Урок 1. обмен веществ – главный признак жизни — Биология — 6 класс
Биология, 6 класс
Урок 1. Обмен веществ – главный признак жизни
Перечень вопросов, рассматриваемых на уроке
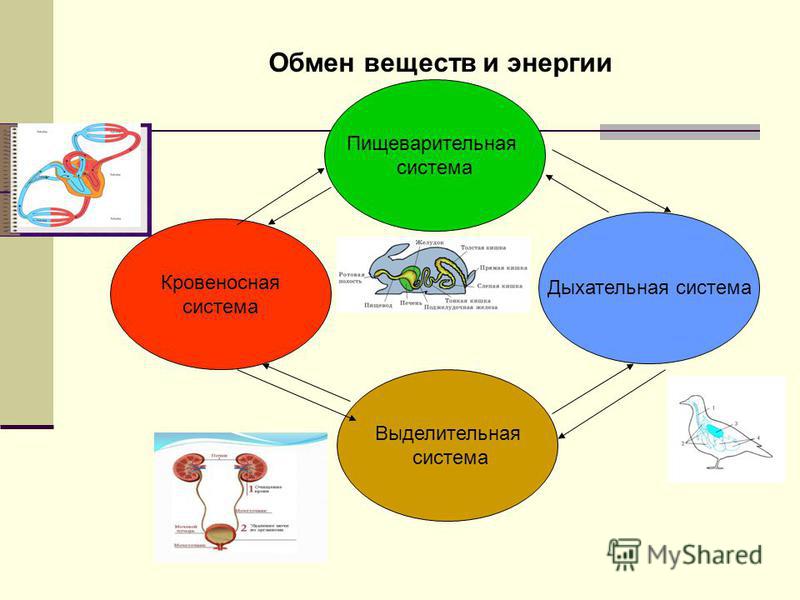
- Обязательным условием жизни является обмен веществ и энергии.
- Обменные процессы, происходящие в организме человека, животных и растений, являются частью общего круговорота веществ в природе.
- Узнаете о процесс создания и образования сложных веществ в тканях и клетках организма.
Тезаурус
Обмен веществ – это процессы поступления нужных организму веществ, их сложных превращений внутри и выведения ненужных веществ в окружающую среду. Эти процессы позволяют организмам расти и размножаться, сохранять свои структуры и отвечать на воздействия окружающей среды.
Гетеротрофы – (др.-греч. heteros – «различный» и trophos – «питание») – организмы, которые питаются готовыми органическими веществами и не способны синтезировать органические вещества из неорганических путём фотосинтеза или хемосинтеза.
Автотрофы — (др.-греч. αὐτός — сам + τροφή — пища) — организмы, синтезирующие органические вещества из неорганических.
*Ассимиляция (Образование веществ) — совокупность процессов синтеза веществ в живом организме.
*Диссимиляция (Разрушение веществ) — это комплекс химических реакций, в которых происходит постепенный распад сложных органических веществ до более простых.
Основная и дополнительная литература по теме урока
- Биология. Линия жизни. 5–6 класс / В. В. Пасечник, С. В. Суматохин, Г. С. Калинова, Г. Г. Швецов, З. Г. Гапонюк – М.: Просвещение, 2018.
- Биология в схемах и таблицах / А.Ю. Ионцева, А.В. Торгалов.
- Введение в биологию: Неживые тела. Организмы : учеб. Для уч – ся 5–6 кл. общеобразоват. учеб. заведений / А.И. Никишов. – М.: Гуманитар. изд. центр ВЛАДОС, 2012.
- Биология. Живой организм. 5 – 6 классы: учебник для общеобразовательных учреждений с приложением на электронном носителе / Л.
 Н. Сухорукова, В. С. Кучменко, И. Я. Колесникова. – М.: Просвещение, 2013.
Н. Сухорукова, В. С. Кучменко, И. Я. Колесникова. – М.: Просвещение, 2013. - Биология. Обо всем живом. 5 класс: учебник / С. Н. Ловягин, А. А. Вахрушев, А. С. Раутиан. – М.: Баласс, 2014.
 Н. Сухорукова, В. С. Кучменко, И. Я. Колесникова. – М.: Просвещение, 2013.
Н. Сухорукова, В. С. Кучменко, И. Я. Колесникова. – М.: Просвещение, 2013.Теоретический материал для самостоятельного изучения
С момента рождения и до смерти в организме происходит обмен веществ и энергии. Для этого из окружающей среды в любой живой организм какие – то вещества должны поступать, а какие – то должны выделяться. Но за счёт каких процессов это возможно? Сегодня на уроке мы будем искать ответ на этот вопрос.
Мы восхищаемся великим разнообразием живых существ. Все они отличаются друг от друга цветом, формой, величиной, строением. Но объединяет всех их одно – жизнь.
Проникнуть в тайны жизни человек пытался давно. Было доказано, что различие между живой и неживой природой заключается в особом строении живого существа и в специфических химических процессах, постоянно происходящих между организмом и внешней средой. Совокупность этих процессов и представляет собой основу жизни.
Примеры и разбор решения заданий тренировочного модуля
Задание 1. Распределите живые организмы по типам питания.
| Гетеротрофы | Автотрофы |
Варианты ответов:
Кактус.
Подсолнечник.
Шиповник.
Берёза.
Человек.
Жук.
Пшеница.
Собака.
Осьминог.
Чайка.
Правильный вариант ответа:
| Гетеротрофы | Автотрофы |
Человек Жук Собака Осьминог Чайка | Кактус Подсолнечник Шиповник Берёза Пшеница |
Задание 2. Распределите элементы по группам. Что такое обмен веществ? Каково его значение для организма?
Значение обмена веществ для организма | Обмен веществ – это процесс |
Варианты ответов:
Выведение вещества из организма.
Без обмена веществ невозможно поступление веществ в организм.
Потребления, превращения, использования и выделения вещества.
Без обмена веществ невозможен рост и обновление клеток организма.
Поступления вещества в организм.
Правильный вариант:
Значение обмена веществ для организма | Обмен веществ – это процесс |
|
|
Обмен веществ и превращение энергии – свойства живых организмов. Энергетический обмен и пластический обмен, их взаимосвязь. Стадии энергетического обмена. Брожение и дыхание / Справочник :: Бингоскул
Обмен веществ и превращение энергии – свойства живых организмов
Обмен веществ является комплексом различных химических преобразований, способствующих сохранению и самовоспроизведению биоструктур.
Он заключается в поступлении веществ в организм во время питания и дыхания, метаболизме внутри клетки или обмене веществ, вдобавок, в высвобождении конечных продуктов метаболизма.
Метаболизм неотрывно соединён с процессами преобразований определённых видов энергии в другие. К примеру, в начале процесса фотосинтеза световая энергия скапливается в виде энергии химических связей сложных органических молекул, в процессе же дыхания она освобождается и применяется для синтезирования новых молекул, механические и осмотические работы, рассеянные в виде тепла и т. д.
Поток химических превращений в живых организмах снабжается биологическими катализаторами белковой специфики — ферментами или энзимами. Наряду с остальными катализаторами, энзимы ускоряют течение химических реакций в клетке до нескольких сотен тысяч раз, при этом они не меняют природу или свойства конечных продуктов клетки. Ферменты представляют собой простые или сложные белковые молекулы, которые, помимо части, состоящей из белка, включают небелковый кофактор, по – другому называемый коферментом. Ферментами являются, например: амилаза слюны, которая расщепляет гликаны при длительном жевании и пепсин, который обеспечивает переваривание белков в желудочно-кишечном тракте.
Ферментами являются, например: амилаза слюны, которая расщепляет гликаны при длительном жевании и пепсин, который обеспечивает переваривание белков в желудочно-кишечном тракте.
Ферменты различаются с небелковыми катализаторами тем, что имеют высокую специфичность действия, в значительной степени увеличенную скорости реакции, а также возможностью регулирования действия путем смены условий реакции или взаимодействия различных веществ с ними. Кроме того, условия, при которых протекает ферментативный катализ, значительно различаются с теми, при которых происходит неферментативный катализ: оптимальная температура для того, чтобы ферменты могли функционировать в организме человека, составляет 37 ° С, а также необходимо, чтобы давление являлось близким к атмосферному, в то время как кислотность среды может значительно варьироваться. Например, для амилазы необходима щелочная среда, для пепсина же наоборот — кислая.
Механизм действия ферментов заключается в том, чтобы снизить энергию активации веществ (субстратов), которые вступают в реакцию вследствие образования промежуточных фермент-субстратных комплексов.
Энергетический и пластический обмен, их взаимосвязь
Метаболизм процессуально слагается из двух частей, происходящих в клетке в одно и то же время: пластического и энергетического обмена.
Пластический метаболизм (анаболизм, ассимиляция) является совокупностью реакций синтеза, сопровождающихся расходом энергии аденозинтрифосфата. Пластический обмен особенно важен тем, что в результате него синтезируются органические вещества, играющие важную роль в жизнедеятельности клетки. Реакциями данного обмена являются, например, процесс фотосинтеза, биологический синтез белковых молекул и репликация молекул ДНК (самодублирование).
Энергетический обмен (катаболизм, диссимиляция) являет собой сочетание реакций разложения сложных веществ на более простые. Результатом данного обмена является накапливание энергии в форме АТФ. Важнейшими процессами энергетического обмена являются дыхание и брожение.
Пластический и энергетический обмены прочно коррелируют между собой, в связи с тем, что синтез органических веществ происходит в процессе пластического обмена, а для этого нужна именно энергия АТФ; в процессе обмена энергии органические вещества разлагаются, и высвобождается АТФ, а затем используется для синтеза.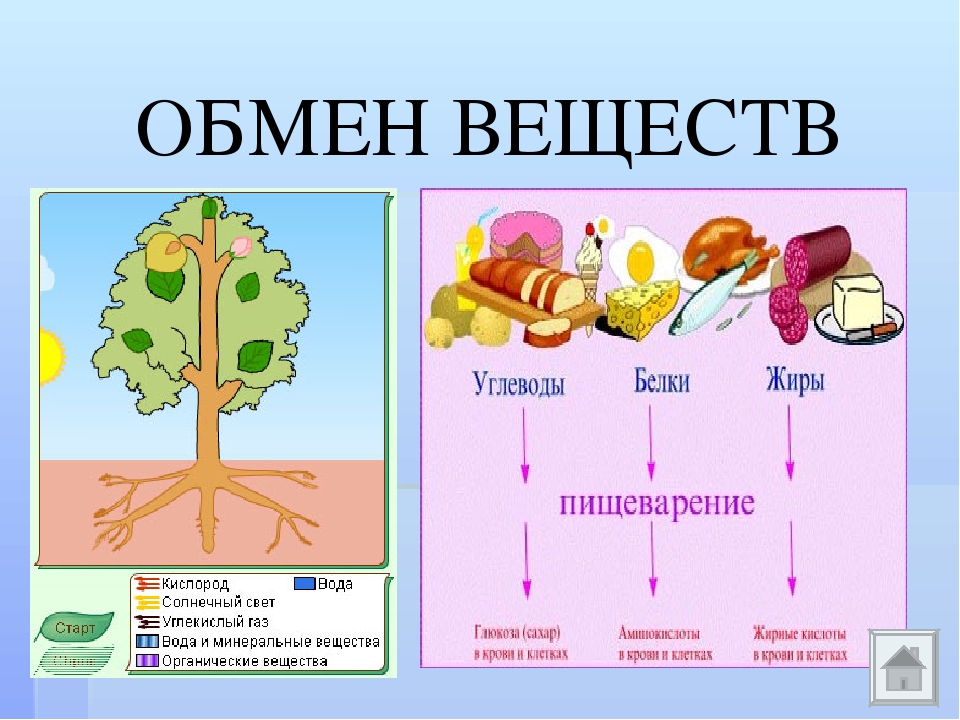
Получение энергии организмами осуществляется в процессе питания, затем высвобождают ее и переводят в форму, доступную главным образом в процессе дыхания. По способу питания все организмы подразделяются на автотрофные и гетеротрофные. Автотрофы способны к самостоятельному синтезу органических веществ из неорганических, а гетеротрофные организмы поглощают уже готовые органические вещества.
Ассимиляция — биосинтез макромолекул, свойственных клеткам организма. Растения и многие бактерии могут создавать молекулы глюкозы из углекислого газа и воды. На этот процесс расходуется и запасается энергия. Животным необходимы готовые молекулы белков, жиров и углеводов (БЖУ). Это важнейший строительный и энергетический материал для клеток.
Ассимиляция — это совокупность процессов создания структур организма с накоплением энергии.
Значение метаболизма:
- Поступление из внешней среды веществ, необходимых для организма;
- Превращение питательных веществ в соединения, которые могут использоваться клетками и тканями;
- Синтез структурных элементов клеток, ферментов и т. д., замена устаревшим новыми;
- Синтез более сложных соединений из более простых;
- Отложение запасов.
 д., замена устаревшим новыми;
д., замена устаревшим новыми;Чтобы организм мог усвоить вещества из пищи, они должны быть сначала разобраны на «кирпичики» или мономеры. Из них в организме «собираются» собственные макромолекулы.
Диссимиляция — распад веществ, противоположный ассимиляции (биосинтезу). Белки гидролизуются до аминокислот. При распаде жиров выделяются жирные кислоты и глицерин. Сложные углеводы разлагаются на простые сахара.
Ассимиляция и диссимиляция происходят согласованно. Распад и окисление веществ с выделением энергии возможны лишь тогда, когда есть субстрат — макромолекулы. Они разлагаются на мономеры, которые участвуют в биосинтезе. Выделяющаяся при диссимиляции энергия затрачивается на образование свойственных организму веществ.
Стадии энергетического обмена
Несмотря на сложность реакций обмена энергии, он разделяется на три фазы:
- подготовительная,
- анаэробная (без кислорода),
- аэробная (кислород).

