Что это такое — модель скелета человека?
Скелет — совокупность костей, образующих твёрдый остов (твёрдый скелет), составляющий 1/5—1/7 массы тела человека. Функции скелета: опорно-защитная, двигательная, депо минеральных солей. Анатомические образования к костям прикрепляет мягкий скелет, к которому относятся сухожилия, связки, фасции. Скелет ограничивает полости с расположенными в них внутренними органами: грудную, брюшную, малого таза, черепа, — защищая от внешних воздействий внутренние органы и являясь их опорой. Он состоит более чем из 200 костей. В это число входит 33—34 пары (66—68 костей), остальные кости непарные. Отделы скелета: скелет головы — череп; скелет туловища — позвоночный столб и грудная клетка; скелет верхних и нижних конечностей состоит из скелета поясов конечностей — плечевого и тазового и скелета свободных конечностей. Череп, позвоночный столб, грудную клетку относят к осевому скелету (скелету туловища).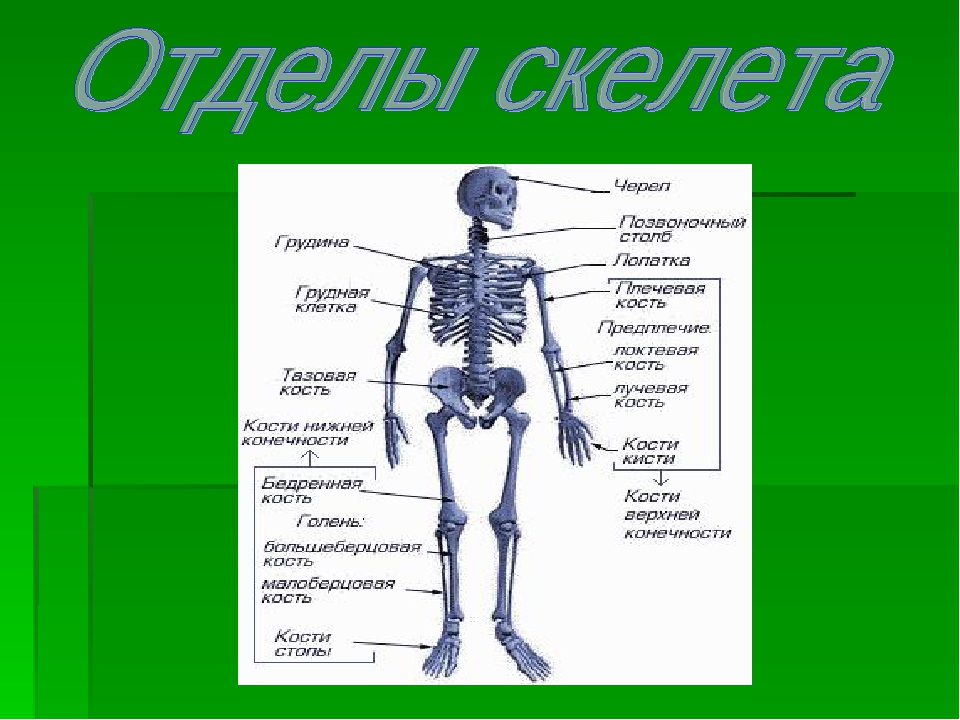
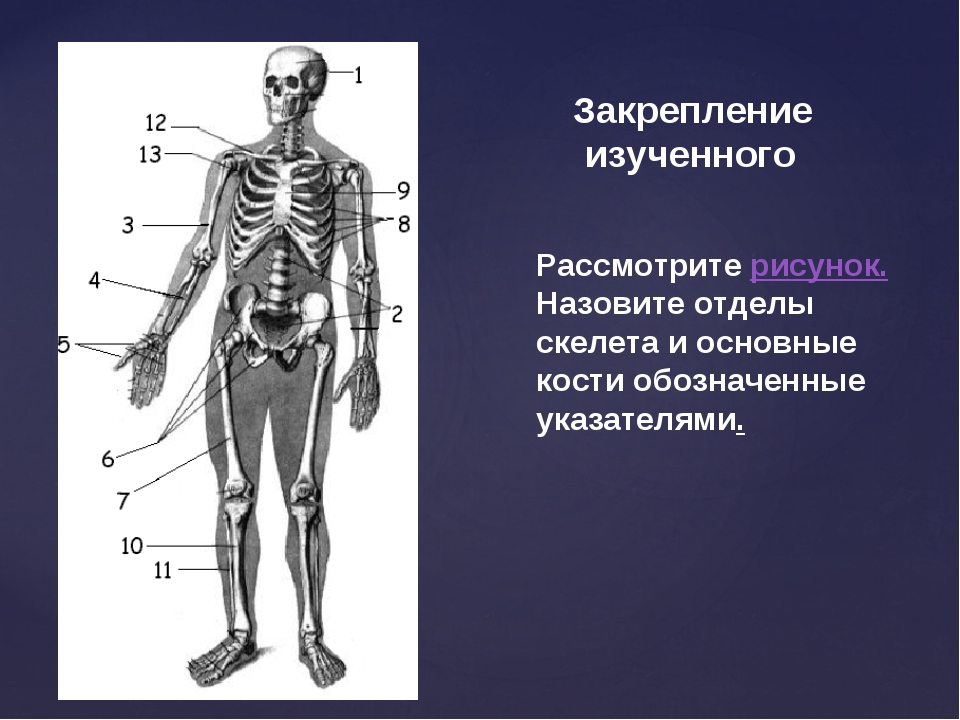
Скелет человека.
I — череп; 2 — позвоночный столб; 3 — ключица; 4 —ребро; 5 — грудина; 6 — плечевая кость; 7 — лучевая кость;8 — локтевая кость; 9 — кости запястья; 10 — кости пясти;
II — фаланги пальцев кисти;12 — седалищная кость; 13 —кости плюсны; 14 — кости предплюсны; 15 — большеберцовая кость; 16 — малоберцовая кость; 17 — надколенник; 18 — бедренная кость;19 — лобковая кость; 20 —подвздошная кость.
Соединения костей
Соединения скрепляют кости скелета в целое и сочетают прочность с упругостью и подвижностью. Различают 3 вида соединений: непрерывные, прерывные (суставы) и симфизы (полусуставы).
Непрерывные соединения, синартрозы, прочны, упруги, их подвижность ограничена. В зависимости от вида ткани они делятся на фиброзные, хрящевые, костные.
Фиброзные соединения — это соединение костей плотной, волокнистой соединительной тканью. Различают три вида непрерывных соединений: синдесмозы, швы, вколачивание.
Виды соединений костей (схема). А — сустав; Б — фиброзное соединение; В — хрящевое соединение; Г — полусустав; 1 — кость; 2 —соединительная ткань; 3 — хрящ; 4 — полость сустава; 5 — капсула сустава; 6 — связка, укрепляющая сустав; 7 — синовиальная оболочка; 8 — хрящевой диск; 9 — щель в хрящевом диске.
Синхондрозы — соединения костей с помощью хрящевой ткани. Упругость хрящевой прослойки и, следовательно, амортизационные свойства зависят от толщины хряща. Постоянные синхондрозы существуют в течение всей жизни (межпозвоночные диски). Непостоянные синхондрозы сохраняются до определённого возраста, а затем замещаются костным сращением — синостозом (например, соединение рукоятки грудины с её телом).
Постоянные синхондрозы существуют в течение всей жизни (межпозвоночные диски). Непостоянные синхондрозы сохраняются до определённого возраста, а затем замещаются костным сращением — синостозом (например, соединение рукоятки грудины с её телом).
Симфиз — промежуточное между синартрозом и суставом соединение. Это фиброзная ткань или волокнистый хрящ с узкой щелевидной полостью в толще: симфизы между рукояткой и телом грудины, межпозвоночный (между крестцом и копчиком), лобковый. Такое соединение не имеет суставной капсулы, синовиальной оболочки, но может быть укреплено связками.
Строение суставов
Прерывные соединения костей (синовиальные соединения или суставы) — подвижные соединения, отличающиеся большим разнообразием движений. Воспаление сустава — артрит. Основной аппарат сустава — суставные поверхности, покрытые хрящом, суставная капсула и суставная полость, содержащая синовиальную жидкость. Вспомогательный аппарат сустава — хрящевые диски, мениски, суставные губы. Суставные поверхности у большинства суставов соответствуют друг другу (конгруэнтны), иногда не соответствуют (не конгруэнтны). Суставной хрящ обычно гиалиновый, реже волокнистый (в височно-нижнечелюстном суставе). Хрящ не имеет сосудов, гладкий, амортизирует толчки, уменьшает трение при движениях суставных поверхностей. Чем больше нагрузка на сустав, тем хрящ толще. Суставная капсула прикрепляется к сочленяющимся костям чаще вблизи краёв суставных поверхностей, образуя герметичную суставную полость. Капсула имеет 2 слоя: наружный (фиброзная мембрана) и внутренний (синовиальная мембрана).
Суставные поверхности у большинства суставов соответствуют друг другу (конгруэнтны), иногда не соответствуют (не конгруэнтны). Суставной хрящ обычно гиалиновый, реже волокнистый (в височно-нижнечелюстном суставе). Хрящ не имеет сосудов, гладкий, амортизирует толчки, уменьшает трение при движениях суставных поверхностей. Чем больше нагрузка на сустав, тем хрящ толще. Суставная капсула прикрепляется к сочленяющимся костям чаще вблизи краёв суставных поверхностей, образуя герметичную суставную полость. Капсула имеет 2 слоя: наружный (фиброзная мембрана) и внутренний (синовиальная мембрана).  Суставная полость — щелевидное пространство между суставными поверхностями, содержит немного синовиальной жидкости.
Суставная полость — щелевидное пространство между суставными поверхностями, содержит немного синовиальной жидкости.Суставные диски и мениски — хрящевые пластинки различной формы, которые располагаются между суставными поверхностями в тех случаях, когда они не соответствуют друг другу (в грудино-ключичном, коленном и других суставах). Эти образования смещаются при движениях в суставе, исправляют несоответствие суставных поверхностей, амортизируют сотрясения и толчки. Суставная хрящевая губа прирастает по краям суставной впадины, дополняя и углубляя её в плечевом, тазобедренном суставе.
Строение сустава (схема). 1 — суставные хрящи; 2 —фиброзная мембрана суставной капсулы; 3 — синовиальная мембрана суставной капсулы;4 — суставная полость; 5 — надкостница; 6 — кость.
Классификация, биомеханика суставов
Суставы скелета человека различаются по форме и количеству сочленяющихся суставных поверхностей. Простой сустав образован двумя суставными поверхностями, сложный сустав — тремя и более суставными поверхностями. Комплексный сустав имеет в полости диск или мениски, которые делят полость сустава на два этажа.
Простой сустав образован двумя суставными поверхностями, сложный сустав — тремя и более суставными поверхностями. Комплексный сустав имеет в полости диск или мениски, которые делят полость сустава на два этажа.
По геометрической форме суставных поверхностей различают суставы цилиндрические, шаровидные, эллипсовидные. Остальные формы суставов возникли как видоизменение основных форм. Например, разновидность цилиндрического сустава — блоковидный сустав (разновидность — винтообразный сустав), шаровидного — чашеобразный и плоский сустав.
Число осей, вокруг которых происходят движения, определяется формой сустава. Одноосные суставы — цилиндрические, блоковидные, винтообразные. Двухосные — эллипсовидные, седловидные, мыщелковые. Трёхосные (они же многоосные за счёт множества продольных осей) — шаровидные, чашеобразные, плоские.
Движения в суставах в зависимости от формы сочленяющихся поверхностей могут совершаться вокруг фронтальной, сагиттальной и продольной осей. Вокруг фронтальной оси совершается сгибание и разгибание, вокруг сагиттальной — отведение и приведение по отношению к срединной плоскости. Вокруг продольной оси осуществляется вращение. При круговом движении, циркумдукции, последовательно используются все оси.
Формы суставов. 1 — блоковидный; 2 — эллипсовидный; 3 — седловидный; 4 — шаровидный.
Объём (амплитуда, размах) движений в суставах зависит от разности угловых величин сочленяющихся поверхностей: чем больше разность, тем больше размах движений. На амплитуду движений в суставе влияют также количество и расположение связок и состояние мышц, окружающих сустав.
Кости (Остеология — Osteologia)
АнатомияКости туловища Кости головы Кости верхней конечности Кости нижней конечности Развитие и возрастные особенности- Остеология, osteologia
- — учение о костях.
 Данный раздел изучает скелет в целом, отдельные кости, костную ткань.
Данный раздел изучает скелет в целом, отдельные кости, костную ткань.
 Данный раздел изучает скелет в целом, отдельные кости, костную ткань.
Данный раздел изучает скелет в целом, отдельные кости, костную ткань.Кости, ossa, являются твёрдой опорой мягких тканей тела и образуют рычаги, перемещающийся силой сокращения мышц.
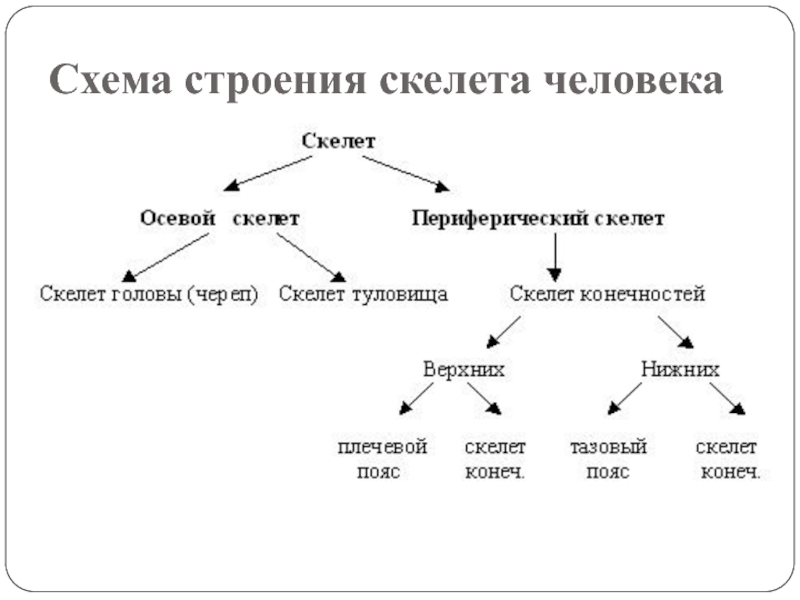
В теле кости образуют систему скелета, systema skeletale, (рис. 1, 2), которая включает осевой скелет, skeleton axiale, и добавочный скелет, skeleton appendiculare,. К осевому скелету относятся череп, cranium,, позвоночный столб, columna vertebralis, и кости грудной клетки, ossa thoracis,. Добавочный скелет объединяет кости верхней конечности, ossa membri superioris, и кости нижней конечности, ossa membri inferioris.рис. 1. Система скелета, systema skeletale; вид спереди.
