэндокринология. 1988; 48: 32-38. Google Scholar 56 Борн Дж., Шпет-Швальбе Э., Швакенхофер Х.