На подготовительном этапе происходит разложение молекул гликанов, липидов, белков, нуклеиновых кислот на более простые, к примеру, на глюкозу, глицерин и жирные кислоты, аминокислоты, нуклеотиды. Эта фаза может осуществляться непосредственно в клетках или в кишечнике, откуда эти вещества переносятся кровотоком.
В анаэробной фазе энергетического катаболизма в дальнейшем происходит расщепление мономеров органических соединений до более простых промежуточных соединений, к примеру, пировиноградной кислоты или пирувата. Он не нуждается в присутствии кислорода, и для организмов, живущих в болотном иле, это единственный способ получить энергию. Анаэробная фаза энергетического обмена проходит в цитоплазме.
Некоторые вещества подвергаются бескислородному расщеплению, при этом глюкоза, чаще всего, остается основным субстратом реакций. Процесс его свободного от кислорода распада принято называть гликолизом. Вследствие гликолиза, молекула глюкозы теряет четыре атома водорода, то есть она окисляется, и образуются две молекулы пировиноградной кислоты, две молекулы АТФ и две молекулы переносчика водорода, восстановленного НАДH + H +:
С6Н12О6+2Н3РО4+2АДФ+2НАД→2С3Н4О3+2АТФ+2НАДН+Н++2Н2О.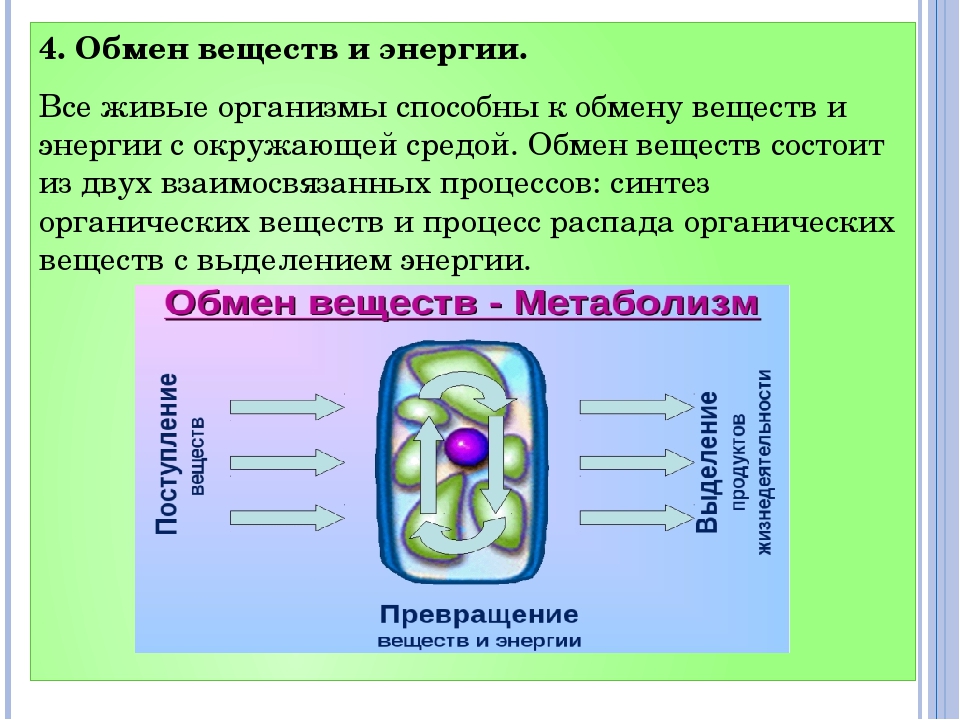
Образование АТФ из АДФ осуществляется за счет прямого переноса фосфат-аниона из предварительно фосфорилированного сахара и называется субстратным фосфорилированием.
Аэробная фаза энергетического катаболизма может происходить только в присутствии кислорода, тогда как промежуточные продукты, образующиеся при бескислородном разложении, окисляются до конечных продуктов (углекислого газа и воды), и большая часть энергии, хранящейся в химических связях органических соединений, высвобождается. В молекулу АТФ входит 36 макроэргических связей. Эта стадия имеет такое название, как тканевое дыхание. Когда кислород отсутствует, происходит преобразование промежуточных продуктов обмена веществ в определённые органические вещества, данный процесс принято называть ферментацией или брожением.
Брожение и дыхание
Брожение и дыхание это две различные формы диссимиляции — разложения веществ в организме для получения энергии.
Брожение
Примеры процессов брожения известны из повседневной жизни, производственной деятельности.
- Спиртовое брожение заключается в метаболическом превращении углеводов микроорганизмами, преимущественно дрожжами. В результате образуется этиловый спирт, АТФ и вода, выделяется углекислый газ. Энергию микроорганизмы используют для жизнедеятельности, деления клеток. Спиртовое брожение используется в производстве алкогольных напитков. Пекарские дрожжи в хлебопечении тоже перерабатывают углеводы на этанол и углекислый газ, разрыхляющий тесто.
- Молочнокислое брожение завершается образованием молекул молочной кислоты, АТФ, водорода и воды. Так скисает молоко, получается пахта, йогурт, сметана, творог. (Рисунок 1). Этот же тип брожения происходит при квашении капусты. Молочнокислые бактерии уменьшают рН субстрата, создают кислую среду. Они не нуждаются в кислороде, но выживают и в кислородной среде.
- Уксуснокислое брожение приводит к изменениям сока, вина. Сначала, в результате спиртового брожения, вырабатывается этанол. Затем, уксуснокислые бактерии перерабатывают спирт на органические кислоты, в основном яблочную, лимонную, молочную. Так получают натуральный уксус из плодово-ягодного сырья.
 Так получают натуральный уксус из плодово-ягодного сырья.
Так получают натуральный уксус из плодово-ягодного сырья.Во всех случаях брожения микроорганизмы изменяют углеводы и производят макроэнергетическое вещество — АТФ. Для этого процесса не требуется кислород, что является важнейшим отличием от дыхания. Общий признак — химическая энергия связей в молекуле глюкозы преобразуется в энергию в форме АТФ, которая используется для жизненных процессов.
Брожение — древнейший и не самый совершенный способ выработки энергии. Из одной молекулы глюкозы образуется 2 молекулы АТФ. Кислородный процесс более эффективен в плане получения энергии.
Организмы, которым необходим кислород для дыхания, являются аэробами (в переводе с греческого «аэр» — воздух). Внешняя сторона процесса заключается в поглощении кислорода из воздуха и выделении диоксида углерода.
Молекулы О2 попадают в организм насекомых через трахеи. Для рыб характерно жаберное дыхание, для млекопитающих — легочное.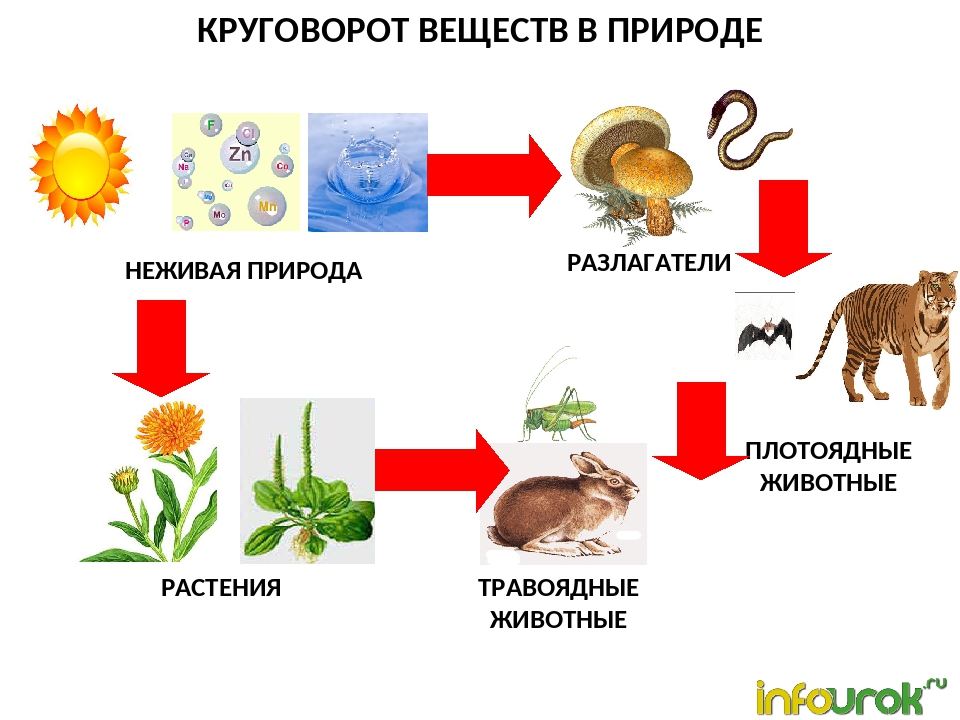 Переносят кислород к органам и транспортируют диоксид углерода красные кровяные клетки, содержащие гемоглобин.
Переносят кислород к органам и транспортируют диоксид углерода красные кровяные клетки, содержащие гемоглобин.
При отсутствии кислорода начинает происходить ферментация. Ферментация является эволюционно более ранним способом генерирования энергии, чем дыхание, но она менее энергетически выгодна, потому что ферментация производит органическое вещество, которое все еще богато энергией. Различают несколько основных видов брожения: уксусно – кислое, спиртовое, маслянокислое, молочнокислое, метановое и др.
Стало быть, в скелетных мышцах в отсутствие кислорода во время ферментации пировиноградная кислота восстанавливается до молочной кислоты, тогда как ранее образованные восстановительные эквиваленты расходуются, и остаются только две молекулы АТФ:
2С3Н4О3 + 2НАДН+Н+ → 2С3Н6О3 + 2НАД.
При ферментации с дрожжами пировиноградная кислота в присутствии кислорода преобразуется в этиловый спирт и окись углерода (IV):
С3Н4О3 + НАДН+Н+ → С2Н5ОН + СО2↑ + НАД+.
Во время ферментации с использованием микроорганизмов пируват также может образовывать уксусную, масляную, муравьиную кислоты и так далее.
Энергия АТФ, которая образуется вследствие энергетического обмена, используется клеткой на различные виды работ:
- Химическая работа включает в себя биосинтез белков, липидов, углеводов, нуклеиновых кислот и других важных соединений.
- Осмотическая работа включает процессы поглощения и удаления веществ из клетки, находящиеся во внеклеточном пространстве в более высоких концентрациях, чем в самой клетке.
- Электрическая работа неразрывно связана с осмотической, ведь именно из – за перемещения заряженных частиц через мембраны формируется заряд мембраны и приобретаются свойства возбудимости и проводимости.
- Механическая работа связана с передвижением веществ и структур во внутриклеточном пространстве и непосредственно клетки в целом.
- К регуляторной работе относят все процессы, которые направлены на координировании процессуальных действий в клетке.
Дыхание
Кислородное дыхание производится в митохондриях, где пировиноградная кислота вначале теряет один атом углерода, что сопровождается синтезом одного восстанавливающего эквивалента молекул НАДН+Н+ и ацетилкофермента A (ацетил-КоА):
С3Н4О3 + НАД+Н ~ КоА → СН3СО ~ КоА + НАДН+Н+ + СО2↑.
Ацетил-КоА в митохондриальном матриксе участвует в цепочке химических превращений, которые в совокупности называются циклом Кребса (цикл трикарбоновых кислот, цикл лимонной кислоты). Во время этих превращений образуются две молекулы АТФ, ацетил-КоА полностью окисляется до диоксида углерода, а его ионы водорода и электроны присоединяются к водородным векторам НАДН+Н+ и НАДh3. Носители переносят протоны и электроны водорода во внутренние митохондриальные мембраны, которые образуют гребни. При помощи белков-носителей протоны водорода вводятся в межмембранное пространство, а электроны переносятся через, так называемую, дыхательную цепь энзимов, которые расположены во внутренней митохондриальной мембране, и разряжаются в атомы кислорода:
O2 + 2e− → O2−.
Важно то, что в дыхательной цепи имеются белки, содержащие железо и серу.
Протоны водорода переносятся из межмембранного пространства в митохондриальный матрикс благодаря специальным ферментам, АТФ-синтетаз, а энергия, выделенная в результате этого процесса, используется для синтеза 34 молекул АТФ из каждой молекулы глюкозы. Этот процесс называется окислительным фосфорилированием. В митохондриальной матрице протоны водорода, прореагировавшие с радикалами кислорода с образованием воды:
4H+ + O2−→ 2H2O.
Набор кислородных дыхательных реакций можно выразить таким уравнением:
2С3Н4О3 + 6О2 + 36Н3РО4 + 36АДФ → 6СО2↑ + 38Н2О + 36АТФ.
Общее уравнение дыхания выглядит следующим образом:
С6Н12О6 + 6О2 + 38Н3РО4 + 38АДФ → 6СО2↑ + 40Н2О + 38АТФ.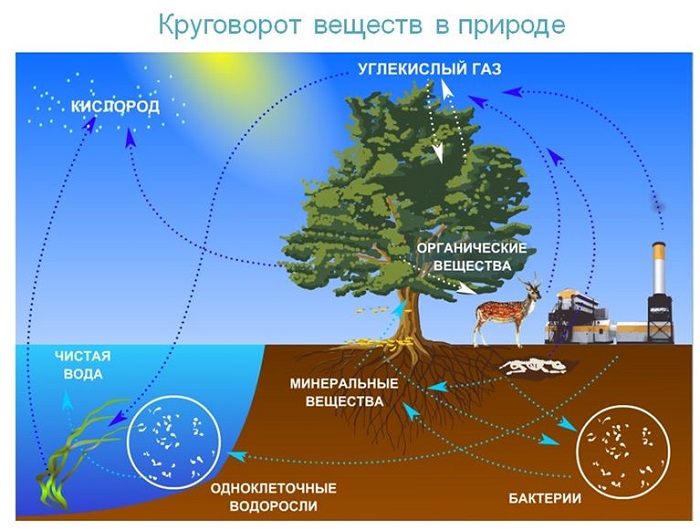
Таким образом, клеточное дыхание в организме человека происходит поэтапно. Гликолиз сопровождается образованием 8 молекул АТФ (2 из них расходуются). Окислительное декарбоксилирование «дает» 6 АТФ, цикл Кребса — 24 АТФ. Итого, разложение молекулы глюкозы приводит к созданию 38 молекул АТФ. Аэробное дыхание — более совершенный способ получения и накопления энергии.