Система скелета Строение кости Классификация костей по форме и строению Костный мозг Кости осевого скелета Кости добавочного скелета Система скелета включает более 200 костей, из них 85 парных.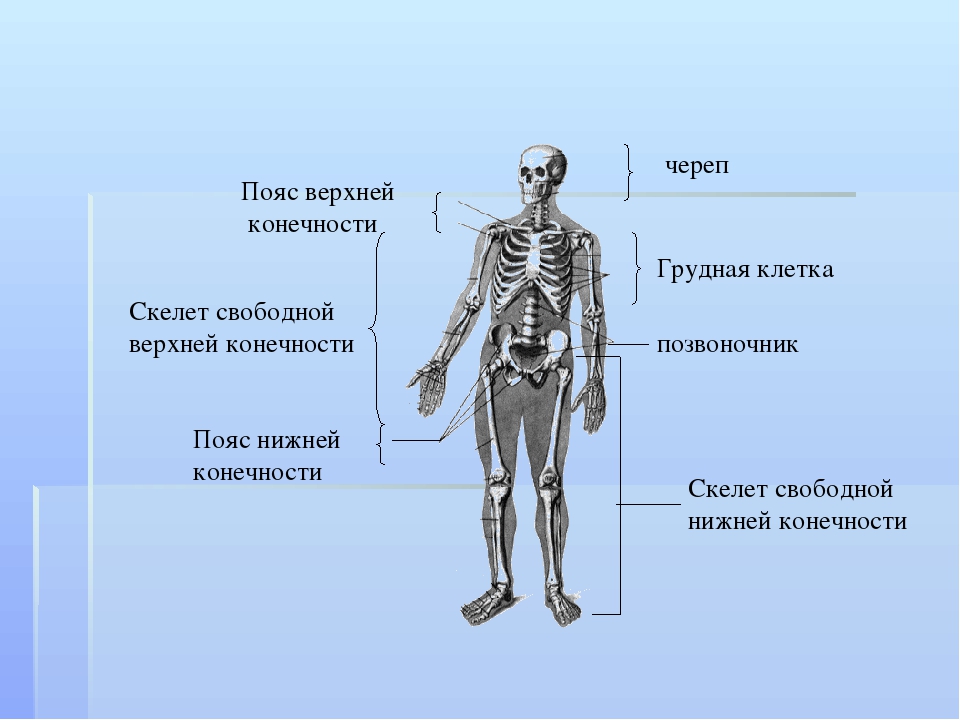
Каждая кость – это орган, построенный из различных видов соединительной ткани, содержащий костный мозг, снабженный сосудами и нервами.рис. 2 Система скелета, systema skeletale; вид сзади.
В системе скелета выделяют костную часть, pars ossea, и хрящевую часть, pars cartilaginosa. Основной в системе скелета является костная часть. В хрящевую часть системы скелета входят суставные хрящи, cartilagines articulares, эпифизарные хрящи, cartilagines epiphysiales, и реберные хрящи, cartilagines costales.
Снаружи кость покрывает тонкая соединительнотканная оболочка – надкостница, periosteum, (рис. 3), в которой различают волокнистый и остеогенный слои. Поверхностно расположенный волокнистый слой соединяется с костью проникающими в нее волокнами (прободающие волокна), содержит кровеносные и лимфатические сосуды, нервы. Отсюда сосуды и нервы проходят в кость через питательные отверстия,  Внутренний, остеогенный слой (рис. 4), включает образующие клетки (остеобласты), участвующие в процессах развития и перестройки костной ткани, в том числе после травм и переломов. На границе с суставным хрящом, покрывающим концы кости, надкостница переходит в надхрящницу, perichondrium. В результате кость оказывается окутанной непрерывной соединительнотканной оболочкой. Эта оболочка покрывает поверхность кости и все находящиеся на ней образования: отростки, processus; ости, spinae; гребни, cristae; бугры, tuberi; бугорки, tuberculi; шероховатые линии, liniae asperae; углубления, foveae; ямки, fossae, и т.д.
Внутренний, остеогенный слой (рис. 4), включает образующие клетки (остеобласты), участвующие в процессах развития и перестройки костной ткани, в том числе после травм и переломов. На границе с суставным хрящом, покрывающим концы кости, надкостница переходит в надхрящницу, perichondrium. В результате кость оказывается окутанной непрерывной соединительнотканной оболочкой. Эта оболочка покрывает поверхность кости и все находящиеся на ней образования: отростки, processus; ости, spinae; гребни, cristae; бугры, tuberi; бугорки, tuberculi; шероховатые линии, liniae asperae; углубления, foveae; ямки, fossae, и т.д.
Более тонкая оболочка – эндост, endosteum, – выстилает кость изнутри.
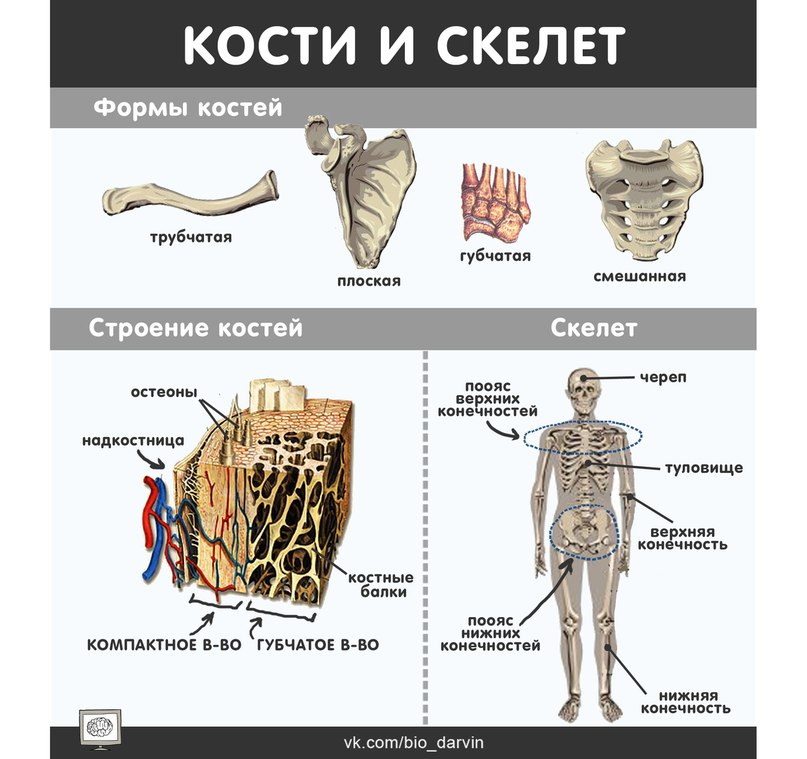
По форме различают кости длинные, ossa longi, короткие, ossa brevia, и плоские, ossa plana, (рис.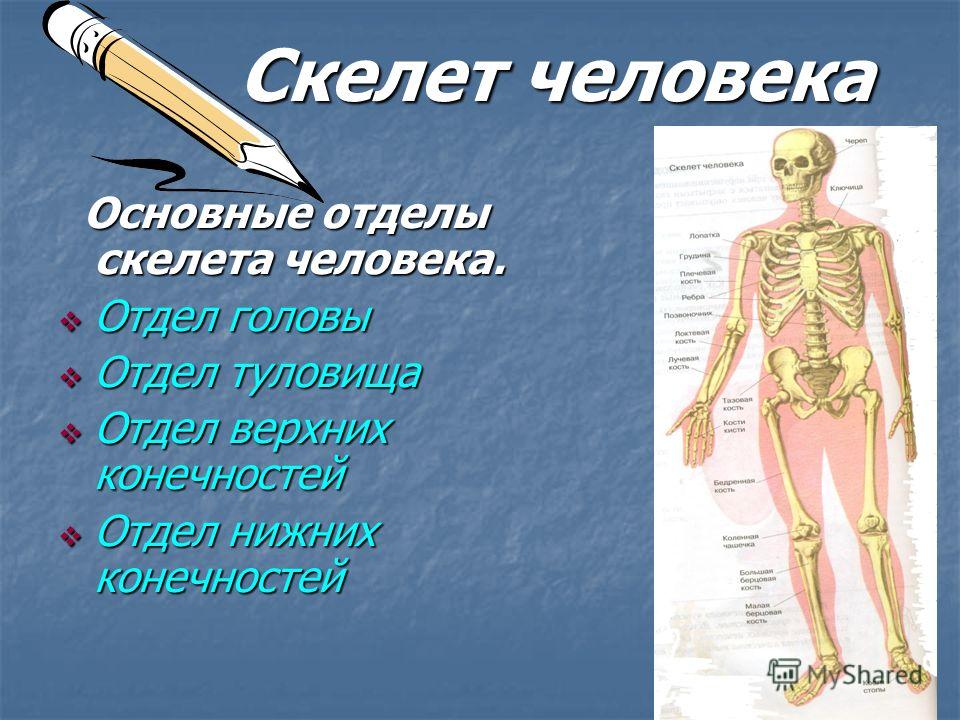 5). Некоторые кости внутри имеют наполненные воздухом полости; такие кости называются воздухоносными, ossa pneumatica. Кроме того, некоторые кости относят к ненормальным (смешанным) костям, ossa irregularea, которые состоят из мастей, имеющих разные форму и строение.
5). Некоторые кости внутри имеют наполненные воздухом полости; такие кости называются воздухоносными, ossa pneumatica. Кроме того, некоторые кости относят к ненормальным (смешанным) костям, ossa irregularea, которые состоят из мастей, имеющих разные форму и строение.
В длинных костях (плечевой, ключице, пястных, фалангах и др.) различают среднюю часть – диафиз, diaphysis, и два концевых отдела – эпифизы, epiphyses. Эпифиз, расположенный ближе к осевому скелету, называют проксимальным, epiphysis proximalis, а эпифиз той же кости, занимающий более отдаленное от осевого скелета положение, – дистальным, epiphysis distalis. Участки длинных костей, находящиеся на границе диафиза и эпифизов, носят название метафизов (metaphyses) (рис. 4). рис. 4. Большеберцовая кость, tibia, правая; вид спереди. (Надкостница удалена. Вскрыта часть костномозговой полости. Эпифизы не сращены с диафизом.) Сама граница заметна только в костях у детей и подростков, пока между диафизом и эпифизами сохраняется прослойка хряща – эпифизарный хрящ, cartilago epiphysialis. За счет этого хряща кость интенсивно растет в длину. Затем эпифизарный хрящ замещается костной тканью, формирующей эпифизарную линию, linea epiphysialis, которая с возрастом становится почти неразличимой.
За счет этого хряща кость интенсивно растет в длину. Затем эпифизарный хрящ замещается костной тканью, формирующей эпифизарную линию, linea epiphysialis, которая с возрастом становится почти неразличимой.
На распиле длинной кости (рис. 6) можно различить компактное вещество, substantia compacta, формирующее наружные слои кости, и губчатое (трабекулярное) вещество, substantia spongiosa, расположенное кнутри от компактного вещества. преимущественно в эпифизах и метафизах. В диафизе длинных костей компактное вещество окружает костномозговую полость, cavitas medullaris, имеющую форму трубки.
рис. 5. Кости, ossa, разные по форме. 1а — локтевая кость, ulna): 1б — таранная кость, talus): 2б — кости запястья, ossa carpi; 2в — позвонок, vertebra; 3 — плоская кость, os planun, (лопатка, scapila)).В коротких костях на распиле у поверхности определяется тонкий слой компактного вещества, окружающий перекладины губчатого вещества, которое формирует большую часть кости. Трабекулы губчатого вещества образуют сложную ячеистую сеть. Они располагаются в каждой кости в строгом соответствии с ее функциональными нагрузками.
Трабекулы губчатого вещества образуют сложную ячеистую сеть. Они располагаются в каждой кости в строгом соответствии с ее функциональными нагрузками.
В плоских костях, напротив, губчатое вещество обычно формирует тонкий слой кости и с двух сторон окружено пластинками компактного вещества. В костях свода черепа губчатое вещество называется диплоэ, diploe — двойное; оно залегает между наружной и внутренней пластинками компактного вещества, laminae externa et interna. В толще губчатого вещества костей свода черепа проходят диплоические каналы, canales diploici, проводящие венозные сосуды.
Некоторые кости черепа (лобная, решетчатая, клиновидная, верхняя челюсть) содержат воздухоносные пазухи, сообщающиеся с полостью носа. В то же время, ряд участков костей черепа образует утолщения – контрфорсы (лобно-носовой, альвеолярно-скуловой, крыловидно-небный, нижнечелюстной), являющиеся опорными местами этих костей (рис. 6, Д). Благодаря контрфорсам ослабляются механические толчки, испытываемые черепом.
В ячейках губчатого вещества кости и в костномозговой полости содержится костный мозг, medulla ossium. Различают красный костный мозг, medulla ossium rubra, и желтый костный мозг, medulla ossium flava.
Красный костный мозг обладает высокой функциональной активностью и способен формировать элементы крови. По мере развитии и роста организма красный костный мозг постепенно замещается желтым. Желтый костный мозг менее активен, выполняет резервную роль, но может активизироваться при определенных условиях, частично превращаясь в красный.
По мере развитии и роста организма красный костный мозг постепенно замещается желтым. Желтый костный мозг менее активен, выполняет резервную роль, но может активизироваться при определенных условиях, частично превращаясь в красный.
Осевой скелет, skeleton axialis, представлен костями черепа, позвоночного стоба и грудной клетки. Последние два отдела составляют кости туловища.
К костям добавочного скелета относят кости верхней конечности и кости нижней конечности.
Остеология — СтудИзба
Остеология
Функцию передвижения человека в пространстве выполняет опорно-двигательный аппарат. Это система костей и мышц, а также их соединений, которые образуют единый в функциональном отношении двигательный аппарат. В зависимости от функциональной значимости в нем различают пассивную и активную часть. К пассивной относятся кости и их соединения, к активной – мышцы.