Смотри также:
1. Как происходит обмен веществ и энергией в живой клетке? Чем он отличается от обменных процессов в неживой природе?
Министерство образования
Российской Федерации
ТОМСКИЙ ГОСУДАСТВЕННЫЙ УНИВЕРСИТЕТ
СИСТЕМ УПРАВЛЕНИЕ И РАДИОЭЛЕКТРОНИКИ (ТУСУР)
Кафедра автоматизированных систем управления (АСУ)
ОТЧЕТ
По контрольной работе №2
по дисциплине «Концепции современного естествознания»
Вариант 0614
студента xxxxxxxxxxxxxxxxxxxx
из
г.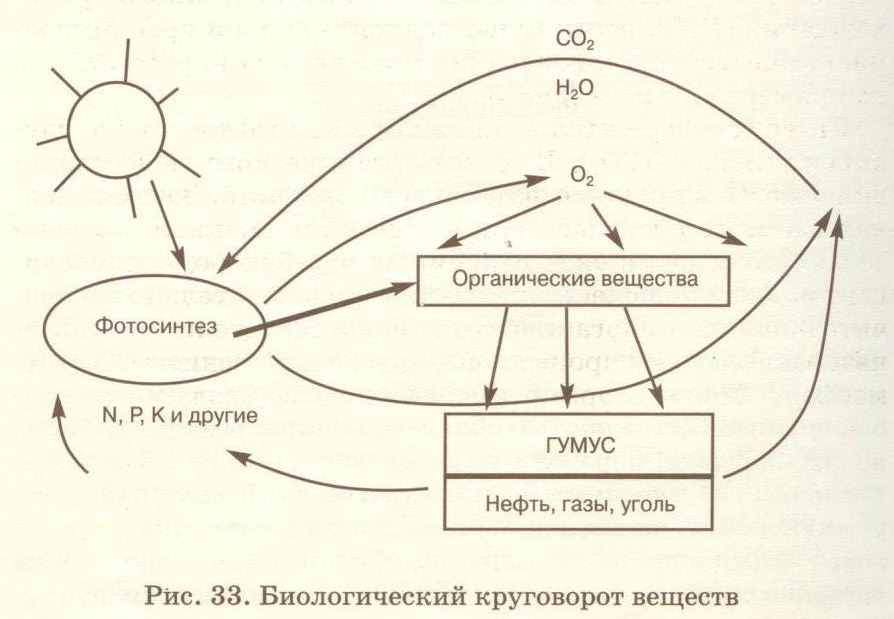 NNNNNNNNNNNNNNNNN
NNNNNNNNNNNNNNNNN
2008
Основное жизненное свойство клетки – обмен веществ. Из межклеточного вещества в клетки постоянно поступают питательные вещества и кислород, выделают продукты распада. Вещества поступившие в клетку, постоянно участвуют в процессах биосинтеза. Биосинтез –это образование специфических белков, жиров, углеводов и их соединений из более простых веществ.
В процессе биосинтеза образуются вещества, свойственным клеткам организма. Например, в клетках мышц синтезируются белки, обеспечивающие их сокращение. Одновременно с биосинтезом в клетках происходит распад органических соединений. В результате распада образуются вещества более простого строения.
Большая часть
реакций распада идет с участием кислорода
и освобождением энергии. Эта энергия
расходуется на жизненные процессы,
протекающие в клетке. Процессы биосинтеза
и распада составляют обмен веществ,
который сопровождается превращениями
энергии клеткам свойственны рост и
размножение.
Клетки тела человека размножаются делением пополам. Каждая из образовавшихся дочерних клеток растет и достигает размеров материнской. Новые клетки выполняют функции материнской клетки. Продолжительность жизни клеток различна: от нескольких часов до десятков лет. Живые клетки способны реагировать на физические и химические изменения окружающей среды. Это свойство клетки называется возбудимостью. При этом из состояния покоя клетка переходит в рабочее состояние – возбуждение.
При возбуждении в клетках меняется скорость биосинтеза и распада веществ, потребление кислорода, температура. В возбужденном состоянии разные клетки выполняют свойственные им функции. Железистые клетки образуют и выделяют вещества, мышечные – сокращаются, в нервных клетках возникает слабый электрический сигнал – нервный импульс, который может распространяться по клеточным мембранам.
Важно отметить,
что обмен веществ может иметь место и
между телами неживой природы. Однако
обмен веществ как свойство живого
качественно отличается от обменных
процессов в неживых телах. Для того
чтобы показать эти отличия, рассмотрим
ряд примеров.
Однако
обмен веществ как свойство живого
качественно отличается от обменных
процессов в неживых телах. Для того
чтобы показать эти отличия, рассмотрим
ряд примеров.
Горящий кусок угля находится в состоянии обмена с окружающей природой: происходит включение кислорода в химическую реакцию и выделение углекислого газа. Образование ржавчины на поверхности железного предмета является следствие обмена со средой. Но в результате этих процессов неживые тела перестают быть тем, чем они были. Для тел живой природы, наоборот, обмен с окружающей средой является условием их существования. В живых организмах обмен веществ приводит к восстановлению разрушенных компонентов, замене их новыми, подобными им, т. е. к самообновлению и самовоспроизведению, построению тела живого организма за счет усвоения веществ из окружающей среды.
Из
сказанного следует, что организмы
существуют как открытые системы. Через
каждый организм идут непрерывные потоки
вещества и энергии.
2.Поясните процессы в расплавах и растворах. Почему при растворении обычно температура понижается? Каковы особенности растворения в воде? Какую роль играют гидрофильные и гидрофобные процессы в живых организмах?
В воде растворяется гораздо больше веществ, чем в любой другой жидкости. Молекулы растворяемого вещества отделяются друг от друга и смешиваются с молекулами растворителя. Из-за своей полярной природы вода растворяет ионные вещества и другие полярные соединения. В то же время соединения не имеющие заряда, в воде не растворяются ,между ними образуется поверхность раздела. Это обстоятельство очень важно для процессов внутри живых структур.
Гидрофильные свойства полярных молекул важны в обеспечении стабильности мембран, белковых молекул, нуклеиновых кислот и других субклеточных структур. Неполярные вещества, такие как липиды, с водой не смешиваются и разделяют
водные
растворы на отдельные компоненты, как
их разделяют мембраны. Поэтому неполярные
части молекул водой отталкиваются и в
присутствии воды притягиваются друг к
другу, или обладают гидрофобным эффектом.
Пример сливание капелек масла в большую
каплю и не растворение ее в воде.
Поэтому неполярные
части молекул водой отталкиваются и в
присутствии воды притягиваются друг к
другу, или обладают гидрофобным эффектом.
Пример сливание капелек масла в большую
каплю и не растворение ее в воде.
Урок биологии «Обмен веществ» 5 класс
Тема: «Обмен веществ и превращение энергии»
Цели урока:
Методы обучения: проблемный, эвристический.
Оборудование: компьютер, мультимедийный проектор, компьютерная презентация «Обмен веществ и энергии», «Рабочая тетрадь. Биология: 6 класс».
ХОД УРОКА
I. Организационный момент
1. Проверка готовности класса к уроку.
II. Этап всесторонней проверки знаний.
Презентация.
Прежде чем приступить к изучению нового материала, давайте, попробуем ответить на вопрос «В чем отличие живых организмов от неживой среды?»
Признаки живого организма
Питание
Дыхание
Рост
Развитие
?
III.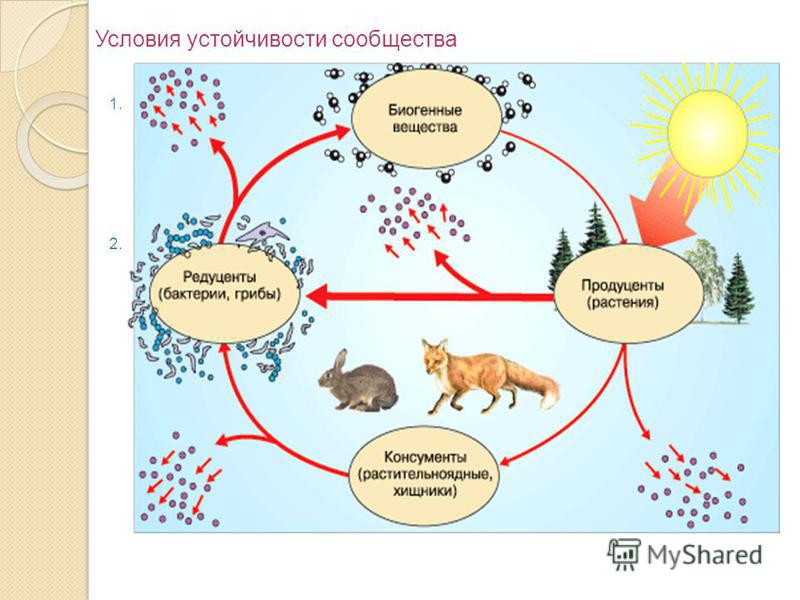 Актуализация знаний
Актуализация знаний
Чтобы узнать какое свойство мы будем изучать следующим, давайте посмотрим на слайд. Как вы понимаете фразу «Организм – открытая, саморегулирующаяся система»?
Для каких веществ организм открыт? Как вещества поступают в организм? Как называются эти процессы?
Что происходит с избытком веществ? Каким образом вещества выводятся из организма?
Этот процесс называется обменом веществ. Сообщение темы урока
Давайте сформулируем определение? Что такое обмен веществ?
Давайте сформулируем определение? Что такое обмен веществ?
Известно, что обмен веществ можно рассматривать с разных сторон: обмен веществ различается у растений и у животных, а также обмен веществ — это химический процесс.
Для того, чтобы узнать как можно больше о процессе обмена веществ мы разделимся на группы:
Химики
Ботаники
Зоологи
Работа в группах:
1 группа: «Химики»
Задача: Изучить химическую природу обмена веществ.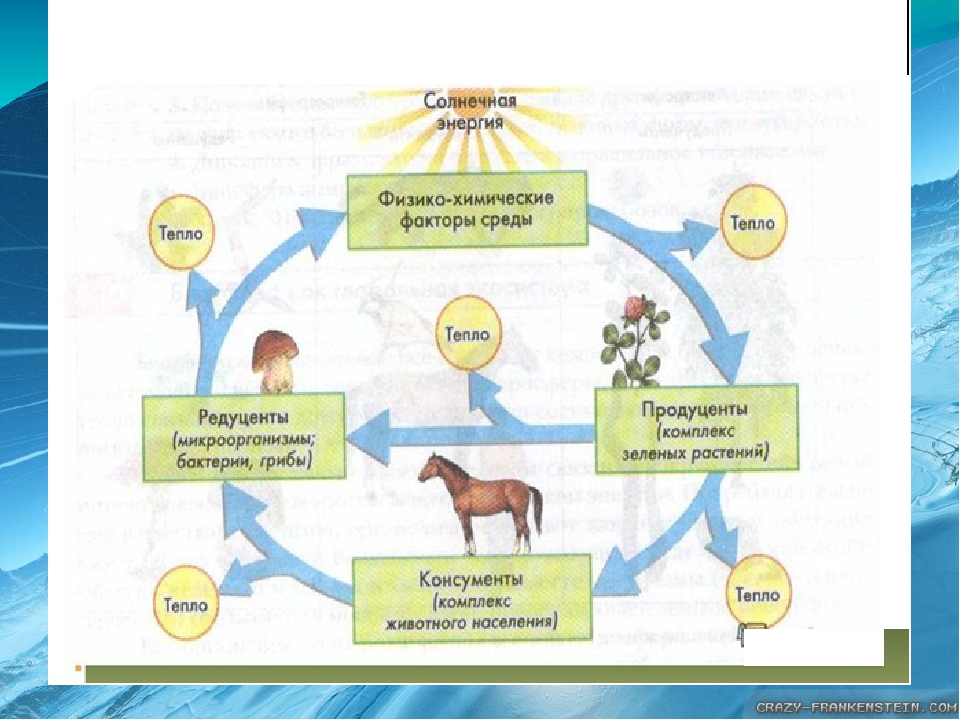
Ход работы:
Прочитайте текст, рассмотрите картинки и составьте план ответа по вопросам:
Как по-другому называется обмен веществ?
На какие стадии делится обмен веществ?
Что происходит на стадии катаболизма?
Что происходит на стадии анаболизма?
Что такое ферменты? Зачем они нужны в химических реакциях?
Заполните схему:
2 группа «Ботаники»
Задача: Выявить особенности обмена веществ у растений.
Ход работы:
Прочитайте текст, рассмотрите картинки и составьте план ответа по вопросам:
О каком процессе рассказывается в вашем тексте?
Как выглядит уравнение фотосинтеза?
Какие вещества поступают в растение?
Какие вещества образуются на свету?
Как используются эти вещества?
Что дальше происходит с этими веществами?
Как называется этот процесс?
Благодаря чему происходит этот процесс?
Как используется полученная энергия?
Заполните таблицу
3 группа « Зоологи»
Задача: Выявить особенности обмена веществ у животных.
Ход работы:
Прочитайте текст, рассмотрите картинки и составьте план ответа по вопросам:
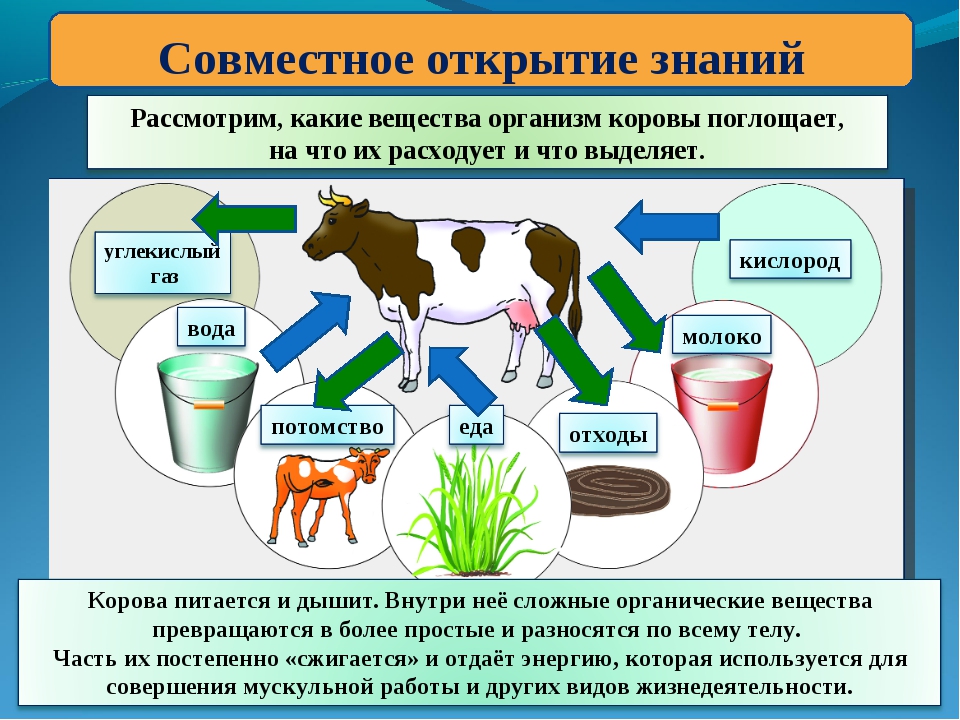
Какие вещества поступают в организм животных?
Как называются процессы, при которых поступают вещества?
Что с ними происходит с веществами внутри организма?
Какие вещества выделяются в результате?
Заполните таблицу
Индивидуальные задания:
Прочитайте текст, рассмотрите картинки и составьте план ответа по вопросам:
Перечислите факторы, влияющие на скорость обмена веществ? Составьте схему.
Какая зависимость обмена веществ от температуры?
Какие бывают группы животных относительно температуры тела? Чем отличается их обмен веществ?
Обмен информацией:
Выступление 1 представителя от каждой группы, заполнение таблицы, схем.
IV. Обобщение:
Обобщение:
Значение обмена веществ:
Обеспечение организма энергией и питательными веществами
Рост и развитие
Круговорот веществ в природе
V. Закрепление нового материала:
Тест
1. Обмен веществ происходит у всех живых организмов.
2.Обмен веществ складывается из двух противоположных процессов.
3.В обмене веществ у растений принимают участие только листья.
4.В растения из окружающей среды поступают кислород, углекислый газ и вода.
5.В обмене веществ у животных принимают участие только органы дыхания и кровеносной системы.
6.В результате фотосинтеза образуются органические вещества и кислород.
7.Теплокровные животные имеют постоянную температуру тела.
8.Змеи и лягушки – это теплокровные животные.
9.Медведь – холоднокровное животное, так как впадает в зимнюю спячку.
10. Зимой у птиц обмен веществ протекает медленно, потому что температура окружающей среды низкая.
11. Активный обмен веществ у птиц и млекопитающих способствовал их широкому распространению на Земле.
12. Обмен веществ и энергии является основным признаком живого организма.
Ответы: 1, 2, 4, 6, 7, 11, 12 — да
V. Подведение итогов урока
Рефлексия:
– Узнали ли вы, что-то новое на уроке?
– Какие знания вы применили на уроке для понимания новой темы?
– Что произвело на вас наибольшее впечатление?
– Хочется ли узнать что-нибудь еще по этой теме?
Нарисуйте смайл.
VI. Домашнее задание
П. 44 – прочитать, выучить определения
Задание по выбору:
Кроссворд «Обмен веществ», 10 вопросов.
Сообщение «Питание и обмен веществ».
Лошади
Молодняк
Жеребенку для оптимального развития необходимы высококачественные протеины, которые он получает вначале с молоком матери, а впоследствии они должны поступать в организм жеребенка в специально составленном стартерном корме с незаменимыми аминокислотами. Карнитин поддерживает обмен веществ в молодом организме и, играя особую роль в углеводном и жировом обменах, оказывает, кроме того, положительное воздействие на развитие скелета.
Карнитин поддерживает обмен веществ в молодом организме и, играя особую роль в углеводном и жировом обменах, оказывает, кроме того, положительное воздействие на развитие скелета.
Пожилые лошади
Стареющие лошади зачастую не в состоянии удерживать свой вес. Это связано, как правило, с изношенностью зубов и проистекающими отсюда трудностями с пережевыванием корма в полном объеме, а также со снижением усвояемости питательных веществ. Поэтому для этой категории лошадей требуется калорийный корм с хорошо усвояемыми протеинами или, иначе говоря, целенаправленная добавка в их корм необходимых аминокислот во избежание ненужного обременения обмена веществ животного. Как правило, для этой группы лошадей рекомендуется стабилизация пищеварительной системы с помощью добавок специальных растительных экстрактов и эфирных масел (Spicemaster GH).
В преклонном возрасте у лошадей нередко возникают нарушения в ходе линьки и проблемы с повышенной чувствительностью кожи. В таких случаях помогают добавки цинк-метионина и биотина. Применение растительных полифенолов помогает при наличии артроза у животного. Если у лошади возникают проблемы с органами дыхания, рекомендуется применение добавок со специальными эфирными маслами (Lovit Breeze) с тем, чтобы облегчить дыхание и снизить кашель.
Применение растительных полифенолов помогает при наличии артроза у животного. Если у лошади возникают проблемы с органами дыхания, рекомендуется применение добавок со специальными эфирными маслами (Lovit Breeze) с тем, чтобы облегчить дыхание и снизить кашель.
Лошади с нарушенным обменом веществ
К корму лошадей с нарушенным обменом веществ, с такими заболеваниями как метаболический синдром лошадей (МСЛ) или синдром Кушинга, предъявляются особенно высокие требования: он должен быть сбалансирован таким образом, чтобы привести нарушенный обмен веществ снова в норму, сдерживать связанные с ним воспалительные процессы и, кроме того, в корме должна быть учтена резистентность таких лошадей к инсулину. Здесь чрезвычайно важное значение имеет применение карнитина и растительных полифенолов.
Главврач назвал основным диагнозом Навального нарушение обмена веществ
Основной диагноз госпитализированного оппозиционера Алексея Навального — нарушение обмена веществ, заявил главный врач омской больницы скорой помощи №1 Александр Мураховский.
«На сегодняшний день у нас есть рабочие диагнозы. Основной из них, к которому мы склоняемся,— нарушение углеводного баланса, то есть, нарушение обмена веществ»,— сказал господин Мураховский. Он допустил, что это может быть вызвано резким понижением уровня сахара в крови.
Нарушение обмена веществ, резкое падение уровня сахара в крови. Заявление Александра Мураховского, главврача больницы, где лежит Алексей Навальный. pic.twitter.com/MxQEoXbvJE
— The Insider (@the_ins_ru) August 21, 2020
Врачи омской больницы впервые огласили диагноз господина Навального, до этого они ссылались на закон о защите персональных данных. Заместитель главного врача омской больницы Анатолий Калиниченко ранее утверждал, что диагноз устно сообщили жене и брату господина Навального. Юлии и Олегу Навальным не сообщали диагноз, а то, что им рассказали — это «набор симптомов, который может быть интерпретирован по-разному», заявила пресс-секретарь политика Кира Ярмыш.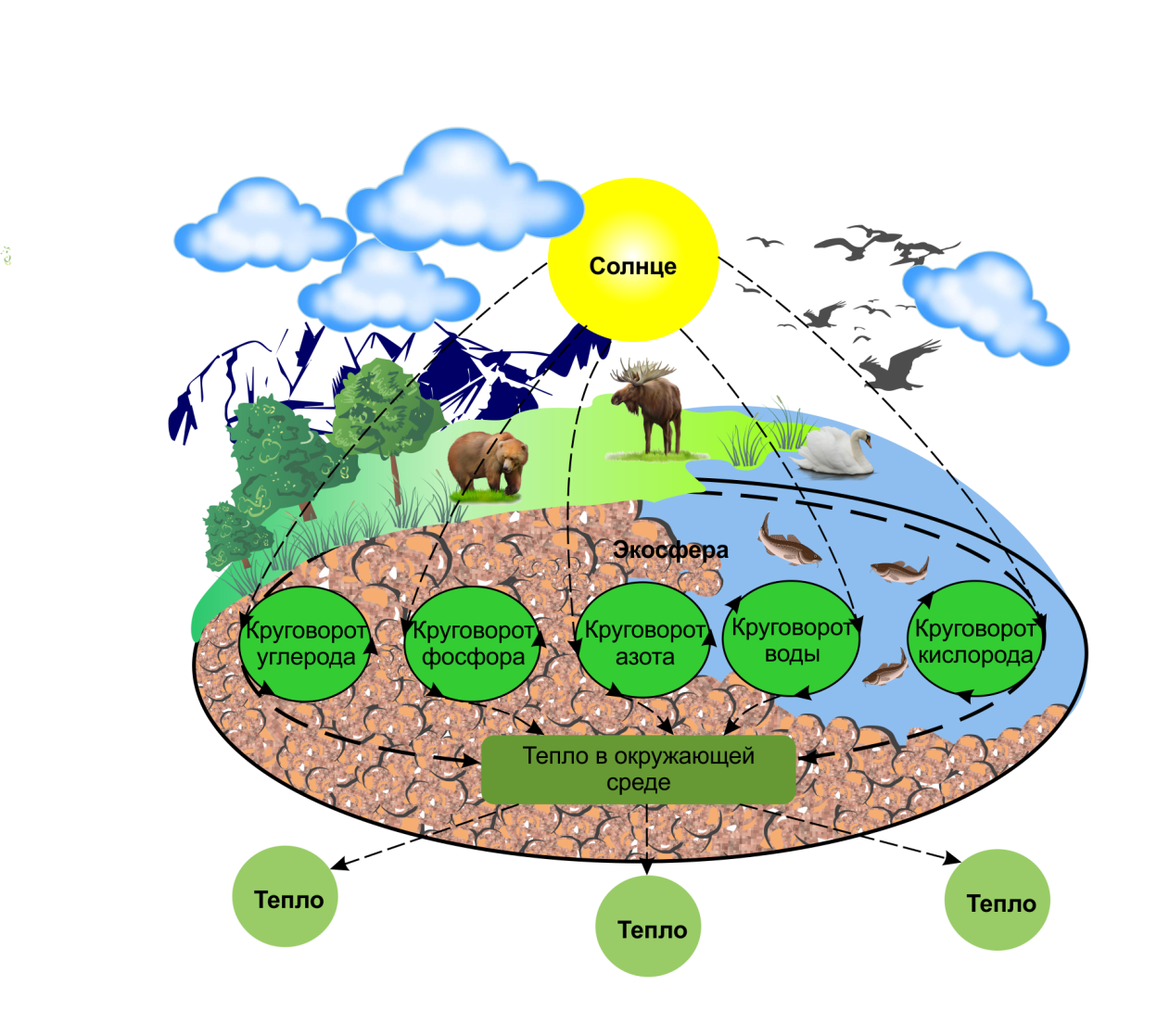
Оксибутираты и барбитураты в крови господина Навального не найдены, сказал главврач. Ранее его заместитель Анатолий Калиниченко говорил, что в крови и моче политика яды или их следы не обнаружены. Вещество, найденное на коже и одежде господина Навального — это материал пластикового стаканчика, «обычная промышленная химия», отметил Мураховский. В крови пациента, по его словам, этого компонента нет.
Ранее сегодня директор Фонда борьбы с коррупцией (ФБК) Иван Жданов заявлял, что транспортная полиция сообщила им о смертельно опасном веществе. Транспортная полиция была в больнице, чтобы помочь с транспортировкой Навального медицинским бортом в Германию. Однако к моменту, когда самолет приземлился в аэропорту Омска, врачи заявили о нетранспортабельности господина Навального. По словам господина Калиниченко, из-за перепадов давления во время полета состояние пациента может ухудшиться.
Пресс-секретарь президента Дмитрий Песков заявил, что для транспортировки Алексея Навального нужно только решение лечащего врача. Накануне господин Песков заявлял, что Кремль готов содействовать переводу господина Навального для лечения за рубеж. В ответ лидер «Альянса врачей» Анастасия Васильева сообщила, что откликнулась на предложение. Спустя день господин Песков сказал, что не получал обращения как такового, но «лишняя бюрократия» в этом случае не нужна.
Накануне господин Песков заявлял, что Кремль готов содействовать переводу господина Навального для лечения за рубеж. В ответ лидер «Альянса врачей» Анастасия Васильева сообщила, что откликнулась на предложение. Спустя день господин Песков сказал, что не получал обращения как такового, но «лишняя бюрократия» в этом случае не нужна.
Напомним, 20 августа господин Навальный был госпитализирован в омскую больницу и впал в кому. Ему стало плохо во время перелета из Томска в Москву. Соратники оппозиционера уверены, что господина Навального отравили веществом, подсыпанным в чай, и настаивают на проведении анализов и лечении за границей. Состояние господина Навального улучшилось к утру 21 августа, при этом в течение дня оно остается нестабильным — это, по заявлению омских врачей, не позволяет перевезти его в другую клинику.