Скелет (skeletos от греч. – высохший, высушенный) представляет собой комплекс костей, выполняющих опорную, защитную, локомоторную, формообразующую, преодолевающую силу тяжести функции. Функции, выполняемые скелетом, подразделяются на две большие группы — механические и биологические. К механическим функциям относятся защитная, опорная, локомоторная и рессорная. Биологическая функция связана с участием скелета в обмене веществ и в кроветворении. Скелет имеет билатеральную симметрию и сегментарное строение. В состав скелета входит более 200 костей. Из них 33-34 непарные (позвонки, крестец, копчик, грудина и некоторые кости черепа), остальные кости парные. Скелет подразделяется на осевой и добавочный. К осевому скелету относятся позвоночный столб (26 костей), череп (29 костей) и грудная клетка (25 костей), к добавочному скелету – кости верхних (64) и нижних (62) конечностей.
Функции, выполняемые скелетом, подразделяются на две большие группы — механические и биологические. К механическим функциям относятся защитная, опорная, локомоторная и рессорная. Биологическая функция связана с участием скелета в обмене веществ и в кроветворении. Скелет имеет билатеральную симметрию и сегментарное строение. В состав скелета входит более 200 костей. Из них 33-34 непарные (позвонки, крестец, копчик, грудина и некоторые кости черепа), остальные кости парные. Скелет подразделяется на осевой и добавочный. К осевому скелету относятся позвоночный столб (26 костей), череп (29 костей) и грудная клетка (25 костей), к добавочному скелету – кости верхних (64) и нижних (62) конечностей.
В своём развитии большинство костей скелета проходит 3 стадии: перепончатую, хрящевую, костную. На ранних стадиях развития скелет зародыша представлен хордой. С середины первого месяца внутриутробной жизни вокруг хорды появляется сгущение мезенхимы и развивается перепончатый скелет. Примерно в середине второго месяца развития мезенхима превращается в гиалиновый хрящ, а скелет получает название хрящевого.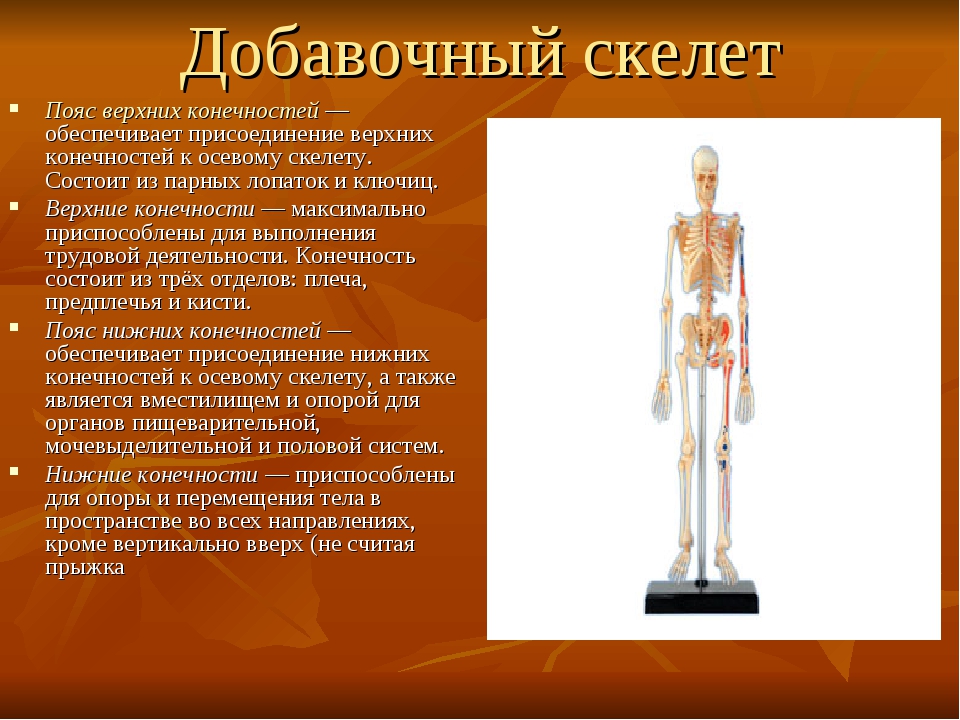 С конца второго — начала третьего месяца развития хрящевой скелет начинает окостеневать. Полное замещение хрящей костной тканью происходит к 20-25 летнему возрасту. С этого времени рост костей в длину прекращается. Кости свода черепа, лицевого черепа и части ключицы, в отличие от других костей скелета, в своем развитии проходят только 2 стадии — перепончатую и костную.
С конца второго — начала третьего месяца развития хрящевой скелет начинает окостеневать. Полное замещение хрящей костной тканью происходит к 20-25 летнему возрасту. С этого времени рост костей в длину прекращается. Кости свода черепа, лицевого черепа и части ключицы, в отличие от других костей скелета, в своем развитии проходят только 2 стадии — перепончатую и костную.
Кость — живой орган, в состав которого входят костная, хрящевая, соединительная ткани и кровеносные сосуды. Все кости составляют 18% общей массы тела. На поверхности каждой кости имеются выпуклости, углубления, борозды, отверстия, шероховатости. На них прикрепляются мышцы, сухожилия, фасции и связки. Возвышения над костями называются отростками, апофизами. На участках, к которым прилежит нерв или кровеносный сосуд, имеются борозды. В местах прохождения через кость сосуда или нерва образуются каналы, щели или вырезки. На поверхности каждой кости имеются отверстия, уходящие внутрь. Они получили название питательных отверстий.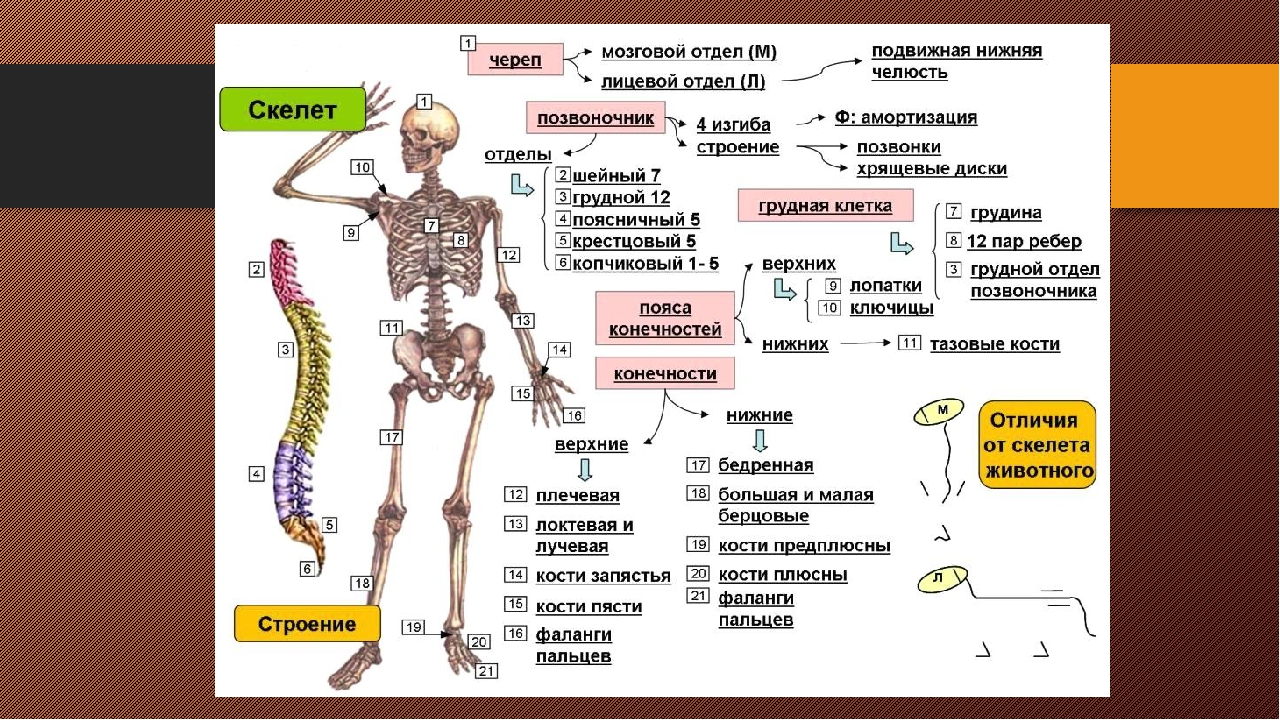
В состав костей входят органические (1/3) и неорганические вещества (2/3). Первые представлены преимущественно оссеином и оссемукоидом, вторые — солями кальция (51% фосфорнокислой извести). Oт наличия органических веществ зависит упругость кости, а от наличия неорганических соединений — ее твердость. В детском возрасте в костях больше оссеина, поэтому они обладают большей упругостью и редко ломаются. С возрастом происходит относительное (в процентах) уменьшение количества органических веществ и увеличение минеральных солей.
Кость снаружи покрыта надкостницей. Последняя отсутствует только на суставных поверхностях, которые покрыты суставным хрящом. Надкостница представляет собой тонкую, соединительнотканную пленку бледно-розового цвета. Она прикрепляется к кости с помощью соединительнотканных пучков — прободающих волокон. Надкостница имеет два слоя: наружный волокнистый (фиброзный) и внутренний костеобразующий (остеoгeнный). Она богата нервами и сосудами, которые участвуют в питании кости и её росте в толщину.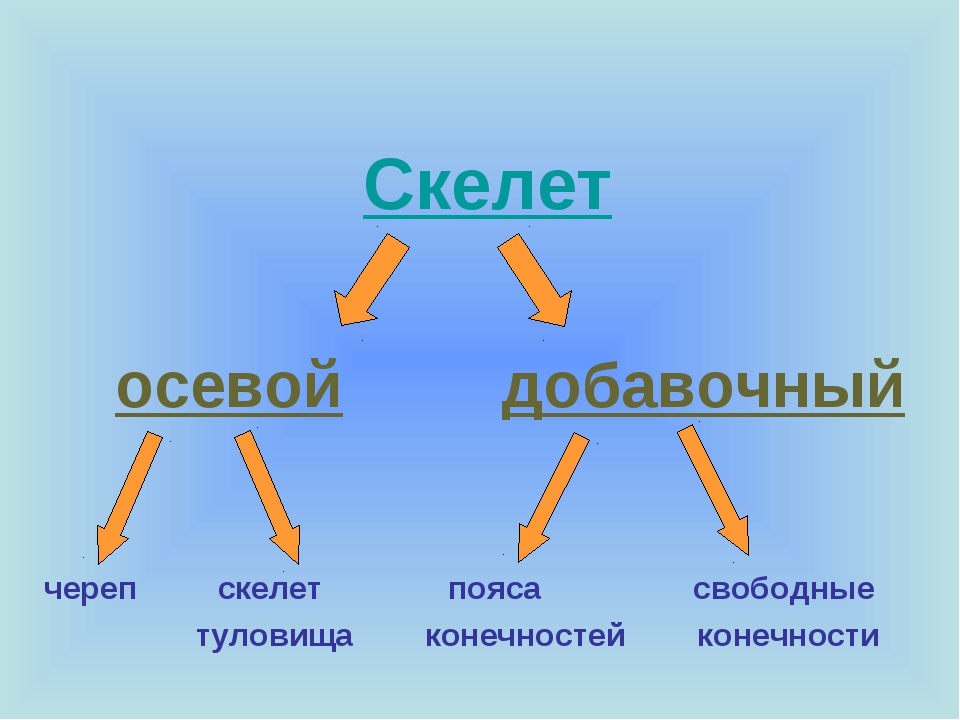 Питание осуществляется за счет кровеносных сосудов, проникающих в большом количестве из надкостницы в наружное компактное вещество кости через многочисленные питательные отверстия. Рост кости осуществляется за счет остеобластов, расположенных во внутреннем слое надкостницы.
Питание осуществляется за счет кровеносных сосудов, проникающих в большом количестве из надкостницы в наружное компактное вещество кости через многочисленные питательные отверстия. Рост кости осуществляется за счет остеобластов, расположенных во внутреннем слое надкостницы.
Структурной единицей кости является остеон — это система костных пластинок, концентрически расположенных вокруг центрального канала, содержащего сосуды и нервы. Он состоит из 5-10 цилиндрических пластинок, вставленных одна в другую. В центре каждого остеона проходит центральный (гаверсов) канал. Диаметр остеона составляет 0.3-0,4 мм. Цилиндры не прилегают друг к другу вплотную, а промежутки между ними заполнены интерстициальными (вставочными, промежуточными) пластинками. Остеоны располагаются не беспорядочно, а соответственно функциональной нагрузке на кость: в трубчатых костях параллельно длине кости, в губчатых — перпендикулярно вертикальной оси, в плоских костях черепа — параллельно поверхности кости и радиально.
Если Вам понравилась эта лекция, то понравится и эта — 29 — Локальные вычислительные сети.
Вместе с интерстициальными пластинками остеоны образуют основной средний слой костного вещества, который покрыт сверху наружными окружающими костными пластинками, а кнутри — внутренними окружающими костными пластинками. Из остеонов состоят более крупные элементы кости — перекладины костного вещества или трабекулы. Из трабекул складывается костное вещество двоякого рода: компактное и губчатое. Первое содержит плотно лежащие трабекулы, а второе – рыхло лежащие трабекулы, образующие между собою костные ячейки наподобие губки. Распределение компактного и губчатого вещества зависит от функциональных условий кости. Компактное вещество находится в тех костях, которые выполняют функцию опоры и движения (диафизы трубчатых костей, поверхности эпифизов). В местах, где при большом объеме требуется сохранить легкость и прочность под компактным находится губчатое вещество (эпифизы трубчатых костей).
Внутри костей, между костными пластинками губчатого вещества и в костных каналах трубчатых костей находится костный мозг, являющийся органом кроветворения и биологической защиты. Красный костный мозг представляет собой красную ретикулярную массу, в петлях которой находятся стволовые клетки, выполняющие функцию кроветворения, и клетки, выполняющие функцию костеобразования. Красный костный мозг пронизан нервами и кровеносными сосудами, питающими кроме костного мозга и внутренние слои кости. Кровеносные сосуды и кровяные элементы придают костному мозгу красный цвет. В процессе онтогенеза красный костный мозг заменяется на жёлтый, который обязан своим цветом жировым клеткам, из которых он и состоит. Во внутриутробном периоде и у новорожденных во всех костных полостях находится красный костный мозг в связи с высокой кроветворной и костеобразующей функциями. У взрослого человека красный костный мозг содержится только в ячейках губчатого вещества плоских костей (грудина, крылья подвздошных костей) и эпифизах трубчатых костей. В диафизах находится желтый костный мозг.