Обмен веществ у растений — Юнциклопедия
Обмен веществ с окружающей средой — главное условие жизни организма. Однако поглощение и выделение веществ — это только внешнее проявление обмена. Основу жизнедеятельности составляет внутриклеточное превращение веществ, называемое метаболизмом.
Основу жизнедеятельности составляет внутриклеточное превращение веществ, называемое метаболизмом.
В основе обмена веществ лежат два тесно связанных и взаимообусловленных процесса: ассимиляция и диссимиляция. Ассимиляция — усвоение питательных веществ, синтез специфических (характерных для данного организма) белков, нуклеиновых кислот, липидов, углеводов и других соединений. Процессы ассимиляции связаны с потреблением энергии. Диссимиляция — расщепление веществ, как поступающих извне, так и входящих в состав клеток организма. Освобождающаяся при их окислении энергия используется на разнообразные процессы жизнедеятельности. Кроме того, диссимиляция поставляет всевозможные промежуточные продукты, необходимые для синтетических реакций.
<addc>G</addc>
Растения по типу питания — автотрофные организмы. Особенность их обмена веществ в том, что они способны синтезировать все необходимые для жизнедеятельности органические вещества из минеральных. Для синтеза органических соединений из углекислого газа и воды растения используют солнечную энергию (см. Фотосинтез). Другой уникальный процесс, который происходит только в растениях, — перевод азота из минеральной формы в органическую, образование аминокислот, которые используются для биосинтеза белка. Эти так называемые незаменимые аминокислоты (лизин, валин, лейцин, изолейцин, метионин и др.) обязательно должны входить в рацион человека и животных.
Фотосинтез). Другой уникальный процесс, который происходит только в растениях, — перевод азота из минеральной формы в органическую, образование аминокислот, которые используются для биосинтеза белка. Эти так называемые незаменимые аминокислоты (лизин, валин, лейцин, изолейцин, метионин и др.) обязательно должны входить в рацион человека и животных.
Фосфорный обмен у растений сводится к образованию связи между остатками фосфорной кислоты и молекулой того или иного органического вещества. Значение образующихся при этом фосфорорганических соединений огромно. Это и аденозинтрифосфорная кислота (АТФ) — переносчик энергии в клетке, и нуклеиновые кислоты (ДНК, РНК), осуществляющие хранение и передачу наследственной информации, и фосфолипиды — компоненты биологических мембран и другие соединения.
Большое значение в обмене веществ имеют калий, кальций, магний, железо и другие элементы минерального питания и витамины. Частично они включаются в органические соединения. Главная же их роль регуляторная.
Таким образом, обмен веществ — это многочисленные согласованные химические процессы. Важную роль в их регуляции играют ферменты — специфические биокатализаторы белковой природы, в состав которых входят витамины и ионы металлов. Количество ферментов и их набор контролируется генетическим аппаратом. Не менее важное значение имеют клеточные мембраны. Они контролируют скорость поступления и выхода веществ, образуют внутри клетки микроскопические отсеки, в которых находятся определенные ферментные системы и происходит метаболизм. Согласованность обмена веществ в целостном организме обеспечивается деятельностью гормонов (см. Фитогормоны).
Любое заболевание, неполноценное питание приводят к нарушениям обмена веществ в организме, которые выражаются в изменении характера превращений веществ, в накоплении промежуточных, а иногда и не свойственных нормальному обмену продуктов. Лечение этих нарушений должно быть направлено на устранение причин, их вызывающих.
Публикация SCALLOP в журнале Nature Metabolism
Исследование белков сердечно-сосудистой системы у более чем 30 000 человек
Крупное протеогеномное исследование SCALLOP выявляет> 450 pQTL и несколько новых потенциальных мишеней для лекарственных препаратов
Иллюстрация Анн-Луизы Бергстрём (Moving Science ApS)
В новом важном исследовании, опубликованном в Nature Metabolism , консорциум SCALLOP идентифицировал локусы количественных признаков белка (pQTL) для 90 сердечно-сосудистых белков у более чем 30 000 человек, что привело к 451 pQTL (генетическим вариантам, связанным с уровнями экспрессии белка) для 85 белков.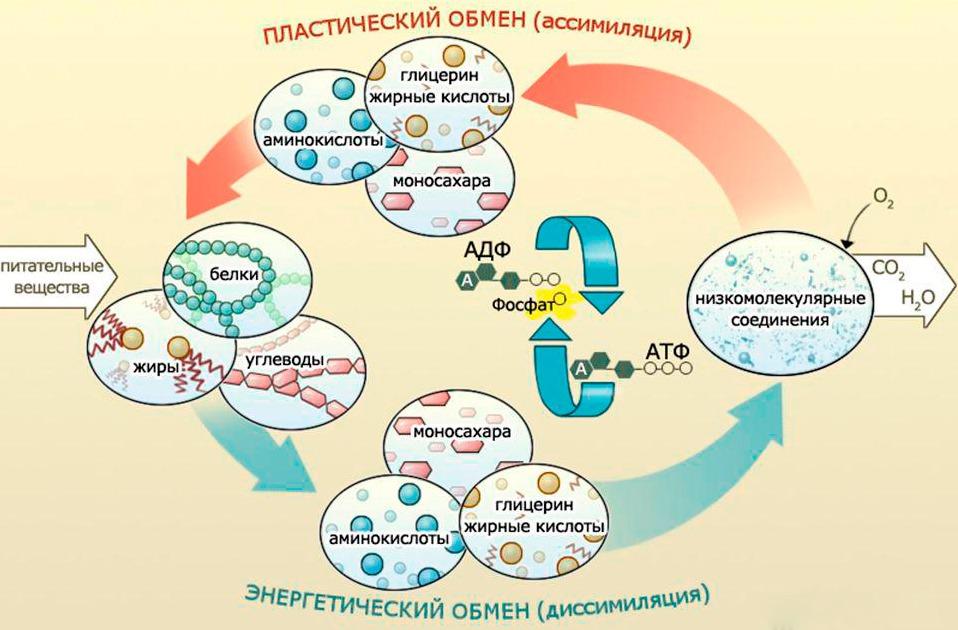 Данные были собраны из 15 когорт (исследования членов консорциума), все из которых использовали оригинальную панель Olink, разработанную для исследований сердечно-сосудистой системы, Olink CVD I.
Данные были собраны из 15 когорт (исследования членов консорциума), все из которых использовали оригинальную панель Olink, разработанную для исследований сердечно-сосудистой системы, Olink CVD I.
В дополнение к примерно 170 цис -pQTL с генетическими вариантами, проксимальными к генам, кодирующим пораженные белки, также были идентифицированы и картированы на регуляторные пути многие транс -pQTL, что в некоторых случаях можно было проверить ортогональными методами. Менделирующая рандомизация также применялась для идентификации 11 белков с новыми причинными доказательствами связи с человеческими заболеваниями, которые могут быть интересны для будущих исследований мишеней для лекарственных препаратов.
Сведения о цитировании
Folkersen, L., Gustafsson, S., Wang, Q. et al. Геномная и целенаправленная оценка лекарств 90 сердечно-сосудистых белков у 30 931 человека. Nat Metab 2, 1135–1148 (2020). https://doi. org/10.1038/s42255-020-00287-2
org/10.1038/s42255-020-00287-2
Ссылка
Что говорят авторы
Доктор Андерс Меларстиг, основатель SCALLOP и один из главных исследователей статьи, сказал об этом знаменательном исследовании следующее:
“ В начале консорциума SCALLOP мы не знали, возможно ли объединение генетики человека с протеомикой Olink в таком крупном масштабе, не говоря уже о том, приведет ли наш подход к новому пониманию биологии болезней.В свете этой статьи я теперь предсказываю, что мы увидим следующее поколение эффективных лекарств, появившихся на основе аналогичных подходов — с тщательно проведенными исследованиями по методу рандомизации по методу Менделя белковых биомаркеров. Вся заслуга в этой работе принадлежит группе разработчиков CVD-I и членам консорциума SCALLOP ».
Любые вопросы или запросы на перепечатку этой статьи следует направлять Андерсу Меларстигу
.Об авторе
Андерс Маларстиг, доктор философии, является директором по целевым наукам в Pfizer Worldwide Research and Development, а также аффилированным исследователем в Каролинском институте.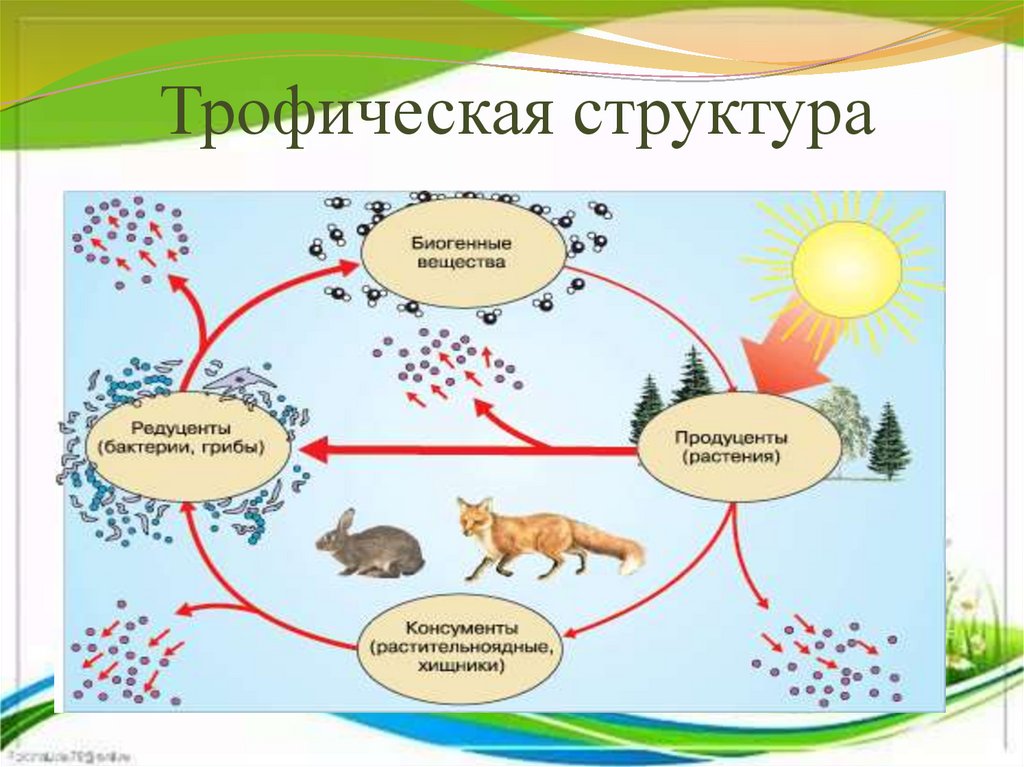
В компании Pfizer д-р Меларстиг отвечает за разработку и применение стратегий для новых лекарственных препаратов и точной медицины. Он является одним из основателей и главным контактным лицом консорциума SCALLOP.
Метаболизм, распределение и экспрессия остатков — Природа остатков в растениях | Набор инструментов для регистрации пестицидов | Продовольственная и сельскохозяйственная организация Объединенных Наций
Цель исследования
Метаболизм пестицида включает процессы транспорта или транслокации в организме, а также превращение в метаболиты или продукты разложения.
Исследования метаболизма проводятся для определения качественной метаболической судьбы активного ингредиента и выяснения его метаболического пути. Многие пестициды претерпевают изменения во время и после нанесения на растения или растительные продукты. Поэтому состав концевого остатка должен быть определен до разработки методологии анализа остатков и количественной оценки остатков.
В исследованиях метаболизма пестицид применяется к культурам в количествах и в течение периода времени, которые могут иметь место на практике, когда соединение используется для борьбы с вредителями (т.е. существующие или предполагаемые передовые методы ведения сельского хозяйства (GAP)). Что касается сельскохозяйственных культур, то корма и продукты питания собираются после интервала, ожидаемого в соответствии с надлежащей сельскохозяйственной практикой.
Дополнительную информацию об исследованиях метаболизма остатков в растениях можно найти в главах 3.2 и 5.3 Руководства по процедурам JMPR.
Примеры работ по этой теме представлены в главе 5 Учебного пособия JMPR.
Обстоятельства, при которых требуется исследование
Эти исследования требуются всякий раз, когда обрабатывается растение или растительный продукт, за исключением случаев, когда никакая часть растений или растительных продуктов не будет использоваться в качестве пищи или корма, или если не применяется «ситуация с нулевым остатком» (например, применение приманок).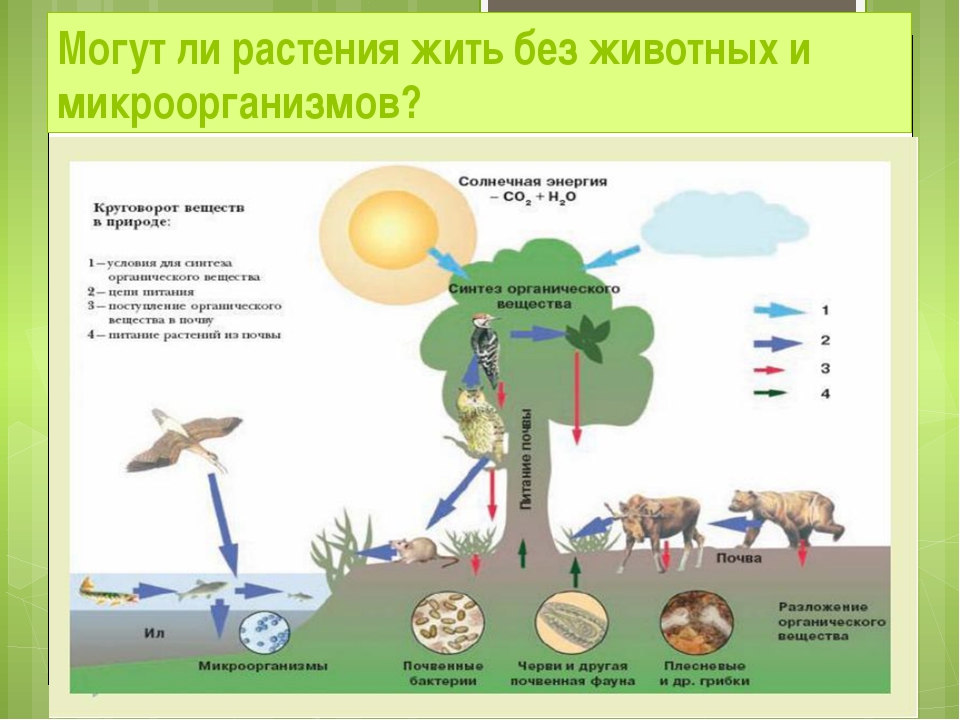
Тестируемый организм или субстрат
Исследование метаболизма должно быть представлено для каждого типа группы культур, для которой предлагается использование
. При изучении метаболизма сельскохозяйственных культур можно отнести к одной из пяти категорий:
- корнеплоды (корнеплоды, луковичные)
- листовые культуры (овощи Brassica, листовые овощи, стеблевые овощи, хмель)
- фруктов (цитрусовые, семечковые, косточковые мелкие фрукты, ягоды, виноград, банан, древесные орехи, плодовые овощи, хурма)
- зернобобовых и масличных культур (бобовые, бобовые, масличные, арахис, бобовые кормовые культуры, какао-бобы, кофейные зерна)
- зерновых (зерновые, травяные и кормовые культуры).