В диафизах находится желтый костный мозг.
В костной ткани располагаются костные клетки трёх видов: остеобласты, остеоциты, остеокласты. Остеобласты являются молодыми костными клетками многоугольной, кубической формы, богатые органоидами: рибосомами, комплексом Гольджи, элементами зернистой эндоплазматической сети. Клетки постепенно дифференцируются в остеоциты, при этом количество органоидов в них уменьшается. Межклеточное вещество, образуемое остеобластами, окружает их со всех сторон. Остеоциты – это уже зрелые многоотростчатые клетки, их отростки контактируют между собой. Клетки не делятся, органеллы в них развиты слабо. Остеокласты представляют собой крупные многоядерные клетки, разрушающие кость и хрящ. На своей поверхности имеют множество цитоплазматических выростов, покрытых цитоплазматической мембраной. Клетки богаты гидролитическими ферментами, митохондриями, лизосомами и вакуолями, хорошо выражен комплекс Гольджи.
По форме кости бывают трубчатые, губчатые, плоские, смешанные и воздухоносные. Трубчатые кости имеют форму трубки с костномозговым каналом внутри. У них различают удлиненную среднюю часть – диафиз – и утолщенные концы — эпифизы. На них располагаются суставные поверхности, покрытые хрящем и служащие для соединения с соседними костями. Участок между диафизами и эпифизами называется метафизом, благодаря которому осуществляется рост костей в длину. Диафизы построены из компактного, эпифизы из губчатого костного вещества, а покрыты сверху слоем компактного. Трубчатые кости делятся на длинные и короткие. Длина первых превышает все их размеры (бедренная, плечевая и локтевая). Короткие кости располагаются в пястье, плюсне, фалангах пальцев. Губчатые кости построены из губчатого вещества, покрытого тонким слоем компактного. Они бывают: длинные (ребра и грудина), короткие (кости запястья, предплюсны, позвонки), сесамовидные (надколенник, гороховидная кость, сесамовидные кости пальцев рук и ног). Сесамовидные кости развиваются в толще сухожилий и располагаются в тех местах, где большая нагрузка сочетается с большой подвижностью.
Трубчатые кости имеют форму трубки с костномозговым каналом внутри. У них различают удлиненную среднюю часть – диафиз – и утолщенные концы — эпифизы. На них располагаются суставные поверхности, покрытые хрящем и служащие для соединения с соседними костями. Участок между диафизами и эпифизами называется метафизом, благодаря которому осуществляется рост костей в длину. Диафизы построены из компактного, эпифизы из губчатого костного вещества, а покрыты сверху слоем компактного. Трубчатые кости делятся на длинные и короткие. Длина первых превышает все их размеры (бедренная, плечевая и локтевая). Короткие кости располагаются в пястье, плюсне, фалангах пальцев. Губчатые кости построены из губчатого вещества, покрытого тонким слоем компактного. Они бывают: длинные (ребра и грудина), короткие (кости запястья, предплюсны, позвонки), сесамовидные (надколенник, гороховидная кость, сесамовидные кости пальцев рук и ног). Сесамовидные кости развиваются в толще сухожилий и располагаются в тех местах, где большая нагрузка сочетается с большой подвижностью. Плоские кости (череп, лопатка, тазовые кости) выполняют защитную функцию и функцию опоры. Смешанные кости (основание черепа, позвонки) образуются при слиянии нескольких костей, различающихся по форме, строению и функции. Воздухоносные кости имеют в своем теле полость, выстланную слизистой оболочкой и заполненную воздухом (лобная, клиновидная, решетчатая кости и верхняя челюсть).
Плоские кости (череп, лопатка, тазовые кости) выполняют защитную функцию и функцию опоры. Смешанные кости (основание черепа, позвонки) образуются при слиянии нескольких костей, различающихся по форме, строению и функции. Воздухоносные кости имеют в своем теле полость, выстланную слизистой оболочкой и заполненную воздухом (лобная, клиновидная, решетчатая кости и верхняя челюсть).
Контрольная работа «Опорно-двигательная система»
Контрольная работа по биологии по теме «Опорно-двигательная система»
Класс: 8
Дата: 2.11.2016
Вариант 1
1. Какая кость в скелете человека является самой крупной?
1) большая берцовая
2) лучевая
3) бедренная
4) локтевая
2. Какую кость НЕ относят к скелету нижней конечности?
1) локтевая кость
2) пяточная кость
3) большая берцовая кость
4) малая берцовая кость
3. Что из перечисленного НЕ входит в состав скелета свободной верхней конечности?
Что из перечисленного НЕ входит в состав скелета свободной верхней конечности?
1) локтевая кость
2) плечевая кость
3) большая берцовая кость
4) лучевая кость
4. Что НЕ входит в скелет мозгового отдела черепа?
1) нижнечелюстная кость
2) затылочная кость
3) височная кость
4) теменная кость
5. Какой тканью выстланы головка и суставная ямка суставов?
1) хрящевой
2) нервной
3) гладкой мышечной
4) поперечнополосатой мышечной
6. В состав какого отдела скелета входит изображённое костное образование?
1) основания черепа
2) позвоночного столба
3) грудной клетки
4) пояса свободных нижних конечностей
7. Что из перечисленного характерно для скелета человека? Выберите три верных ответа из шести и запишите в таблицу цифры, под которыми они указаны.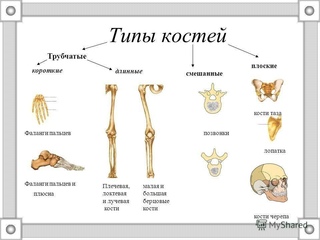
1) сводчатая стопа
2) прямой позвоночник без изгибов
3) позвоночник с S-образным изгибом
4) широкий чашевидный пояс нижних конечностей
5) сжатая с боков грудная клетка
6) массивные челюсти
8. Какие кости входят в состав поясов конечностей человека? Выберите три верных ответа из шести и запишите в таблицу цифры, под которыми они указаны.
1) рёбра 2) лопатки 3) плечевые
4) тазовые 5) ключицы 6) грудина
9. Установите соответствие между типом соединения костей и местом в скелете человека, где такое соединение существует.
МЕСТО СОЕДИНЕНИЯ КОСТЕЙ ТИП СОЕДИНЕНИЯ
А) крестцовые позвонки 1) неподвижное
Б) плечо и предплечье 2) полуподвижное
В) грудные позвонки 3) подвижное
Г) кости мозговой части черепа
Д) голень и стопа
Е) копчик
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам:
А | Б | В | Г | Д | E |
|
|
|
|
|
|
10. Назовите кости, обозначенные на рисунке буквами А и Б. Укажите, к какому отделу скелета их относят. Каково значение этого отдела скелета?
Назовите кости, обозначенные на рисунке буквами А и Б. Укажите, к какому отделу скелета их относят. Каково значение этого отдела скелета?
11. Назовите кости, отмеченные на рисунке буквами А, Б, В, Г. К каком отделу черепа они относятся?
12. Вставьте в текст «Типы соединения костей» пропущенные термины из предложенного перечня, используя для этого цифровые обозначения. Запишите в текст цифры выбранных ответов, а затем получившуюся последовательность цифр (по тексту) впишите в приведённую ниже таблицу.
ТИПЫ СОЕДИНЕНИЯ КОСТЕЙ
Скелет человека образован большим количеством костей, соединённых между собой. Там, где требуется максимальная прочность и защита, наблюдается __________ (А). Примером может служить соединение костей мозгового отдела черепа. Некоторые мелкие кости, например, позвонки, соединены друг с другом с помощью эластичного __________ (Б). Такой тип соединения называют __________ (В).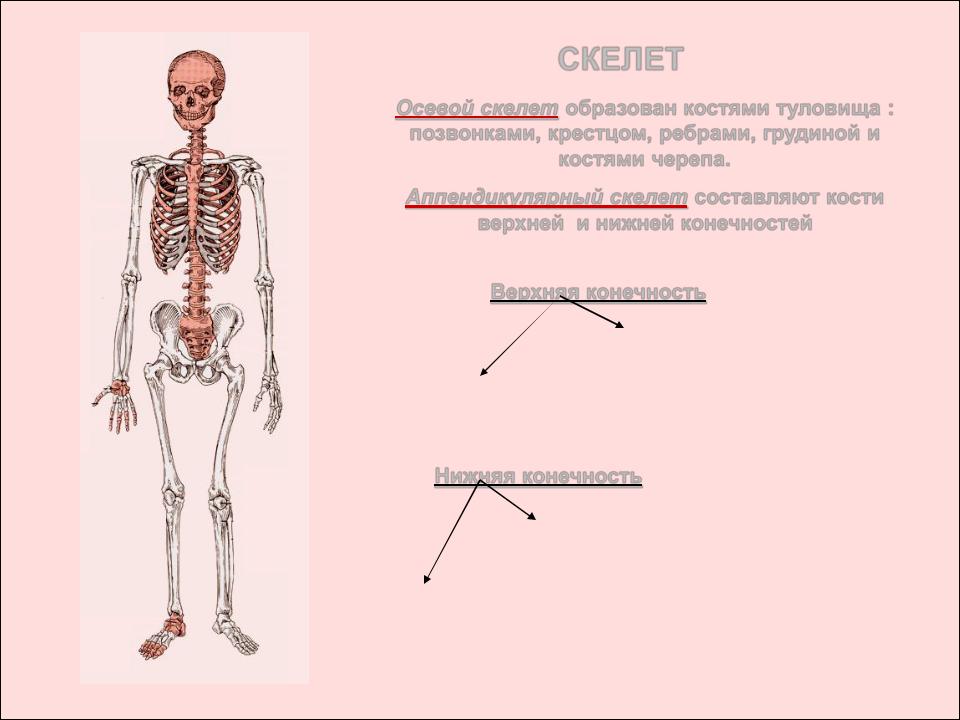 Наибольшая подвижность достигается при соединении костей с помощью __________ (Г).
Наибольшая подвижность достигается при соединении костей с помощью __________ (Г).
1) сустав
2) связка
3) сухожилие
4) неподвижное соединение
5) суставная сумка
6) полуподвижное соединение
7) мышца
8) хрящ
13. Дайте развернутый ответ. Как доказать, что неорганические вещества придают кости твердость?
Контрольная работа по биологии по теме «Опорно-двигательная система»
Класс: 8
Дата: 2.11.2016
Вариант 2
1. К скелету свободной верхней конечности относят
1) плюсну 2) лучевую кость 3) грудину 4) ключицу
2. Функцию питания и роста кости в толщину выполняет
1) жёлтый костный мозг
2) красный костный мозг
3) надкостница
4) губчатое вещество
3. Какая из перечисленных костей черепа является парной?
1) лобная 2) теменная 3) затылочная 4) нижняя челюсть
4. Лопатка входит в состав
Лопатка входит в состав
1) свободной верхней конечности
2) свободной нижней конечности
3) пояса верхней конечности
4) пояса нижней конечности
5. Какую долю от общего количества позвонков составляет число позвонков в шейном отделе позвоночника человека?
1) половину
2) больше половины
3) больше одной четверти
4) меньше одной четверти
6. Какая цифра обозначает кость предплечья?
1) 1 2) 2 3) 3 4) 4
7. Какие кости в скелете человека соединяются подвижно? Выберите три правильных ответа
1) височная и теменная
2) позвонки грудного отдела
3) нижняя челюсть с черепом
4) бедренная и тазовая
5) лобная и теменная
6) бедра и голени
8. Какие кости входят в состав поясов конечностей человека? Выберите три верных ответа из шести и запишите в таблицу цифры, под которыми они указаны.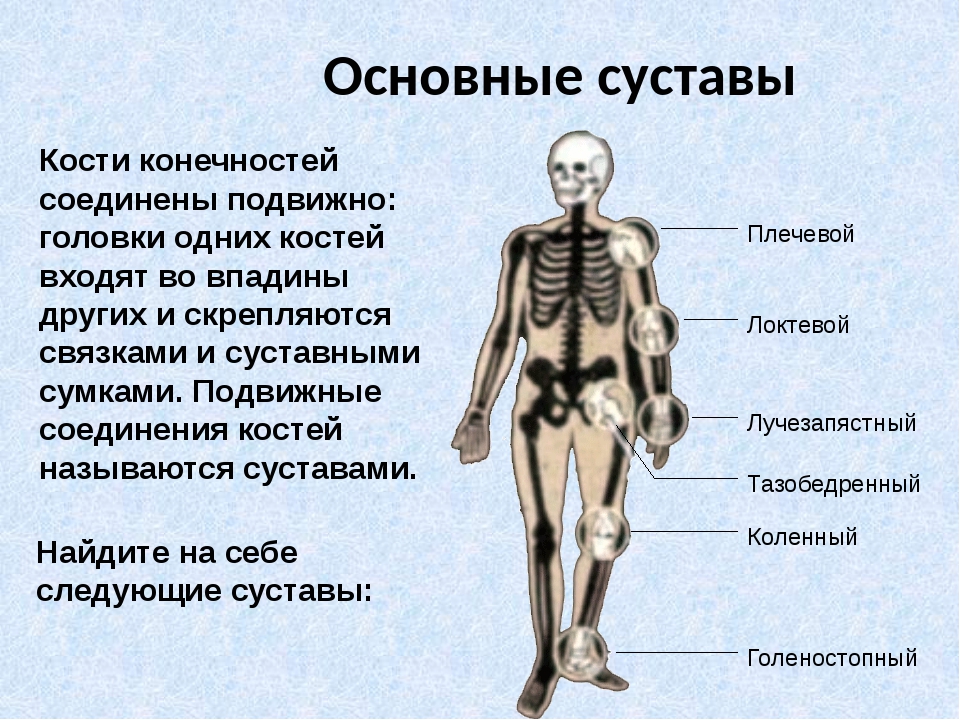
1) рёбра 2) лопатки 3) плечевые
4) тазовые 5) ключицы 6) грудина
9. Установите соответствие между типом соединения костей и местом в скелете человека, где такое соединение существует.
МЕСТО СОЕДИНЕНИЯ КОСТЕЙ ТИП СОЕДИНЕНИЯ
А) шейные позвонки 1) неподвижное
Б) запястье и предплечье 2) полуподвижное
В) поясничные позвонки 3) подвижное
Г) кости мозговой части черепа
Д) голень и стопа
Е) соединение ребер с грудиной
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам:
А | Б | В | Г | Д | E |
|
|
|
|
|
|
10. Назовите отделы позвоночника, указанные цифрами и количество позвонков в каждом отделе.
Назовите отделы позвоночника, указанные цифрами и количество позвонков в каждом отделе.
11. Назовите кости, обозначенные на рисунке цифрами 1,2,3,4. К какому отделу черепа они относятся?
12. Вставьте в текст «Строение трубчатой кости» пропущенные термины из предложенного перечня, используя для этого цифровые обозначения. Запишите в текст цифры выбранных ответов, а затем получившуюся последовательность цифр (по тексту) впишите в приведённую ниже таблицу.
СТРОЕНИЕ ТРУБЧАТОЙ КОСТИ
Скелет человека образует множество разнообразных костей, среди которых хорошо различимы трубчатые кости, образующие в основном скелет __________ (А). Тело трубчатой кости состоит из __________ (Б). В головке кости оно переходит в __________ (В), состоящее из перемычек, образующих ячейки, в которых находится __________ (Г), выполняющий кроветворную функцию.
1) губчатое вещество
2) надкостница
3) компактное вещество
4) жёлтый костный мозг
5) грудная клетка
6) свободная верхняя конечность
7) хрящ
8) красный костный мозг
13. Дайте развернутый ответ. Как доказать, что органические вещества придают кости эластичность, гибкость?
Дайте развернутый ответ. Как доказать, что органические вещества придают кости эластичность, гибкость?
Ключи
Вариант 1
1-3
2-1
3-3
4-1
5-1
6-2
По 1 баллу
7-134 3 балла
8- 245 3 балла
9
А | Б | В | Г | Д | Е |
1 | 3 | 2 | 1 | 3 | 1 |
6 баллов
10.
1) А – ключица; Б – лопатка
2) Пояс верхних конечностей
3) пояс верхних конечностей — опора, обеспечивает присоединение верхних конечностей к осевому скелету
4 балла
11. А- височная кость
А- височная кость
Б- затылочная кость
В-теменная кость
Г-лобная кость
Относятся к мозговому отделу черепа 5 баллов
12.А- 4 Б—8 В-6 Г-1
4 балла
13. Если прокаливанием кости удалить из нее все органические вещества, то кость, хотя и сохранит свою форму, станет хрупкой, но твердой. 1 балл
Итого – 32 балла
0-15 – 2
16-21- 3
22-27 -4
28-32 — 5
Вариант 2
1-2
2-3
3-2
4-3
5-4
6-3
7-346
8-245
9
А | Б | В | Г | Д | Е |
2 | 3 | 2 | 1 | 3 | 2 |
10.
1-шейный 7 позвонков
2-грудной – 12 позвонков
3- поясничный -5 позвонков
4- крестцовый – 5 позвонков
11.
1- нижняя челюсть
2- верхняя челюсть
3- носовая кость
4- скуловая кость
Относятся к лицевому отделу черепа
12.
А -6
Б-3
В- 1
Г- 8
13. Если кость опустить в раствор соляной или другой минеральной кислоты, то она станет гибкой, так как в ней останутся только органические вещества.
Источник: https://sdamgia.ru /
7.1 Подразделения скелетной системы — анатомия и физиология
Цели обучения
К концу этого раздела вы сможете:
- Обсудить функции костной системы
- Различать осевой скелет и аппендикулярный скелет
- Определите осевой каркас и его компоненты
- Определите остов отростка и его компоненты
Скелетная система включает в себя все кости, хрящи и связки тела, которые поддерживают и придают форму телу и структурам тела.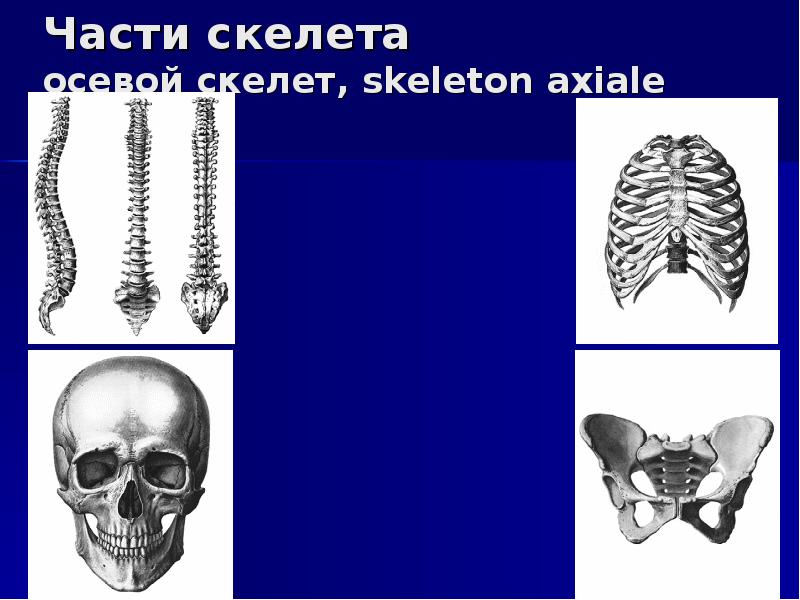 Скелет состоит из костей тела. У взрослых в скелете 206 костей. У молодых людей больше костей, потому что некоторые кости сливаются вместе в детстве и юности, образуя взрослую кость. Основные функции скелета — обеспечить жесткую внутреннюю структуру, которая может выдерживать вес тела против силы тяжести, и обеспечить структуру, на которую мышцы могут воздействовать, вызывая движения тела. Нижняя часть скелета предназначена для обеспечения устойчивости при ходьбе или беге.Напротив, верхний скелет имеет большую подвижность и диапазон движений, функции, которые позволяют поднимать и переносить предметы или поворачивать голову и туловище.
Скелет состоит из костей тела. У взрослых в скелете 206 костей. У молодых людей больше костей, потому что некоторые кости сливаются вместе в детстве и юности, образуя взрослую кость. Основные функции скелета — обеспечить жесткую внутреннюю структуру, которая может выдерживать вес тела против силы тяжести, и обеспечить структуру, на которую мышцы могут воздействовать, вызывая движения тела. Нижняя часть скелета предназначена для обеспечения устойчивости при ходьбе или беге.Напротив, верхний скелет имеет большую подвижность и диапазон движений, функции, которые позволяют поднимать и переносить предметы или поворачивать голову и туловище.
Каркас не только обеспечивает поддержку и движения тела, но и выполняет защитные и запасающие функции. Он защищает внутренние органы, включая головной и спинной мозг, сердце, легкие и органы малого таза. Кости скелета служат основным местом хранения важных минералов, таких как кальций и фосфат. Костный мозг, находящийся внутри костей, хранит жир и содержит ткань тела, вырабатывающую клетки крови.
Скелет подразделяется на два основных отдела — осевой и аппендикулярный.
Осевой скелет
Скелет подразделяется на два основных отдела — осевой и аппендикулярный. Осевой скелет образует вертикальную центральную ось тела и включает в себя все кости головы, шеи, груди и спины (рис. 7.2). Он служит для защиты головного и спинного мозга, сердца и легких. Он также служит местом прикрепления мышц, которые перемещают голову, шею и спину, и мышц, которые действуют через плечевые и тазобедренные суставы, чтобы двигать их соответствующие конечности.
Осевой скелет взрослого человека состоит из 80 костей, включая череп, позвоночник и грудную клетку. Череп образован 22 костями. С головой также связаны еще семь костей, включая подъязычную кость и слуховые косточки (по три маленькие кости в каждом среднем ухе). Позвоночный столб состоит из 24 костей, каждая из которых называется позвонком, а также крестца и копчика. Грудная клетка включает 12 пар ребер и грудину, уплощенную кость передней части грудной клетки.
Рис. 7.2. Осевой и аппендикулярный скелет. Осевой скелет поддерживает голову, шею, спину и грудь и, таким образом, образует вертикальную ось тела. Он состоит из черепа, позвоночника (включая крестец и копчик) и грудной клетки, образованной ребрами и грудиной. Аппендикулярный скелет состоит из всех костей верхних и нижних конечностей.
Аппендикулярный скелет
Аппендикулярный скелет включает все кости верхних и нижних конечностей, а также кости, прикрепляющие каждую конечность к осевому скелету.В аппендикулярном скелете взрослого человека 126 костей. Кости аппендикулярного скелета рассматриваются в отдельной главе.
Скелетная система: Осевой скелет
Эта статья, третья из серии из четырех частей, исследует осевой каркас. В четвертой части будет рассмотрен аппендикулярный скелет. В теле взрослого человека всего 206 костей (Tortora, Grabowski, 2003), которые разделены на две основные части (рис. 1). Осевой скелет состоит из костей черепа, позвоночника, грудины и ребер и состоит из 80 костей. Аппендикулярный скелет состоит из костей грудного, тазового и конечностей, а всего 126 костей. Осевой каркас состоит из:
Аппендикулярный скелет состоит из костей грудного, тазового и конечностей, а всего 126 костей. Осевой каркас состоит из:
- Череп (череп) — восемь;
- Череп (лицевой) — 14;
- Слуховые косточки (по три в каждом ухе — не показано) — шесть;
- Подъязычная (шейная кость, вспомогательная речь — не показана) — одна;
- Позвоночный столб — 26;
- Thorax (грудина) — одна;
- Грудь (грудная клетка) — 24.
Кости можно классифицировать на основе их формы (Таблица 1), причем некоторые формы костей связаны либо с осевым, либо с аппендикулярным скелетом.Позвоночник. Позвоночный столб (также называемый позвоночником, позвоночником или позвоночником) состоит из 33 костей неправильной формы (в детстве), называемых позвонками. Эти 33 кости делятся на пять категорий в зависимости от того, где они расположены в позвоночнике (рис. 2). Со временем некоторые более мелкие кости сливаются вместе, в результате чего у взрослых получается 26 позвонков (Bogduk, 2005; Tortora and Grabowski, 2003).
- Шейные позвонки — первые семь костей. Эти кости, расположенные в верхней части позвоночника, образуют шею и поддерживают голову.
- Грудные позвонки — следующие 12 костей. Они образуют задний якорь грудной клетки. Они больше шейных позвонков и увеличиваются в размерах сверху вниз.
- Поясничные позвонки — пять костей. Это самые большие кости позвоночного столба. Эти позвонки поддерживают большую часть веса тела и прикреплены ко многим мышцам спины.
- Крестец — треугольная кость чуть ниже поясничных позвонков, образует заднюю стенку тазового пояса. У детей он состоит из четырех или пяти крестцовых позвонков, которые срастаются во взрослом возрасте.
- Копчик — это низ позвоночника. Он состоит из трех-пяти костей, которые срослись вместе у взрослых. Многие мышцы соединяются с копчиком.
Череп Череп состоит из двух наборов костей — восьми черепных и 14 лицевых (основные из них показаны на рис. 3). Вместе они образуют большую полость черепа, которая защищает мозг. Череп также поддерживает структуру лица и содержит глазницы, носовые кости, височную кость, защищающую структуру уха, а также зубы и челюсти, позволяющие кусать и жевать (Waugh and Grant, 2006).Грудина и ребра Грудина — это длинная плоская кость в середине грудной клетки. Он состоит из манубрия, гладиолуса и мечевидного отростка. Он поддерживает ключицы и соединяется с хрящами первых семи пар ребер. У большинства людей есть 12 пар ребер, которые окружают грудную полость и защищают легкие, сердце и другие органы грудной клетки (у некоторых людей на одну пару больше или на одну пару меньше). Ребра соединены с позвонками. Пара с восьмой по десятую соединяется спереди с хрящом пары, расположенной выше, в то время как две нижние пары «плавают».Эластичность грудной клетки позволяет дышать. Грудина Различают три области (сверху вниз):
Череп также поддерживает структуру лица и содержит глазницы, носовые кости, височную кость, защищающую структуру уха, а также зубы и челюсти, позволяющие кусать и жевать (Waugh and Grant, 2006).Грудина и ребра Грудина — это длинная плоская кость в середине грудной клетки. Он состоит из манубрия, гладиолуса и мечевидного отростка. Он поддерживает ключицы и соединяется с хрящами первых семи пар ребер. У большинства людей есть 12 пар ребер, которые окружают грудную полость и защищают легкие, сердце и другие органы грудной клетки (у некоторых людей на одну пару больше или на одну пару меньше). Ребра соединены с позвонками. Пара с восьмой по десятую соединяется спереди с хрящом пары, расположенной выше, в то время как две нижние пары «плавают».Эластичность грудной клетки позволяет дышать. Грудина Различают три области (сверху вниз):
- Манубриум;
- Тело грудины — гладиолус;
- Мечевидный отросток.
Между рукояткой и телом грудины находится грудинный угол (хрящевой сустав, который действует как шарнир). Именно в этот момент грудина может двигаться вперед и назад, позволяя легким расширяться и сокращаться при дыхании (Marieb, 2003). Между телом и мечевидным отростком находится мечевидный отросток, который обычно используется в качестве ориентира при запуске сердечных сокращений в основных средствах жизнеобеспечения.Грудина состоит из высокососудистой губчатой ткани, которая покрыта тонким слоем компактной кости. Это наиболее толстая часть в области ладони между суставными фасетками ключиц. В верхней части грудины находится надгрудинная выемка,
Именно в этот момент грудина может двигаться вперед и назад, позволяя легким расширяться и сокращаться при дыхании (Marieb, 2003). Между телом и мечевидным отростком находится мечевидный отросток, который обычно используется в качестве ориентира при запуске сердечных сокращений в основных средствах жизнеобеспечения.Грудина состоит из высокососудистой губчатой ткани, которая покрыта тонким слоем компактной кости. Это наиболее толстая часть в области ладони между суставными фасетками ключиц. В верхней части грудины находится надгрудинная выемка,
— большое видимое углубление в месте соединения грудины с шеей. Ребра У большинства людей есть 12 пар ребер, хотя у некоторых есть 13-е, известное как шейные ребра — остатки ребер седьмого шейного позвонка, — а у некоторых только 11 пар. Ребра образуют грудную клетку, которая защищает легкие, сердце и другие органы грудной клетки.Они начинаются у грудных позвонков и изгибаются к передней части тела. Семь верхних пар ребер прикрепляются непосредственно к грудины с помощью гиалинового хряща. Нижние пять пар называются «ложными» ребрами — два нижних набора «плавают» и не прикрепляются к грудины (хотя они встроены в боковую мышечную стенку). Три набора ребер между ними и семью верхними ребрами прикреплены реберным хрящом к седьмому набору ребер и поэтому косвенно связаны с грудиной (Marieb, 2003).Промежутки между ребрами известны как межреберные промежутки.