Одна культура из группы будет охватывать всю группу в целях метаболизма этих культур внутри группы. Чтобы экстраполировать метаболизм пестицида на все группы культур, следует провести исследования метаболизма как минимум на трех репрезентативных культурах (из пяти различных категорий культур).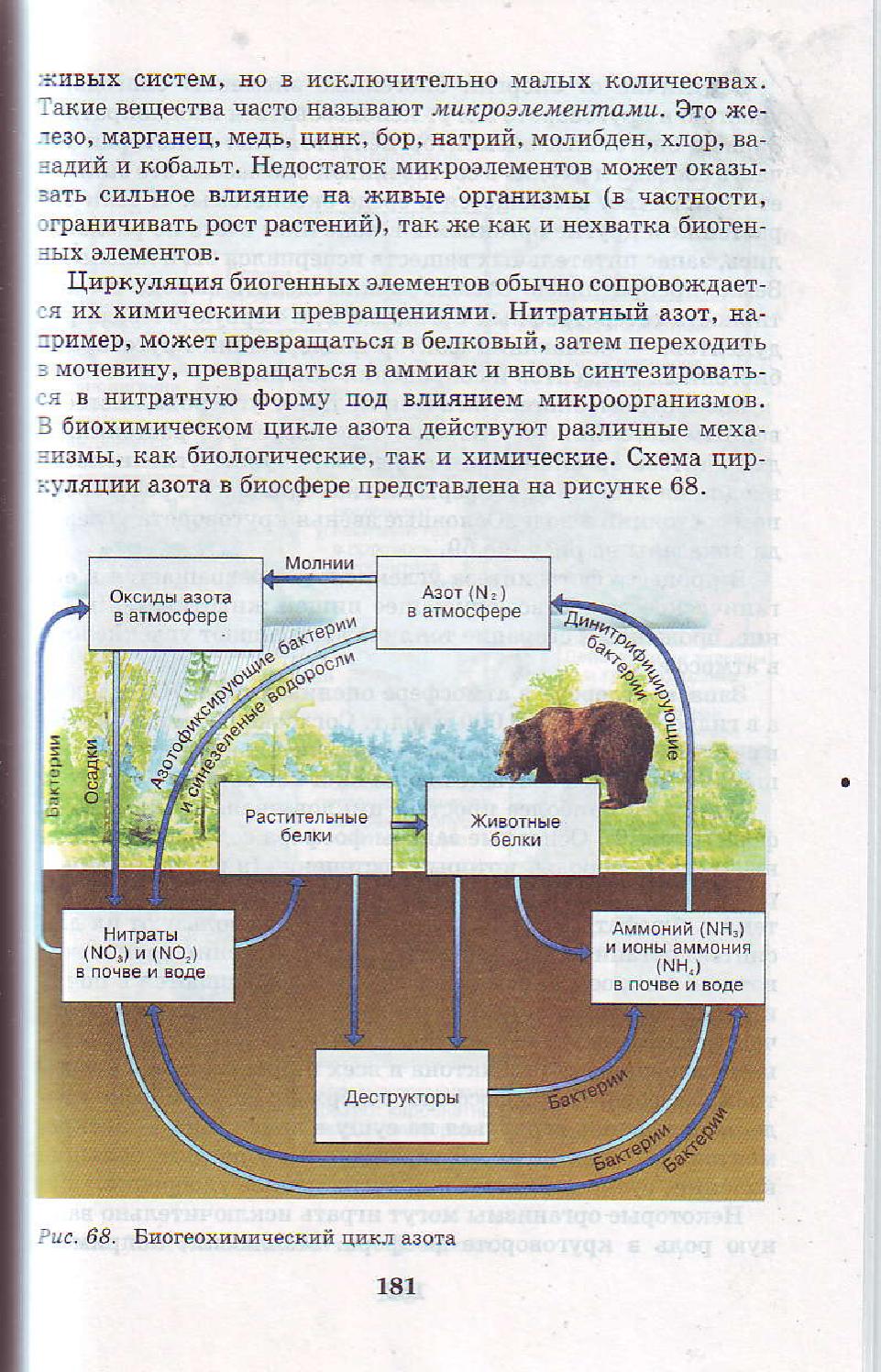 Если результаты этих трех исследований указывают на сопоставимый метаболический путь, то дополнительные исследования культур в двух других группах не потребуются.
Если результаты этих трех исследований указывают на сопоставимый метаболический путь, то дополнительные исследования культур в двух других группах не потребуются.
Тестовое вещество
Урожай должен быть обработан активным ингредиентом с радиоактивной меткой, предпочтительно содержащим ингредиенты рецептуры, типичные для конечного продукта, применяемого в поле.
Типичные конечные точки исследования
- Доля экстрагируемой и неизвлекаемой радиоактивности в съедобных тканях, как% от общих радиоактивных остатков (TRR) и мг эквивалента исходного соединения / кг материала образца;
- Часть охарактеризованной, но не идентифицированной радиоактивности в съедобных тканях и потенциальных пищевых продуктах;
- Идентификация метаболитов по систематическому химическому названию — для всех экстрагируемых остатков при> 10% TRR или при> 0.05 мг / кг;
- Характеристика метаболитов — для всех экстрагируемых остатков при TRR <10% или 0,01-0,05 мг / кг;
- Наличие метаболитов в конъюгированной форме, экстрагируемость конъюгированных метаболитов;
- Схематическая диаграмма метаболических путей у растений с кратким объяснением распределения и участвующих химических реакций;
- Передача радиоактивности в тканях растений. Идентификация транслоцированной радиоактивности (возможно, только некоторые метаболиты перемещаются из точки приложения).
- Доля остатков, которые можно удалить с помощью мытья поверхности.
- Распределение радиоактивности в кожуре и мякоти или различных частях растений (например, в кормах, соломе, зернах, шелухе;
Руководство по тестированию
Для определения природы остатков в растениях можно использовать следующие руководящие принципы испытаний:
Новое исследование роли макрофагов вне воспаления опубликовано в Nature Metabolism
Мириам Ауади, доктор философии (справа), руководитель группы интегрированного кардио-метаболического центра, отделение медицины, Худдинге, Каролинский институт, и Сесилия Моргантини, доктор медицины, доктор философии (слева), научный сотрудник лаборатории Ауади, научный сотрудник отделения эндокринологии в Каролинской больнице, недавно опубликованы исследование в Nature Metabolism.Исследование было проведено в сотрудничестве с учеными из AstraZeneca.
Сводка
В связи с ожирением, которым страдают 600 миллионов взрослых во всем мире, и неэффективностью существующих методов лечения, направленных на то, чтобы остановить быстро растущую распространенность связанных с ним метаболических заболеваний, таких как диабет 2 типа, существует острая необходимость в лучшем понимании этих заболеваний и разработке новых терапевтических стратегий. По данным шведского агентства здравоохранения, в Швеции 50% населения страдают избыточным весом или ожирением.
При ожирении неспособность жировой ткани накапливать избыточные питательные вещества в форме жира приводит к эктопическому отложению липидов в нежировой ткани, такой как печень. Это чрезмерное накопление липидов в печени, называемое стеатозом печени или неалкогольной жировой болезнью печени (НАЖБП), тесно связано с метаболической дисфункцией печени. Учитывая важную роль печени в регуляции метаболизма липидов и глюкозы, стеатоз печени может иметь большое влияние на обмен веществ в организме и, следовательно, способствовать метаболическим заболеваниям, таким как диабет 2 типа.
Роль воспаления печени в развитии метаболических заболеваний является предметом дискуссий на протяжении многих лет. Макрофаги — это клетки иммунной системы, которые играют важную роль в защите организма от патогенов. Однако недавно они были описаны как ключевые игроки в развитии метаболических заболеваний. Макрофаги присутствуют по всему телу, и они представляют наибольшую долю иммунных клеток в печени. Наши текущие знания о макрофагах печени ограничены из-за исследований с использованием неспецифических трансгенных моделей, методов истощения или транскриптомного анализа всей печени.
К сожалению, этот пробел в наших знаниях о биологии макрофагов печени привел к предположению, что макрофаги печени подвергаются провоспалительной активации, приводящей к чрезмерной выработке воспалительных факторов, которые способствуют резистентности к инсулину, вызванной ожирением. Эта догма побудила исследовательские группы изучить противовоспалительные препараты как потенциальные средства лечения метаболических заболеваний. В своем исследовании, недавно опубликованном в журнале Nature Metabolism, Morgantini et al. сделал наблюдение, что макрофаги печени на самом деле не вызывают воспалительных процессов при ожирении и инсулинорезистентности у трех видов: дрозофилы, мышей и человека.Используя определенные методы выделения макрофагов в сочетании с секвенированием следующего поколения, они обнаружили, что на транскриптомный профиль макрофагов печени сильно влияет ожирение, но независимо от их воспалительного статуса. «Эти неожиданные открытия изменили парадигму описания воспалительных макрофагов и воспаления как движущих сил метаболических заболеваний печени», — говорит ведущий автор исследования Сесилия Моргантини.
Для выяснения механизма, посредством которого макрофаги печени могут регулировать метаболизм в печени, Cecilia Morgantini et al.использовала запатентованную технологию, разработанную Мириам Ауади, для доставки миРНК и подавления генов в макрофагах специфическим для клеток и тканей образом. Используя эту уникальную технологию, они описали новый механизм, с помощью которого макрофаги могут нарушать метаболизм в печени и способствовать инсулинорезистентности за счет секреции невоспалительных факторов, таких как Igfbp7. «Было интересно наблюдать консервативный механизм, посредством которого IGFBP7, продуцируемый макрофагами печени у мышей и гемоцитами у Drosophila, может регулировать метаболизм глюкозы и липидов», — говорит Мириам Ауади, руководитель проекта.
У человека они обнаружили эволюционный регуляторный механизм, посредством которого IGFBP7 подвергается редактированию РНК с большей частотой у пациентов с ожирением и метаболическими нарушениями. Это событие редактирования РНК, которое происходит только у людей, но не у мышей или дрозофилы, приводит к генерации новой изоформы IGFBP7 с защитными свойствами. «Наше исследование раскрывает ранее неизвестную роль макрофагов помимо воспаления и открывает новые возможности для исследований в области иммунометаболизма и разработки новых терапевтических стратегий для метаболических и связанных заболеваний печени.Сейчас мы исследуем множество ролей макрофагов и их участие в заболеваниях печени. Это действительно захватывающее время, поскольку мы откроем новую биологическую функцию макрофагов примерно через сто лет после их открытия », — говорит Мириам Ауади.
Абстрактное графическое изображение
У мышей в условиях чувствительности к инсулину: IGFBP7, продуцируемый макрофагами печени, связывает печеночный рецептор инсулина IR и усиливает активацию AKT инсулином; в инсулинорезистентном состоянии, при котором AKT не реагирует на инсулин, IGFBP7 связывает IR с меньшим сродством, но все же может активировать путь ERK и индуцировать глюконеогенез и липогенез.У человека edIGFBP7 чаще встречается в инсулинорезистентном состоянии. EdIGFBP7 обладает более высокой способностью связывать IR и активировать AKT по сравнению с wtIGFBP7. IR: рецептор инсулина. ред: отредактировано; wt дикого типа.
Пункты списка
- Макрофаги печени не активируются при ожирении и инсулинорезистентности у дрозофилы, мышей и людей.
- Макрофаги печени продуцируют невоспалительные факторы, такие как IGFBP7, которые непосредственно регулируют метаболизм печени.
- IGFBP7, экспрессируемый макрофагами печени у человека, но не у мышей, подвергается редактированию РНК с высокой частотой у тучных инсулинорезистентных пациентов.
- Редактирование РНК IGFBP7 в макрофагах печени человека приводит к производству новой защитной изоформы с более высокой способностью индуцировать путь передачи сигналов инсулина.
Изображение: гибридизация in situ РНК печени. Изображение Igfbp7 (зеленый) в макрофагах печени, идентифицированных маркером Emr1 (F4 / 80) и DAPI. Изображение справа: эндогенная экспрессия гомолога Igfbp7 (Impl2) в гемоцитах (макрофагах) у дрозофилы.
Новая модель метаболизма основана на термодинамике и омиксе
Все живые существа состоят из углерода и сахаров, т.е.грамм. глюкоза — очень распространенный его источник. Следовательно, большинство клеток хорошо поедают сахар, используя ферменты для их переваривания посредством ряда химических реакций, которые превращают исходный сахар в различные компоненты клетки, включая аминокислоты, строительные блоки ДНК и жиры. Поскольку они помогают эффективно протекать реакциям метаболизма сахара, ферменты называются биокатализаторами.
Учитывая, насколько важны все ферменты для самой жизни, ученые построили несколько математических моделей, которые описывают, как клетки используют ферменты для преобразования сахара.Такие модели успешно использовались, например, для улучшения производства биотоплива 2-го поколения или определения целевых лекарств от малярии, но они не принимают во внимание метаболическую «стоимость» производства ферментов, которые катализируют все эти химические реакции.
Учет этого явления, называемого «экспрессией», является ключом к описанию многих других явлений, включая ферментацию пива и рост раковых клеток. Но все это в первую очередь зависит от точного моделирования механизмов выражения.
Теперь профессор Василий Хациманикатис из EPFL и Пьер Сальви, аспирант в своей лаборатории, разработали математическую модель, которая может эффективно моделировать экспрессию ферментов в живых клетках, а также связанные с этим метаболические затраты. Модель называется ETFL, что означает «поток выражений и термодинамики», и ее точность основана на учете как биохимии, так и термодинамики — набора физико-химических законов, описывающих, как энергия течет в системах. Объединив это с математическими инструментами из области оптимизации, исследователи смогли значительно повысить точность прогнозов модели.
«Эта интеграция метаболизма, экспрессии и термодинамики является первой в своем роде и в 10–100 раз быстрее, чем предыдущие современные модели, в которых нет термодинамики», — говорит Салви.
Чтобы еще больше повысить свою предсказательную силу, модель ETFL была разработана с учетом широкого спектра измерений, сделанных с помощью огромного поля «омиков», которое измеряет такие характеристики клеток, как экспрессия генов, профили белков и т. Д. «Наш алгоритм может могут использоваться для улучшения производства биохимических продуктов или для точного прогнозирования метаболизма раковых клеток », — говорит Салви.«Это также может открыть двери для приложений в персонализированной медицине».
###
Номер
Пьер Сальви, Василий Хациманикатис. Формула ETFL позволяет интегрировать мультиомиксы в совместимые с термодинамикой модели метаболизма и экспрессии. Nature Communications 13 января 2020 г. DOI: 10.1038 / s41467-019-13818-7
Заявление об ограничении ответственности: AAAS и EurekAlert! не несут ответственности за точность выпусков новостей, размещенных на EurekAlert! посредством участвующих организаций или для использования любой информации через систему EurekAlert.
Наука метаболизма
Перейти к
Энергетические превращения в метаболизме
Энергия для различных функций человеческого тела поступает от молекул питательных веществ, которые были расщеплены в процессе, называемом метаболизмом. Метаболизм состоит из двух основных частей: анаболизма (наращивание, также называемое биосинтезом) и катаболизм (разрушение).
Точный характер катаболических реакций отличается от организма к организму, и организмы можно классифицировать на основе их источников энергии и углерода:
- Органотрофы, в качестве источника энергии используются органические источники
- в литотрофах, используются неорганические субстраты
- у фототрофов, солнечный свет используется как химическая энергия
- у гетеротрофов в качестве источника энергии используются органические соединения (соединения не синтезируются организмом, а получаются с пищей).
Основные общие реакции катаболизма включают окислительно-восстановительные (окислительно-восстановительные) реакции, которые включают перенос электронов от восстановленных молекул-доноров, таких как органические молекулы, вода, аммиак, сероводород или ионы двухвалентного железа, к молекулам-акцепторам, таким как кислород, нитрат или сульфат.
У людей и животных (гетеротрофов) окислительно-восстановительные реакции включают разложение сложных органических молекул на более простые молекулы, такие как диоксид углерода и вода. В фотосинтезирующих организмах, таких как растения и цианобактерии (фототрофы), эти реакции переноса электронов не выделяют энергию.Эти реакции просто помогают накапливать энергию, поглощаемую солнечным светом.
Энергия из пищевых продуктов
У людей энергия тела поступает из жиров, углеводов и белков, содержащихся в пище, что делает нас гетеротрофами. Из трех органических молекул жир является наиболее концентрированным источником энергии, поскольку дает более чем в два раза больше энергии для данного веса, чем белок или углевод.
Энергетические потребности обычно выражаются в калориях или килокалориях.Килокалория (ккал) — это количество тепловой энергии, необходимое для повышения температуры одного килограмма воды на один градус Цельсия. Жир обеспечивает наибольшую энергию на массу — 9 ккал / г, за ним следуют белки и углеводы (4 ккал / г) и гидратированные углеводы (1,3 ккал / г). Липиды расщепляются на жирные кислоты, белки — на аминокислоты, а углеводы — на глюкозу. Затем эти продукты подвергаются окислительно-восстановительным реакциям.
Базальный уровень метаболизма (BMR) — это тепло, отводимое телом в состоянии покоя при нормальной температуре.Среднестатистическому человеку требуется 2000–2400 калорий в день, тогда как крупному человеку, выполняющему тяжелую работу, может потребоваться до 6000 калорий в день. Энергетические потребности детей сильно различаются в зависимости от их возраста, размера и уровня активности.
Катаболические процессы
В целом, оба процесса метаболизма, катаболизма и анаболизма, должны происходить одновременно, потому что катаболизм обеспечивает энергию, необходимую для анаболизма. Тело использует энергию для множества функций.Энергия необходима для выполнения механической работы, которая включает изменение положения или ориентации части тела или самой клетки. Это включает движение мышц. Кроме того, существует молекулярный транспорт и синтез биомолекул.
АТФ (аденозинтрифосфат) — это энергетическая молекула, передающая химическую энергию в клетках человека. Как правило, энергия для синтеза молекул АТФ должна поступать из молекул пищи. АТФ в основном синтезируется в митохондриях клеток, а некоторые дополнительные АТФ синтезируются в цитоплазме.
АТФ-АДФ-цикл. Аденозинтрифосфат (АТФ) — это органическое химическое вещество, обеспечивающее клетки энергией. внутриклеточная передача энергии. Аденозиндифосфат (АДФ) — это органическое соединение, отвечающее за метаболизм в клетке. Кредит иллюстрации: Designua / Shutterstock
Катаболизм можно разделить на три основных этапа:
- Пищеварение — большие органические молекулы (белки, липиды, углеводы) перевариваются на более мелкие компоненты (жирные кислоты, аминокислоты и глюкоза соответственно) вне клетки пищеварительными ферментами, такими как глюкозидгидролазы для углеводов и пепсин для белков.Это происходит в пищеварительном тракте человека.
- Высвобождение энергии — более мелкие компоненты перемещаются в клетки активными транспортными белками и преобразуются в более мелкие молекулы, как правило, ацетил-кофермент А (ацетил-КоА), который выделяет некоторую энергию. В цитоплазме глюкоза далее трансформируется в пируват, который вызывает синтез двух молекул АТФ. У людей эти небольшие молекулы транспортируются через ткани пищеварительной системы в ткани кровообращения, а затем распределяются по телу туда, где они необходимы для выработки энергии.
- Производство АТФ — в цикле лимонной кислоты (также называемом циклом Креба или TCA) ацетильная группа КоА окисляется до воды и диоксида углерода. Высвобождаемая при этом энергия сохраняется в АТФ за счет восстановления кофермента адениндинуклеотида (НАД +) до НАДН в цепи переноса электронов. Этот процесс называется окислительным фосфорилированием и выделяет углекислый газ в качестве побочного продукта.
Различные органические молекулы обеспечивают различное количество АТФ. Каждая молекула жирной кислоты высвобождает более 100 молекул АТФ, а каждая молекула аминокислоты высвобождает около 40 молекул АТФ.
Аминокислоты могут быть окислены до кетокислот путем удаления аминогруппы, которая поступает в цикл мочевины. Затем именно кетокислота входит в цикл лимонной кислоты и способствует выработке АТФ.
При отсутствии кислорода (анаэробные условия) вырабатывается меньше АТФ. Цикл гликолиза производит лактат через фермент лактатдегидрогеназу, который повторно окисляет НАДН до НАД + для повторного использования в гликолизе. Жиры могут расщепляться до глицерина, который вступает в процесс гликолиза.
Ключевые биомолекулы
Хотя белки, углеводы и жиры являются важными источниками катаболических реакций, они также необходимы для различных других функций организма.Некоторые из них также вырабатываются анаболизмом в дополнение к ДНК. Минералы также важны для метаболических целей.
Белки
Белки состоят из аминокислот. В процессе синтеза белка аминокислоты соединяются в длинные цепи, называемые полипептидными цепями. Они соединены пептидными связями. Полипептидные цепи подвергаются дальнейшей модификации с образованием белков.
Некоторые белки используются для формирования структуры клеток и тканей, а многие другие представляют собой ферменты, катализирующие различные химические реакции в организме.Белки также важны для передачи сигналов клеток, иммунных ответов, клеточной адгезии, активного транспорта через мембраны и клеточного цикла.
Коферменты — это небелковые вещества (например, минералы или металлы), которые опосредуют несколько химических реакций в метаболических путях тела. Они подпадают под несколько основных типов реакций, которые включают перенос функциональных групп.
Коэнзимы также помогают в передаче энергии. Одним из центральных коферментов является аденозинтрифосфат (АТФ), энергетическая валюта клеток.В клетках лишь небольшое количество АТФ, но он постоянно регенерируется. Другие включают никотинамидадениндинуклеотид (НАДН), производное витамина B3, которое действует как акцептор водорода.
Сотни отдельных типов дегидрогеназ удаляют электроны со своих субстратов и восстанавливают NAD + до NADH. Эта восстановленная форма кофермента затем является субстратом для любой из редуктаз в клетке, которым необходимо восстанавливать свои субстраты. НАДН существует в клетке в двух связанных формах: НАДН и НАДФН.Форма НАД + / НАДН более важна в катаболических реакциях, тогда как НАДФ + / НАДФН используются в анаболических реакциях.
Углеводы
Углеводы — основной источник энергии в организме. Углеводы — это альдегиды или кетоны с прямой цепью с гидроксильными группами, которые могут существовать в виде прямых цепей или колец.
Углеводы в изобилии в природе и играют несколько ролей в живых организмах. Их можно преобразовать в гликогены и использовать в качестве источников энергии в качестве структурных компонентов (целлюлоза в растениях, хитин у животных) и в качестве прямого источника энергии (глюкоза).
Липиды
Липиды — важные биохимические вещества, выполняющие разностороннюю функцию в организме. Они образуют структурную часть биологических мембран, например клеточную мембрану, или используются в качестве источника энергии. Жиры представляют собой большую группу соединений, содержащих жирные кислоты и глицерин. Продукция часто принимает форму стероидов, таких как холестерин, еще один основной класс липидов, производимых в клетках.
Иллюстрация липидов и жиров.Кредит: Naeblys / Shutterstock
Нуклеотиды
Нуклеотиды помогают в образовании ДНК и РНК. ДНК и РНК — это длинные цепочки нуклеотидов, критически важные для хранения и использования генетической информации. РНК и ДНК также кодируют синтез белка. Кроме того, нуклеотиды могут действовать как коферменты в реакциях передачи метаболических групп.
Кофакторы и минералы в метаболизме
Органические соединения (белки, липиды и углеводы) содержат большую часть углерода и азота у человека, тогда как большая часть кислорода и водорода присутствует в форме воды.
Есть несколько минералов и витаминов, которые играют решающую роль в метаболизме. Среди них часто встречаются натрий и калий. Другие важные минералы включают кальций, фосфор, железо, ионы хлора, медь, цинк, фтор, йод и магний. Металлические микроэлементы попадают в организм с помощью определенных переносчиков.
Катионы часто действуют как кофакторы, которые прочно связаны с определенным белком. Кофакторы ферментов можно модифицировать во время катализа, но кофакторы всегда возвращаются в свое исходное состояние после того, как катализ прошел.
Термодинамика метаболизма
Метаболические процессы — это химические реакции, которые часто включают выделение тепла. Клеточный метаболизм сочетает спонтанные процессы катаболизма с неспонтанными процессами анаболизма. С термодинамической точки зрения, метаболизм поддерживает баланс.
Химические реакции классифицируются как экзэргонические или эндергонические. Это означает, что реакция может либо выделять энергию, полезную для работы (экзэргоническая реакция), либо требовать энергии для продолжения (эндергоническая реакция).Следовательно, производство АТФ во время катаболизма является экзэргоническим, тогда как анаболизм — эндергонической реакцией.
Контроль метаболизма
Метаболические пути сложны и взаимозависимы. При изменении окружающей среды необходимо точно регулировать реакции метаболизма, чтобы поддерживать постоянный набор условий внутри клеток — состояние, называемое гомеостазом. Контроль метаболических путей также позволяет организмам реагировать на сигналы и взаимодействовать с окружающей средой.
Уровни регуляции обмена веществ
Есть несколько уровней регуляции метаболизма.Что касается внутренней регуляции метаболических путей, реакции саморегулируются, чтобы отвечать на изменения уровней субстратов или продуктов. Например, уменьшение количества продукта может увеличить метаболический путь. Это называется механизмом обратной связи.
Внешний контроль включает клетку в многоклеточном организме, изменяющую свой метаболизм в ответ на сигналы от других клеток. Сигналы достигают путей через растворимые посредники, такие как гормоны и факторы роста. Например, гормон инсулин из бета-клеток поджелудочной железы вырабатывается в ответ на повышение уровня глюкозы в крови.Связывание гормона с рецепторами инсулина на клетках затем активирует каскад протеинкиназ, которые заставляют клетки поглощать глюкозу и превращать ее в запасные молекулы, такие как жирные кислоты и гликоген.
Регуляция углеводного обмена
Гомеостаз глюкозы — сложное взаимодействие метаболических путей, но он жизненно важен для живых организмов. Эти процессы либо увеличивают, либо уменьшают концентрацию глюкозы в крови, но работают вместе, чтобы поддерживать оптимальный уровень.