Богдук, Н. (2005) Клиническая анатомия поясничного отдела позвоночника и крестца. Эдинбург: Черчилль Ливингстон.
Мариеб, Э. (2003) Анатомия и физиология человека (6-е изд). Менло-Парк, Калифорния: Издательство Бенджамина Каммингса.
Мартини, Ф.Х. (2005) Основы анатомии и физиологии (7-е изд.). Менло-Парк, Калифорния: Издательство Бенджамина Каммингса.
Tortora, G.J., Grabowski, S.R. (2003) Принципы анатомии и физиологии (10-е изд).Нью-Йорк, Нью-Йорк: Джон Уайли и сыновья.
Во, А., Грант, А. (2006) Анатомия и физиология здоровья и болезней (10-е изд.). Лондон: Черчилль Ливингстон.
Разнообразие | Бесплатный полнотекстовый | О шее, хоботе и хвосте: осевое разнообразие у позвоночных
1. Введение
Несмотря на замечательное анатомическое разнообразие, наблюдаемое у позвоночных, все они имеют осевой скелет, состоящий из сегментарных единиц, позвонков.Разнообразие, наблюдаемое в количестве, размере и специфических характеристиках различных позвонков, а также в их специфической сборке в позвоночном столбе, отражает различные стратегии адаптации к широкому разнообразию экологических ниш, завоеванных позвоночными. В этом обзоре я буду обсуждать различные аспекты разнообразия тел позвоночных, в основном сосредотачиваясь на осевом скелете и стремясь дать отчет о возможных механизмах развития некоторых характерных особенностей, которые часто служат определяющими признаками данного вида.
Сомиты представляют собой первый признак сегментации по основной оси тела. Это парные эпителиальные структуры по обе стороны от развивающегося спинного мозга, которые образуют позвонки и ребра, дерму тела и скелетные мышцы тела и конечностей [1]. Сомиты генерируются последовательно от рострального к каудальному развитию в процессе сомитогенеза, происходящего на каудальном конце оси растущего тела. Сомитогенез подробно описан во многих прекрасных обзорах [2, 3, 4, 5] и не будет здесь обсуждаться.Общее количество сомитов и, следовательно, позвонков, образующихся во время эмбрионального развития, широко варьируется среди позвоночных, но остается довольно постоянным в пределах данного вида. Эта изменчивость представляет собой один из основных источников морфологического разнообразия филогении позвоночных. Следует отметить, что у амниотов вклад сомитов в формирование позвонков начинается с пары сомитов 5, так как большее количество передних пар сомитов участвует в развитии затылочной кости [6]. У анамниотов только первые три пары сомитов вносят вклад в элементы черепа [7,8].Это важное соображение, так как оно напрямую влияет на оценку воздействия, которое дифференциальная региональная регуляция дифференцировки сомитов оказывает на позвоночную идентичность. Морфологически сходны сомиты, когда они впервые образуются во время сомитогенеза, но порождают позвонки с различными индивидуальными особенностями, которые, опять же, в высшей степени сохраняются в пределах данного вида, но значительно различаются по филогении позвоночных. Хотя в целом уникальные анатомические особенности могут быть приписаны каждой позвоночной единице, их подмножества имеют общие черты, которые привели к их классификации на пять различных групп, а именно шейные, грудные, поясничные, крестцовые и хвостовые, которые могут быть применены к большинству видов позвоночных.Распределение позвонков среди этих различных групп, обычно известное как осевая формула [9], является еще одним из основных источников разнообразия среди клад позвоночных. Большое количество экспрессионных и генетических исследований показали, что как индивидуальная, так и региональная идентичность различные позвонки в основном являются результатом активности генов семейства Hox. Учитывая их важную роль в создании морфологического разнообразия, я кратко опишу их основные особенности, но обширные обзоры их функции можно найти в другом месте [10,11,12].У позвоночных различные члены семейства Hox генов расположены в кластерах, которые, как полагают, возникли в результате последовательных дупликаций генома во время появления позвоночных [13]. Количество кластеров варьируется в зависимости от филогении позвоночных. Например, млекопитающие, которые часто служат основным ориентиром, содержат четыре кластера, названные от A до D [13], тогда как костистые рыбы, такие как рыба данио (Danio rerio), имеют семь кластеров [14]. Отдельные гены Hox (39 у млекопитающих) классифицируются на 13 паралоговых групп в соответствии с гомологией последовательностей и их положением в кластере, распределены от 1 до 13 в 3’ – 5 ’ориентации внутри кластера [13].Геномное расположение Hox-генов влияет на их пространственную и временную экспрессию во время эмбрионального развития: паралоги с меньшим числом активируются первыми в передних эмбриональных областях, за ними почти последовательно следуют Hox-гены с увеличивающимся числом паралогов в более задних эмбриональных областях по мере того, как эмбрион расширяется на его хвостовой конец [13]. Этот тип регуляции приводит к экспрессии различных комбинаций Hox генов на разных осевых уровнях, что, как полагают, играет существенную роль в генерации различных сегментарных идентичностей вдоль основной оси тела [13].2. Шея: различные решения схожих проблем
Размер шеи существенно различается у разных видов позвоночных. Основным источником этой вариации является количество позвоночных единиц, расположенных в этой области осевого скелета. Амфибии и змеи занимают один конец спектра, содержащий от одного до трех шейных позвонков, в то время как птицы помещаются на другом конце, содержащие переменное, но всегда довольно большое количество шейных позвонков, за которое борются лебеди с их 25 единицами (Рисунок 1). [7,15,16,17].Млекопитающие расположены в середине шкалы, в основном придерживаясь правила семи шейных позвонков, за исключением ламантинов и ленивцев [18]. Происхождение этой изменчивости восходит к Hox-генам. Сравнительные исследования экспрессии гена Hox у эмбрионов видов с разным количеством шейных позвонков, включая змей (Pantherophis guttatus), ящериц (Aspidoscelis uniparens), слепых кишок (Ichthyophis cf. kohtaoensis), кур (Gallus gallus), гусей ( мыши (Mus musculus), аллигаторы (Alligator mississippiennsis) и лягушки (Xenopus laevis) [15,17,19,20,21] показали сильную корреляцию между уровнем сомита, при котором гены Hox групп паралогов 5 и 6 активируются и анатомическое положение перехода шеи в туловище в позвоночнике.Экспериментальное подтверждение этой гипотезе было предоставлено генетическими экспериментами на мышах. В частности, было показано, что передняя экспансия экспрессии Hoxb6 предвосхищает переход в структуры туловища с сопутствующим уменьшением размера шеи [22]. Напротив, глобальная инактивация генов Hox группы 5 или группы 6 перемещает переход от шеи к туловищу назад на один или два сегмента [23]. Хотя эти нулевые фенотипы выглядят несколько мягче, чем можно было бы ожидать, вполне вероятно, что это происходит из-за функциональной избыточности между генами в этих двух группах паралогов.В самом деле, трансгетерозиготные животные по генам Hoxb6 и Hoxb5 указали на их функциональную кооперацию в процессах формирования скелетного паттерна [24]. Разнообразие в количестве шейных позвонков может, таким образом, отражать видоспецифические вариации в механизмах активации генов Hox5 и Hox6 паралогов. До сих пор большая часть наших знаний о механизмах, регулирующих экспрессию гена Hox, получена в результате генетических экспериментов на мышах и эмбриональных стволовых клетках, и поэтому невозможно предоставить должным образом задокументированный отчет о дифференциальных механизмах активации между видами с разными шейными позвонками. подсчитывает.Однако идентификация регуляторных механизмов, включающих структуру хроматина как критический компонент в последовательной активации Hox генов [12,25,26,27,28], дает некоторые указания на возможные механизмы этой дифференциальной регуляции. В частности, было показано, что 3 ‘и 5’ части Hox-кластеров встроены в два соседних топологически ассоциирующих доменов (TAD) [29,30,31], трехмерные территории хроматина, маркирующие геномное пространство, которое, как считается, допускает взаимодействия. между генами и регуляторными элементами [32,33].Активация гена Hox контролируется регуляторными ландшафтами в каждом из двух TAD [12]. Последовательная активация Hox, таким образом, требует, чтобы в определенный момент регуляция транскрипции переключалась с элементов в 3 ’TAD на элементы в 5’ TAD [28,30]. Этот переключатель может играть важную роль в профилях активации Hox5 и Hox6, наблюдаемых у разных видов. В частности, граница между двумя Hox-ассоциированными TADs расположена на уровне паралогов Hox5 и Hox6, и было показано, что целостность этой границы важна для гарантии правильной регуляции Hox генов [29,34].Подробные эксперименты на мышах также показали, что активация Hoxa5 и Hoxa6 связана с изменением регуляции Hox с WNT- на Cdx-зависимые механизмы [35]. Таким образом, возможно, что разница в количестве шейных позвонков является результатом вариаций в механизмах, вызывающих переключение между регуляторными ландшафтами TAD, которые могут быть результатом различных внутренних свойств факторов, связанных с Cdx или WNT, или их нижестоящих эффекторов. Чтобы понять, задействованы ли те или иные механизмы в этом процессе, потребуется прямая оценка у разных видов.Не все вариации размера шеи связаны с разным количеством позвонков, отнесенных к этому сегменту осевого скелета. Млекопитающие, например, содержат фиксированное количество шейных позвонков, но их относительный размер шеи сильно варьируется в зависимости от клады. Исследование с участием 352 видов показало, что, за некоторыми исключениями, относительный размер шейного отдела позвоночника млекопитающих обратно пропорционален массе тела животного [36]. Жирафы представляют собой наиболее известное исключение из этой обратной аллометрии и представляют парадигматический пример, иллюстрирующий образование длинных шеек посредством значительного увеличения размеров их шейных позвоночных единиц [37].Важно отметить, что шейные позвонки являются единственными элементами осевого скелета жирафа, размер которых значительно отличается от размера, наблюдаемого в гомологичных костях родственных видов с более короткой шеей [37] (Рисунок 1), что указывает на очень точное региональное ограничение механизмов, ответственных за увеличенный размер позвонков. Несмотря на то очарование, которое вызывали у жирафов длинные шеи за долгие годы, мы все еще далеки от убедительного описания механизмов, порождающих эту замечательную анатомическую особенность.Некоторые возможные объяснения были предложены на основе анализа геномной последовательности двух видов жирафов (Giraffa camelopardalis и Giraffa tippelskirchi) [38,39]. Одна из таких гипотез возникла из значительных специфичных для жирафов нуклеотидных изменений, наблюдаемых у некоторых членов путей NOTCH, WNT и FGF. Учитывая существенное участие этих путей во временной динамике сомитогенеза и, в конечном итоге, размер сомитов [4], было высказано предположение, что регуляторный баланс между этими путями во время сомитогенеза жирафов д. Продуцировать более крупные сомиты.Особое внимание было уделено передаче сигналов FGF, потому что экспериментальные изменения его активности во время сомитогенеза курицы сопровождались значительными изменениями в размере сомита [40]. Это указывает на возможность того, что функциональные модификации в Fgfrl1, одном из наиболее дивергентных генов у жирафов [38], д. Скорректировать динамику сомитогенеза способом, который способствует продукции более крупных сомитов. Недавние генетические данные, однако, связали специфические для жирафов особенности Fgfrl1 не с размером шейных позвонков, а с их замечательной устойчивостью к высокому кровяному давлению [39].Альтернативный механизм образования больших позвонков включает в себя дифференцированный рост скелетных элементов, происходящих из сомитов одинакового размера, однако, независимо от конкретного механизма, полное понимание происхождения больших шейных позвонков жирафа также должно учитывать его точные региональные особенности. специфичность. Hox гены, и особенно те из паралогов групп 3 и 4, являются главными кандидатами на выполнение этой роли, поскольку они, как было показано, формируют анатомию и размер шейных позвонков у мышей [41,42,43].Значительные изменения последовательности в Hox-генах жирафов по сравнению с okapi (Okapia johnstoni) были обнаружены только в Hoxb3 [38], таким образом, маловероятно, что внутренние свойства этих генов оправдывают образование больших шейных позвонков. Hox-гены могут по-прежнему контролировать избирательный рост шейных позвонков посредством значительных функциональных изменений в их последующих эффекторах. Например, учитывая, что сигнальные пути NOTCH, WNT и FGF также участвуют в процессах роста и ремоделирования скелета [44,45,46], возможно, что специфические для жирафов изменения в этих путях увеличивают их способность способствовать росту скелета.Если какой-либо из этих факторов является частью специфических регуляторных сетей ниже белков Hox3 или Hox4, то активность этих Hox генов в эмбрионе жирафа может способствовать региональному специфическому увеличению шейных позвонков. Эта или другие возможные гипотезы происхождения скелетного рисунка шеи жирафов потребуют прямого экспериментального подтверждения. Развитие шейных позвонков у китообразных шло по пути, противоположному таковому у жирафов: у них также есть семь позвоночных единиц, но они очень маленькие и сросшиеся [18 ], структурная характеристика, которая, как считается, облегчает плавание, обеспечивая стабильность за счет подвижности головы.Происхождение этого рисунка шейки матки по существу неизвестно, но может быть связано с вариациями в процессах, аналогичных процессам образования длинных элементов шейки матки у жирафов, но с противоположным значением. Независимо от механизмов, наличие одинакового количества шейных позвонков у жирафов и китообразных, независимо от их совершенно разных относительных размеров у взрослого животного, подчеркивает неспособность млекопитающих адаптироваться к давлению окружающей среды, требующему изменения размера шеи путем изменения количества позвоночных элементов, полагаясь вместо этого на альтернативные механизмы.Действительно, правило семи шейных позвонков, по-видимому, нарушают только ламантины и ленивцы [18,47], и даже для ленивцев недавние данные показывают, что, хотя у них есть 8–10 позвонков перед первым сегментом, связанным с грудным ребром, критерии окостенения указывают на что только первые семь позвонков соответствуют параметрам шейки матки [48], что заставляет ленивцев снова придерживаться правила семи шейных позвонков. Есть два основных направления мысли, объясняющих это замечательное ограничение развития. Один из них связывает это с наличием мышечной диафрагмы, характерной для млекопитающих особенностью, играющей фундаментальную роль в их механике дыхания и, как полагают, позволила развить отличительные физиологические особенности млекопитающих, включая повышенную температуру тела и высокую скорость метаболизма в состоянии покоя [47 , 49].Модуль развития, который генерирует и направляет пути миграции мышечных предшественников для диафрагмы, тесно скоординирован с теми, которые направляют развитие мышц передних конечностей, включая смежные сомиты предполагаемой шейной области [50]. Было высказано предположение, что тесные функциональные связи между этими двумя процессами снижают способность системы допускать значительные изменения без фатального вмешательства в фундаментальные особенности физиологии млекопитающих [49]. Наблюдение, что диафрагма ламантинов отличается по положению от диафрагмы у других млекопитающих, вероятно, связанное с уникальным положением их легких вдоль спины животных [51], по-видимому, в некоторой степени подтверждает гипотезу, основанную на диафрагме.Другая линия мысли проистекает из наблюдения увеличения числа пороков развития, потенциально связанных с измененной экспрессией гена Hox, таких как эктопия шейных ребер, у детей, страдающих врожденным раком [52]. Затем было высказано предположение, что у млекопитающих изменения в экспрессии гена Hox, необходимые для изменения количества шейных позвонков, будут связаны с повышенным образованием злокачественных новообразований, которые будут мешать исправлению новых особенностей развития, что с меньшей вероятностью произойдет у птиц или рептилий. учитывая их явно повышенную устойчивость к раку по сравнению с млекопитающими [52].Следует отметить, что, хотя вариация длины шеи у птиц в значительной степени зависит от количества шейных позвонков, которое варьируется от 10 до 25, также было показано, что размер отдельных элементов также влияет на разнообразие длины шеи. наблюдается во всей птичьей филогении, которая, в отличие от того, что наблюдается у млекопитающих, показывает общую положительную аллометрию с размером тела [53].3. Размер и форма туловища: крайние различия
В целом можно считать, что туловище — это область тела, в которой находится большинство органов сердечно-сосудистой, пищеварительной, выделительной и репродуктивной систем, что примерно соответствует грудопоясничным и крестцовым отделам. области осевого скелета.Длина туловища у позвоночных широко варьируется: от нескольких позвонков у лягушек до более 300 у некоторых змей.
Обычно переход от шеи к туловищу в осевом скелете отмечается положением первого позвонка, содержащего подвижное ребро, прикрепленное к грудины. Следовательно, положение первого позвонка туловища и длина шеи — две стороны одного и того же процесса. Тогда возникает новый вопрос: что определяет, где заканчивается ствол? Как обсуждалось в отношении размера шеи, корреляция между активацией специфических групп Hox паралогов и анатомическими переходами в осевом скелете, вместе с ролью этих генов в определении сегментарных идентичностей, предполагает их возможное участие в качестве ключевых детерминант изменчивости размера туловища среди позвоночных.Эти корреляции также вывели эти гены на центральное место в поисках происхождения строения тела змеи, что стало основным двигателем исследований механизмов, лежащих в основе анатомического разнообразия позвоночных. В самом деле, открытие, что активация Hox паралогов групп 10-13 сильно задерживается у змей по сравнению с другими позвоночными, коррелируя с концом структур туловища [17,20], согласуется с этой гипотезой. Сходным образом, более задняя активация тех же Hox паралогов у мутантных мышей с удлиненным хоботом также указывает на связь между Hox генами и размером туловища [54,55,56].Однако обширные генетические данные у мышей, по-видимому, опровергают причинную роль Hox-генов в этом процессе, поскольку основные изменения в экспрессии Hox-генов, будь то их единственная или комбинированная инактивация, или усиление функциональных подходов, не привели к значительным изменениям в туловище. размер, оцениваемый по расстоянию между зачатками передних и задних конечностей, несмотря на то, что в некоторых случаях вызывает серьезные изменения позвоночника вдоль передней задней оси [10]. В отличие от того, что наблюдалось для генов Hox, генетическая инактивация различных факторов, участвующих в Путь передачи сигналов TGFβ приводил к значительному расширению области туловища, часто за счет развития более каудальных областей тела (рис. 2).Коллективный анализ этих экспериментов выявил Gdf11, действующий через рецепторы Tgfbr1 и Acvr2b, в качестве ключевого активатора перехода от туловища к хвосту и, таким образом, основного игрока в определении длины ствола [54,55,57,58,59, 60,61,62]. В то время как большинство генетических экспериментов было выполнено на мышах, недавние данные показывают, что роль Gdf11 в контроле конца развития туловища, вероятно, является общей чертой филогении позвоночных. В частности, было показано, что его экспрессия тесно коррелирует с положением перехода к развитию хвоста у разных видов позвоночных [56,63].Кроме того, экспериментальная модуляция активности Gdf11 была способна изменить положение перехода туловища к хвосту у животных, отличных от мышей [63,64]. Также сообщалось о расширении области туловища у мышей, лишенных всех трех генов miR196 [65]. ]. Интересно, что экспериментальная активация или подавление miR196 во время эмбрионального развития рыбок данио вызывает небольшие сокращения или экспансии туловища, соответственно, дополнительно связывая miR196s с регуляцией размера туловища [66]. Гены miR196 расположены в кластерах Hox и, как сообщается, способны контролировать экспрессию некоторых генов Hox [67].Hox гены также обнаруживают аномальные паттерны у глобальных мутантов miR196, которые коррелируют с изменениями в осевом скелете [65]. Однако неясно, происходят ли эти изменения в паттернах экспрессии Hox из-за прямой регуляции с помощью miR196 и являются ли изменения в экспрессии гена Hox причиной или следствием расширенной области ствола у мутантов miR196. В этом отношении есть некоторые указания на то, что miR196s м. Контролировать путь Gdf11, поскольку глобальный мутантный фенотип miR196 напоминает особенности, наблюдаемые у некоторых гетерозиготных Gdf11 мышей, а экспрессия Gdf11 была значительно снижена у мутантных эмбрионов miR196 [65].Потребуется прямой экспериментальный анализ, чтобы определить, действительно ли miR196 и Gdf11 функционально связаны, а также природу такого возможного соединения. Хотя вмешательство в передачу сигналов Gdf11 или инактивация семейства miR196 изменили размер ствола, они не оказали существенного влияния на общую протяженность. оси тела [54,65]. Эти наблюдения показывают, что изменений в этих действиях недостаточно, чтобы сформировать типичный план тела змеи. Действительно, сочетание экспериментального и модельного подходов привело к идентификации двух дополнительных процессов, которые могут играть важную роль в производстве удлиненных хоботов змей.Один из этих процессов состоит из продолжительного периода активности Oct4 (Pou5f1) в эпибласте эмбрионов змеи, что, скорее всего, является результатом серьезных изменений в регуляции его экспрессии [56], что может быть ключом к обеспечению непрерывного производства ткани. требуется, чтобы продлить ось корпуса через область туловища. Возможность прямого экспериментального вмешательства в эмбрионы змей очень снижена, что ограничивает степень прямой проверки, которая может быть проведена с этими эмбрионами, но трансгенные эксперименты на мышах предоставили некоторую поддержку этой гипотезе [56].Второй процесс, который мог способствовать большому количеству позвонков, характерных для змей, — это ускоренный сомитогенез, который приводит к генерации большего числа сегментов, образующих туловищные позвонки [16]. Эти сомиты меньше, чем у эмбрионов других видов позвоночных, но образуют позвонки с размерами, эквивалентными размерам других позвоночных аналогичного размера, и, следовательно, расширение каждого отдельного сегмента во время дифференцировки приводит к значительному глобальному удлинению оси их основного тела.Базовая структура тела, генерируемая передачей сигналов Oct4 и Gdf11 (нельзя исключать участие дополнительных факторов), приводит к образованию структур тела посредством скоординированного действия нижестоящих факторов, которые гарантируют производство тканей, соответствующих различным областям тела. Это стадия, на которой вклад Hox генов существенен благодаря регулированию компоновки позвоночных паттернов, соответствующих специфическим требованиям различных регионов тела вдоль основной оси тела [68].Специфические атрибуты связанного с туловищем скелета неоднородны, значительно варьируются в зависимости от филогении позвоночных и часто корректируются для облегчения определенных характеристик физиологии или поведения животных. Хороший тому пример — млекопитающие. Их скелет туловища является наиболее четко регионализированным среди позвоночных, обычно состоит из закрытой грудной клетки (грудной области), состоящей из ребер, соединяющих позвонок с грудиной по средней линии вентральной части, за которыми часто следует поясничная область без ребер перед крестцом, которая отмечает конец. стволовых конструкций.Число грудопоясничных позвонков у млекопитающих не такое строгое, как на шее, но все же остается в относительно небольшом диапазоне — от 19 у копытных животных до 24 у африканских слонов [69]. Интересно, что утверждалось, что специфическая анатомия и эволюционная консервация позвоночной структуры этой области являются результатом адаптации к типу передвижения вида. Считается, например, что наличие гладкой и гибкой поясничной области важно для обеспечения быстрых и подвижных движений, что не так важно для более медленных и крепких животных [69].Действительно, у слонов отсутствует поясничная область без ребер, характерная для большинства других млекопитающих. Птицы также имеют уникальную конфигурацию осевого скелета туловища, приспособленную для обеспечения их крыльям и ногам сильной опоры, необходимой для облегчения их механики полета и двуногой ходьбы. Он состоит из компактной грудной клетки с уменьшенным количеством ребер, за которыми следует синсакрум, структура, образованная слиянием их поясничных, крестцовых и последних грудных звеньев [70], которые также сливаются с тазовым поясом, обеспечивая прочный крепление для задних конечностей, необходимое для двуногой ходьбы, с центром тяжести, ростральным по отношению к положению ног.Змеи демонстрируют совершенно другой тип осевого скелета туловища, состоящий из большого количества ребер, содержащих позвонки, простирающихся до анальной области. Количество туловищных позвонков варьируется как у разных видов змей, так и часто внутри одного вида, демонстрируя половой диморфизм, причем самки обычно содержат большее количество туловищных позвонков, которые производят более длинные хоботы, что, как считается, способствует плодовитости [71]. Важно отметить, что у змей нет грудины. Следовательно, в отличие от млекопитающих, птиц или ящериц с конечностями, у змей отсутствует замкнутая грудная клетка, и их ребра свободно оканчиваются в мягких тканях в брюшной части тела, что может иметь важное значение для обеспечения экстремального расширения тела, необходимого для их кормления. стратегии.Субрегионализация осевого скелета туловища в большой степени зависит от дифференциальной экспрессии Hox генов и особенно от генов паралогов 7-11, как было выявлено генетическими экспериментами [10,11]. Парадигматический пример представлен генами Hox10, которые необходимы для генерации безрубчатого поясничного домена у млекопитающих [72,73]. Сопоставление данных экспрессии с паттернами скелета у видов, не относящихся к млекопитающим, предполагает сохранение участия гена Hox в субрегионализации осевого скелета туловища [17,20] за пределами млекопитающих.Непредсказуемой иллюстрацией этой идеи послужили эксперименты на змеях, где точный морфометрический анализ отдельных позвонков выявил существование тонкой субрегионализации вдоль передней задней оси, примерно соответствующей конкретным доменам экспрессии гена Hox, что ставит под сомнение классическую концепцию отсутствия регионализации. в осевом скелете змеи [74]. Однако эксперименты со змеями также показали, что активность Hox-паттерна скелетных элементов не может быть просто предположена на основании их экспрессии и что дополнительные скрытые слои могут формировать их функцию, которая может довольно различаться у разных видов.В частности, было обнаружено, что гены Hoxa10 и Hoxc10 змеи хорошо экспрессируются в сомитной области, образующей реберно-содержащие позвонки туловища [17,20], что противоречит известной роли генов Hox10 в возникновении без ребер поясничного отдела позвоночника. позвонки, выявленные в ходе генетических экспериментов на мышах [72,73]. Было показано, что это очевидное несоответствие является результатом полиморфизма одного нуклеотида в важном Hox-чувствительном энхансере, который делает его нечувствительным к активности блокирования ребер белков Hox10 [75], что позволяет телам позвонков в области каудального туловища приобретать поясничный отдел. черты типа, при этом оставаясь прикрепленными к взрослым ребрам.Интересно, что тот же полиморфизм в гомологичном энхансере слонов, по-видимому, является источником наличия ребер в позвоночной области, соответствующей поясничной области у других млекопитающих [75]. Хотя предполагается, что гены Hox также могут играть важную роль. роль в производстве специфического для птиц осевого скелета туловища [15], его явное отклонение от паттернов мышей затрудняет понимание, какова эта роль. Генетические эксперименты на мышах показали существенную роль Hox paralog group 11 в создании характерного слияния крестцовых позвонков у млекопитающих [72].Это предполагает участие этих генов в генерации синсакрума. Хотя данные по экспрессии у куриных эмбрионов с хорошо развитыми задними конечностями помещают активацию гена Hox11 позади уровней сомитов, образующих пояснично-крестцовую область [15], на более ранних стадиях их экспрессия перекрывается с областью, которая дает позвонки, участвующие в синсакруме [76 ]. У мышей было показано, что активность формирования паттерна некоторых Hox генов обеспечивается в пресомитной мезодерме, когда формируются сомиты, а не в самих сомитах [73].Следовательно, если тот же принцип применим к генам Hox11 у куриных эмбрионов, эти гены также могут играть значительную роль в формировании синсакрума. Черепахи заслуживают особого упоминания в контексте осевого скелета туловища, поскольку их панцирь представляет собой крайнюю модификацию их ребра, их тела позвонков и связанные с ними мышечные и дермальные производные сомитов [77]. В частности, во время ранних стадий дифференцировки сомитов черепах их зачатки ребер растут латерально, вероятно, управляемые сигналами от специализированной структуры в боковой стенке эмбриона, в конечном итоге покрывая развивающиеся зачатки конечностей дорсально [77,78].Развитие мышечных предшественников также нарушается, вероятно, из-за специфической для черепах модификации гена Myf5 [79], и межреберные мышцы между ребрами заменяются кальцинированной дермальной структурой, которая завершает структуру панциря [80].4. Хвост — это не просто хвост
Хвост представляет собой область тела, простирающуюся от основной оси тела кзади до ануса. При рассмотрении осевого скелета основными компонентами хвоста являются хвостовые позвонки. У амфибий размер хвоста у млекопитающих и рептилий напрямую связан с их количеством хвостовых позвонков, которое может варьироваться от нескольких слитых элементов, таких как копчик человека, до более сотни у некоторых видов ящериц, таких как длиннохвостая травяная ящерица ( Takydromus sexlineatus).Однако, в отличие от шеи или туловища, размер хвоста не всегда является прямым показателем удлинения позвоночного столба, а зависит от других характеристик. У птиц, например, хвостовые перья являются основным фактором, определяющим размер и разнообразие форм хвоста, часто включая половой диморфизм, приспособленные для облегчения механики полета различных видов и часто участвующие в стратегиях спаривания [81]. Это разнообразие не является следствием различий в размерах их хвостового осевого скелета, который довольно похож у разных видов и состоит из нескольких небольших хвостовых позвонков, за которыми следует пигостиль, скелетный элемент, состоящий из 4-7 сросшихся позвоночных элементов [70] который обеспечивает точку прикрепления хвостовых перьев.Другой источник изменения размера и формы хвоста обеспечивается хвостовым плавником рыб, отростком, простирающимся по оси тела за пределы положения последнего хвостового позвонка, представляющего собой главный источник анатомического разнообразия среди клад рыб [82]. В целом, хвостовые плавники содержат скелет, состоящий из нескольких лучей плавников, костных или хрящевых элементов, по-разному прикрепленных к хвостовым позвонкам. Лучи плавников сильно различаются среди клад по количеству, размеру, форме, жесткости и их связи с осевым скелетом [82,83].В зависимости от их общего строения различают три основных типа хвостовых плавников [84]. У акул гетероцеркальный тип хвостового плавника, т.е. его спинная доля больше брюшной. В этой конфигурации позвоночный столб изогнут дорсально, входя в дорсальную долю [82]. У костистых рыбок, как и у рыбок данио, есть хвостовой плавник гомоцеркального типа, что означает, что у них одинаковые размеры спинных и брюшных плавников. В то время как гомоцеркальный плавник внешне симметричен, его позвоночный скелет все еще изгибается дорсально на каудальном конце, становясь связанным с рядом новых скелетных элементов, в основном вентрального происхождения, которые также различаются у разных видов, которые служат одной из основных поддерживающих структур для лучи плавников [82,85,86].Наконец, двоякодышащие рыбы имеют дифицеркальный тип плавников, у которых лопасти действительно симметричны, поскольку они отходят от спинной и брюшной сторон оси [84]. Даже в пределах этих трех основных типов хвостовых плавников разнообразие среди видов очень велико как по внутренней структуре, так и по подвижности, что позволяет различать профили плавания у разных видов [82,83]. Водные млекопитающие, включая сирен и китообразных, также имеют веерообразная структура на заднем конце хвоста, двуустка, которая, подобно хвостовому плавнику рыб, облегчает движение животного в водной среде.Несмотря на их общую функцию, сосальщики и хвостовые плавники не являются гомологичными структурами. Они по-разному ориентированы относительно тела: хвостовой плавник проходит вертикально вдоль дорсовентральной оси тела, а лапы — это боковые (горизонтальные) продолжения хвоста. Структурно двуустка состоит из плотной волокнистой соединительной ткани [87], лишенной внутренних скелетных или мышечных элементов, которые можно наблюдать в разных типах хвостовых плавников [88,89]. Эти различия могут быть результатом их идентичности в развитии.Происхождение развития, процессы роста и формирования паттерна, участвующие в формировании хвостового плавника, сходны с таковыми у других медальных и боковых плавников, что делает их настоящими непарными придатками рыб [89]. Контроль за формированием двуустки практически неизвестен. У них не видоизмененные задние конечности. Действительно, в то время как взрослые сирены и китообразные не имеют задних конечностей, зачаток конечности фактически индуцируется у эмбриона в более переднем положении по отношению к двуустке, но их развитие останавливается, оставляя на месте только небольшую бедренную кость в мышечной ткани в точке конец брюшной полости [90,91].Пока что несколько описаний развития двуустки не показывают каких-либо элементов, которые, как известно, регулируют развитие конечностей, которые могли бы классифицировать их вместе с парными придатками позвоночных [92], что соответствует отсутствию какой-либо скелетной или мышечной ткани, типичной для этих придатков. Следовательно, хвостовые плавники и сосальщики не могут считаться гомологичными структурами, и, учитывая филогенетические расстояния между сиренами и китообразными, возможно, что их сосальщики также возникли в результате двух независимых событий.Таким образом, хвостовые плавники и сосальщики представляют собой яркий пример конвергентной эволюции, позволяющей приспособить подвижность животного к водной среде. Во время эмбрионального развития осевое удлинение через области туловища и хвоста различается по нескольким параметрам. Основное различие между двумя стадиями осевого расширения — это связь развития шеи и туловища с процессом гаструляции, а удлинение хвоста — с активностью хвостовой почки [93,94,95]. У рыб развитие хвоста также связано с морфогенезом хвостового плавника [82,85], начиная с образования зачатка плавника в брюшной части развивающегося хвоста, который перемещается к заднему концу хвоста по мере его роста и дифференциации [ 96].Зачаток хвостового плавника содержит сигнальные центры, гомологичные тем, которые управляют развитием плавников и конечностей [89,96], которые обеспечивают спинно-вентральную и боковую полярность плавника. На основании локализации таких центров было высказано предположение, что спинной край зачатка хвостового плавника эквивалентен переднему краю парных плавников или зачатка конечности, создавая дорсовентральную полярность, реализация которой может быть ключевым фактором. морфологического разнообразия хвостового плавника [89,96]. У большинства видов позвоночных размер хвоста у взрослого животного напрямую связан со способностью эмбриона продолжать вытягивать ось тела и генерировать сомиты после завершения образования структур туловища.Лягушки представляют собой заметное исключение из этого общего правила: на стадии головастика у них есть длинный и полностью развитый хвост, который имеет много структурных характеристик с хвостом других четвероногих, но приспособлен для облегчения плавания. При переходе к модели взрослого тела во время метаморфоза хвост полностью теряется, в результате чего образуется взрослая лягушка без хвоста [7]. У млекопитающих переход от примитивной полосы к развитию, зависящему от хвостовой почки, запускается Gdf11, действующим на Рецептор Tgfbr1 [57,97].В то время как развитие туловища и хвоста разделяет несколько механизмов контроля роста, в том числе те, которые включают T (Brachyury), гены Cdx или передачу сигналов FGF и WNT [98,99], переключение от примитивной полоски к развитию хвостовой почки связано с изменением в факторах на вершине регуляторной иерархии, что в конечном итоге влияет на окончательный размер хвоста. В противоположность развитию ствола, рост хвостовых зачатков не зависит от Oct4, полагаясь вместо этого на механизмы, управляемые Lin28 / let7 [100,101,102]. В самом деле, стойкая экспрессия Lin28 в хвостовой почке вызывает значительное удлинение хвоста, тогда как его инактивация приводит к его укорочению.Переход от развития туловища к развитию хвоста у рыбок данио и, возможно, также у других анамниотов, по-видимому, зависит от другого набора контролирующих факторов, включая переход к BMP-зависимой сети [103,104]. Генетические эксперименты на мышах показывают, что Hox-гены paralog group 13 может быть центральным компонентом сети, приводящей к концу рост хвоста, по крайней мере, частично за счет подавления активности Lin28 в предшественниках хвостовых зачатков [101,105,106]. Важно отметить, что активности как Lin28 / let7, так и Hox13, по-видимому, влияют на распространение через хвостовую зачатку, но имеют небольшой эффект на удлинение туловища [100,101], указывая на разные внутренние характеристики предшественников туловища и хвоста.Остается определить, действуют ли механизмы, запускающие конец удлинения хвоста, идентифицированные у мышей, и у других видов позвоночных. Исследования на куриных эмбрионах показали, что экспрессия генов Hox13 следует паттернам, напоминающим наблюдаемые у мышей, и что эти гены способны замедлять осевое расширение, когда преждевременно активируются в зоне предшественников куриных эмбрионов [76]. Таким образом, возможно, что механизмы, наблюдаемые у мышей, также действуют, по крайней мере, у цыплят. Однако недавняя работа показала положительное участие Hox13 генов в осевом удлинении эмбрионов рыбок данио [107], что, по-видимому, противоречит данным, полученным на мышах и цыплятах.Причина этого несоответствия не совсем ясна, но, возможно, отражает различия между видами. В частности, в то время как у рыбок данио усечение оси наблюдалось при комбинированном подавлении Hoxd13a и Hoxa13b [107], у мышей двойная инактивация генов Hoxa13 и Hoxd13 усекала аутопод конечностей, но не сообщалось об изменениях в осевом развитии [108,109]. Напротив, инактивация Hoxb13 у мышей приводит к удлинению хвоста [106], указывая на его участие в остановке, а не в продвижении осевого удлинения, также согласуясь с усечением хвоста, наблюдаемым при преждевременной активации Hoxb13 в зоне предшественников мыши [101].Следовательно, возможно, что эмбрионы мышей и рыбок данио используют разные регуляторные сети для удлинения своих хвостовых зачатков, что может отражать сообщаемые различия в механизмах осевого удлинения между мышами и рыбками данио [104]. Действительно, хотя удлинение хвоста у эмбрионов мышей и рыбок данио разделяет некоторые регуляторные особенности, такие как потребность в генах Brachyury или Cdx [110,111,112], генетические данные предполагают, что в том, что касается генов Hox13, развитие хвоста у рыбок данио больше похоже на формирование дистальных конечностей, чем на осевое. расширение у млекопитающих.Интересно, что было описано, что Hoxb13 активируется во время начальных стадий регенерации хвоста аксолотля (Ambystoma mexicanum) [113], что совместимо с ролью в стимулировании удлинения хвоста, а не в качестве стоп-сигнала. Следовательно, возможно, что гены Hox13 влияют на осевое удлинение по-разному у разных клад позвоночных.Кости черепа — Строение — Переломы
Череп представляет собой костную структуру, которая поддерживает лицо и образует защитную полость для мозга.Он состоит из множества костей, которые образованы внутримембранозной оссификацией и соединены швами (фиброзные суставы).
Кости черепа можно разделить на две группы: кости черепа (которые состоят из крыши и основания черепа) и кости лица .
В этой статье мы рассмотрим анатомию костей черепа — их ориентацию, суставы и клиническую значимость.
Череп
Череп (также известный как нейрокраниум) образован верхней частью черепа.Он охватывает и защищает мозг, мозговые оболочки и сосуды головного мозга.
Анатомически череп можно разделить на крышу и основание :
- Крыша черепа — состоит из лобной, затылочной и двух теменных костей. Он также известен как свод черепа.
- Основание черепа — состоит из шести костей: лобной, клиновидной, решетчатой, затылочной, теменной и височной. Эти кости соединяются с 1-м шейным позвонком (атлас), лицевыми костями и нижней челюстью (челюстью).
[старт-клиника]
Клиническая значимость: переломы черепа
Переломы черепа обычно возникают в результате удара тупым предметом или проникающей травмы. При рассмотрении переломов черепа одной областью, имеющей клиническое значение, является птерион — Н-образное соединение между височной, теменной, лобной и клиновидной костями.
Птерион покрывает среднюю менингеальную артерию , и переломы в этой области могут повредить сосуд.Кровь может скапливаться между черепом и твердой мозговой оболочкой, образуя экстрадуральную гематому.
Рис. 2 — Вид черепа сбоку, показывающий путь менингеальных артерий. Обратите внимание на птерион, слабое место черепа, где передняя средняя менингеальная артерия находится под угрозой повреждения. [/ Caption][окончание клинической]
Лицо
Лицевой скелет (также известный как висцерокраниум) поддерживает мягкие ткани лица.
Он состоит из 14 костей, которые сливаются, чтобы вместить орбиты глаз, носовую и ротовую полости, а также пазухи.Лобная кость, обычно кость свода черепа, иногда входит в состав лицевого скелета.
Лицевые кости:
- Скуловая (2) — образует скулы лица и сочленяется с лобной, клиновидной, височной и верхнечелюстной костями.
- Слезная (2) — самые маленькие кости лица. Они составляют часть медиальной стенки глазницы.
- Носовая (2) — две тонкие кости, расположенные у переносицы.
- Нижние носовые раковины (2) — расположены в носовой полости, эти кости увеличивают площадь поверхности носовой полости, тем самым увеличивая количество вдыхаемого воздуха, который может контактировать со стенками полости.
- Небный (2) — расположен в задней части ротовой полости и образует часть твердого неба.
- Верхняя челюсть (2) — включает часть верхней челюсти и твердого неба.
- Вомер — образует заднюю часть носовой перегородки.
[старт-клиника]
Клиническая значимость: переломы лица
Переломы лицевого скелета относительно распространены и чаще всего возникают в результате дорожно-транспортных происшествий, драки на кулаках и падений.
Четыре наиболее распространенных типа переломов лица:
- Перелом носа — наиболее частый перелом лица, вызванный выступающим положением костей носа на переносице.Часто наблюдается значительный отек мягких тканей и связанное с ним носовое кровотечение.
- Перелом верхней челюсти — связанный с высокоэнергетической травмой. Переломы костей верхней челюсти классифицируются по классификации Le Fort в диапазоне от 1 до 3.
- Перелом нижней челюсти — часто двусторонний, возникает непосредственно на стороне травмы и косвенно на противоположной стороне из-за передаваемых сил. Клинические признаки включают боль в месте перелома и смещение зубов (неправильный прикус)
- Перелом скуловой дуги — связанный с травмой боковой части лица.Смещенные переломы могут повредить близлежащий подглазничный нерв, что приведет к ипсилатеральной парестезии чека, носа и губы.
[окончание клинической]
Швы черепа
Швы — это тип фиброзного сустава, который уникален для черепа. Они неподвижны и полностью срастаются примерно к 20 годам.
Эти суставы важны в контексте травмы, поскольку они представляют собой точки потенциальной слабости черепа.Основные швы на черепе взрослого человека:
- Венечный шов — сращивает лобную кость с двумя теменными костями.
- Сагиттальный шов — сращивает обе теменные кости друг с другом.
- Лямбдовидный шов — сращивает затылочную кость с двумя теменными костями.
У новорожденных из-за неполного сращивания швов образуются перепончатые щели между костями, известные как роднички.