Глюкоза получается из углеводов, поступающих с пищей. Углеводы перевариваются до простых сахаров: глюкозы, фруктозы и галактозы. Эти сахара всасываются в кишечнике и транспортируются в печень через воротную вену. После этого печень превращает фруктозу и галактозу в глюкозу. Повышение уровня глюкозы в крови стимулирует высвобождение инсулина из бета-клеток островков Лангерганса в поджелудочной железе.
Инсулин — единственный гормон, который снижает уровень глюкозы в крови, и делает это за счет активации механизмов транспорта глюкозы и метаболических путей использования глюкозы в различных тканях тела.Таким образом, инсулин подавляет пути образования глюкозы.
Инсулин и глюкоза. Бета-клетки (в поджелудочной железе) выделяют инсулин в кровеносный сосуд. Инсулин стимулирует всасывание глюкозы в скелетных мышцах. Крупным планом поджелудочная железа и островки Лангерганса. Кредит изображения: Designua / Shutterstock
Инсулин стимулирует захват глюкозы (мышечной и жировой тканью), гликолиз, гликогенез (образование гликогена из свободной глюкозы) и синтез белка. И наоборот, инсулин подавляет глюконеогенез (образование глюкозы из жирных кислот и т. Д.), липолиз (расщепление жирных кислот), протеолиз (расщепление белков) и кетогенез (образование кетоновых тел).
Нарушения обмена веществ и манипуляции
Метаболические пути сложны и часто взаимозависимы. Любое изменение проводящих путей может вызвать сложные расстройства. Например, нарушение баланса гомеостаза глюкозы и углеводного обмена связано с диабетом. Это делает исследование метаболических путей и манипулирование ими важными для клинической диагностики и лечения.
Исследования метаболических путей и нарушений
Одним из наиболее полезных инструментов для исследования несбалансированных метаболических путей является оценка конечных продуктов метаболических путей. Например, при сахарном диабете не хватает гормона инсулина, который поддерживает нормальный уровень сахара в крови, и оценка уровня сахара в крови натощак (после 8-10 часов отсутствия еды) и после приема пищи (через 2 часа после приема пищи) помогает диагноз.
Другие методы, с помощью которых метаболические пути могут быть исследованы в ходе исследований (но не в клинической практике), заключаются в использовании радиоактивных индикаторов или метаболомики.Радиоактивные индикаторы могут помочь определить пути от прекурсоров к конечным продуктам, идентифицируя радиоактивно меченные промежуточные продукты и продукты. После оценки помеченных химических веществ ферменты, катализирующие эти химические реакции, могут быть очищены, а их кинетика и реакция на ингибиторы могут быть исследованы. Метаболомические исследования могут предоставить информацию о структуре и функциях простых метаболических путей. Однако эти исследования могут быть неадекватными при применении к более сложным системам, таким как метаболизм целой клетки.Это потому, что метаболические сети внутри клетки содержат тысячи различных ферментов и сложных сетей. Геномы показывают, что существует около 45 000 генов, которые могут кодировать ферменты и другие кофакторы метаболических путей.
Манипуляции с метаболическими путями
С момента появления геномных исследований было разработано управление экспрессией генов на основе протеомных исследований и исследований ДНК-микрочипов. Многие из врожденных метаболических нарушений лечили с помощью генной терапии и манипуляции с генами, кодирующими дефектные ферменты и белки в метаболических путях.
На основе генетики создана модель метаболизма человека, которая будет определять будущие открытия лекарств и биохимические исследования. Эти модели сейчас используются в сетевом анализе для классификации болезней человека по группам, которые имеют общие белки или метаболиты.
Метаболическая инженерия — это целенаправленное и целенаправленное изменение метаболических путей в организме. Это помогает понять и использовать клеточные пути для химической трансформации, передачи энергии и супрамолекулярной сборки.Метаболическая инженерия опирается на принципы химической инженерии, вычислительных наук, биохимии и молекулярной биологии для разработки и анализа путей.
Метаболическая инженерия использует организмы, такие как дрожжи, растения или бактерии, которые были генетически модифицированы, чтобы сделать их более полезными в биотехнологии и помочь в производстве лекарств, таких как антибиотики или промышленные химические вещества 1,3-пропандиол и шикимовая кислота. Эти модификации направлены на снижение количества энергии, используемой для производства продукта, повышение урожайности и сокращение образования отходов.
Evolution
Метаболические пути включают несколько длительных и сложных молекулярных и химических реакций, которые были сохранены в ходе эволюции, так что даже самые простые организмы разделяют некоторые общие метаболические пути со сложными организмами, такими как человек.
Сохранение этих древних путей может быть результатом того, что эти реакции являются оптимальным решением их конкретных метаболических проблем. Например, гликолиз и цикл лимонной кислоты производят свои конечные продукты высокоэффективно и за минимальное количество этапов.Эта экономия и оптимальная ситуация привели к развитию этих реакций с течением времени.
Развитие цикла лимонной кислоты
Эволюционное происхождение цикла лимонной кислоты уже давно является модельным примером в понимании происхождения и эволюции метаболических путей. Хотя химические этапы цикла сохраняются в природе в неизменном виде, различные организмы по-разному используют ее химию. В некоторых случаях организмы используют только избранные части цикла.
Было предложено более одной гипотезы для объяснения эволюции метаболических путей. К ним относятся последовательное добавление новых ферментов к гораздо более коротким более ранним путям, а также привлечение уже существующих ферментов и их сборка в новый путь реакции.
Геномные исследования показали, что ферменты пути, вероятно, имеют общее происхождение, предполагая, что многие пути эволюционировали пошагово. По мере развития путей новые функции были созданы из ранее существовавших этапов пути.
Альтернативная гипотеза исходит из исследований, отслеживающих эволюцию структур белков в метаболических сетях. Это показывает, что ферменты задействованы повсеместно. Эти процессы набора приводят к эволюционной ферментативной мозаике.
Существует также вероятность того, что некоторые части метаболизма могут существовать в виде «модулей», которые можно повторно использовать в различных путях и выполнять аналогичные функции на разных молекулах. Кроме того, со временем теряются некоторые функции и части путей, которые не являются необходимыми для выживания.
История
Метаболизм и метаболические пути изучались на протяжении нескольких столетий, и в ходе ранних исследований они перешли от изучения целых животных к изучению индивидуальных метаболических реакций в современной биохимии и молекулярной биологии.
Ранние метаболические исследования
Метаболические исследования были проведены еще в тринадцатом веке Ибн ан-Нафисом (1213-1288), который заявил, что «тело и его части находятся в непрерывном состоянии растворения и питания, поэтому они неизбежно претерпевают постоянные изменения.«
Первые зарегистрированные и более сложные исследования метаболизма начались в последние десятилетия шестнадцатого века. Именно в это время прямое наблюдение было дополнено инструментами, которые позволяли количественную оценку и, следовательно, проверку в науках, особенно в биологических системах. В медицине прогресс зависел от применения точных наук, таких как химия, математика и физика, к изучению функций.
Санторио Санкториус (1561–1636) внес свой вклад в изучение метаболизма, изучая потоотделение.Его усилия на протяжении многих лет экспериментов привели к исследованиям метаболического баланса. Первые контролируемые эксперименты по метаболизму человека были опубликованы Санторио Санторио в 1614 году в его книге «Ars de statica medecina». В своих экспериментах он взвешивался до и после еды, сна, работы, секса, голодания, питья и выделения. Он обнаружил, что большая часть еды, которую он принимал, была потеряна из-за того, что он назвал «нечувствительным потом».
Первоначальные исследования метаболизма проводились на живых животных или людях-добровольцах.Механизмы этих метаболических процессов еще не были идентифицированы, и считалось, что жизненная сила оживляет живую ткань.
Метаболические исследования XIX века
Это было в 19 веке, когда Луи Пастер экспериментировал с дрожжевым брожением сахара в спирт, он заметил, что брожение катализируется веществами внутри дрожжевых клеток, которые он назвал «ферментами».
Это открытие, наряду с публикацией Фридриха Велера в 1828 году о химическом синтезе мочевины, заложило основу для изучения органических соединений и химических реакций, обнаруженных в клетках, составляющих метаболические пути.
Исследования метаболизма ХХ века
Эдуард Бухнер в начале 20 века продвинул знания дальше, открыв ферменты. Он обнаружил, что изучение химических реакций метаболизма отличается от биологических исследований клеток, и начал понимать основы биохимии. В начале 20 века биохимические исследования бурно развивались.
Наиболее заметными открытиями стало открытие цикла лимонной кислоты Гансом Кребсом, внесшим огромный вклад в изучение метаболизма.Он открыл цикл мочевины, а позже, работая с Гансом Корнбергом, цикл лимонной кислоты и цикл глиоксилата.
Текущие исследования метаболизма
Метаболизм сейчас изучается с помощью методов молекулярной биотехнологии и геномики. Обычно используются такие инструменты, как хроматография, дифракция рентгеновских лучей, ЯМР-спектроскопия, мечение радиоизотопов, электронная микроскопия и моделирование молекулярной динамики. Эти методы позволили открыть и детально проанализировать метаболические пути и генетические основы метаболических нарушений.
Исследования, проведенные за последние два столетия, также сделали успехи в понимании метаболизма лекарств и метаболизма ксенобиотиков.
Источники
- Berg, J.M. et al. (2002). Глава 22 из Биохимия . 5-е издание. https://www.ncbi.nlm.nih.gov/books/NBK21173/
- Макки, Т., и Макки, Дж. Р. (2015). Глава 8 из Молекулярная основа жизни . 6-е издание. www.oup.com/…/McKee_Chapter8_Sample.pdf
- Да Поян А.T., et al. (2010). Использование питательных веществ у людей: пути метаболизма. Природное образование . 3 (9): 11. www.nature.com/…/nutrient-utilization-in-humans-metabolism-pathways-14234029
- Chemistry LibreTexts. Катаболизм белков. (2014). chem.libretexts.org/…/26.09%3A_The_Catabolism_of_Proteins
- Alves, R. et al. (2002). Эволюция ферментов в метаболизме: сетевая перспектива. Журнал молекулярной биологии .324 (2): 387. https://doi.org/10.1061/S0022-2836(02)01157-9
- Forst, C.V., и Schulten, K. (1999). Эволюция метаболизма: новый метод сравнения метаболических путей с использованием информации геномики. Журнал вычислительной биологии. 6 (3): 343-360. https://doi.org/10.1089/106652799318319
- Экноян Г. (1999). Санторио Санкториус — отец-основатель исследований метаболического баланса. Американский журнал нефрологии. 19 (2): 226-233. https: // doi.org / 10.1159 / 000013455
- Манчестер, К.Л. (1995). Луи Пастер — Случай и подготовленный разум. Тенденции в биотехнологии. 13 (12): 511-515. https://doi.org/10.1016/S0167-7799(00)89014-9
- Корнберг, Х. (2000). Кребс и его троица циклов. Nature Reviews Молекулярная клеточная биология. 1 (3): 225-228. http://www.ncbi.nlm.nih.gov/pubmed/11252898
Дополнительная литература
Этот сайт использует файлы cookie для повышения производительности.Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.Чтобы принять файлы cookie с этого сайта, используйте кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.Вы должны отключить приложение при входе в систему или уточнить у системного администратора.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует от сайта создания нового сеанса для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в cookie-файлах может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Frontiers | Влияние микробиома кишечника на метаболизм хозяина посредством регуляции высвобождения гормона кишечника
Введение
Желудочно-кишечный тракт (ЖКТ) является хозяином очень сложной микробной экосистемы, состоящей из бактерий, дрожжей, грибов, бактериофагов и других вирусов (Scarpellini et al., 2015), а также простейшие и археи (Koskinen et al., 2017; Laforest-Lapointe, Arrieta, 2018). Комменсальные бактерии, далее называемые микробиотой кишечника, встречаются по всей длине желудочно-кишечного тракта и в наибольшей плотности в слепой и толстой кишке и вместе со своими генами и генными продуктами (вместе именуемыми микробиомом кишечника) выполняют несколько функций. которые сильно влияют на физиологию хозяина. Мало того, что микробиота кишечника играет решающую роль в модуляции иммунной защиты хозяина (Belkaid and Hand, 2014) и функции мозга (Rogers et al., 2016), он также играет роль в регуляции метаболизма хозяина (Turnbaugh et al., 2006; Turnbaugh, Gordon, 2009; Vrieze et al., 2012, 2014; Le Chatelier et al., 2013; Nieuwdorp et al., 2014 ; Blaut, 2015; Hartstra et al., 2015; Seeley et al., 2015; Suarez-Zamorano et al., 2015; Aguirre et al., 2016; Pedersen et al., 2016; Molinaro et al., 2017; Rodrigues et al., 2017; Brubaker, 2018; Fabbiano et al., 2018). Это хорошо иллюстрируется передачей микробиоты от худых и страдающих ожирением человеческих близнецов стерильным (GF) мышам, лишенным нативного кишечного микробиома, что приводит к передаче метаболического фенотипа хозяина (Ridaura et al., 2013). Истощение микробиоты у мышей обеспечивает значительную защиту от метаболической дисрегуляции, вызванной диетой с высоким содержанием жиров, такой как ожирение, непереносимость глюкозы и инсулинорезистентность (Suarez-Zamorano et al., 2015), которые являются отличительными признаками метаболических заболеваний, включая диабет 2 типа ( T2D). Хотя бактериально-опосредованное воспаление связано с пагубными метаболическими эффектами у мышей (Lam et al., 2012; Molinaro et al., 2017), основные механизмы, с помощью которых микробиота кишечника влияет на метаболизм, до сих пор полностью не изучены.Микробиом кишечника вносит значительный вклад в сбор энергии хозяина, превращая недоступные источники питательных веществ, такие как полисахариды растений и другие сложные углеводы, в легко усваиваемые метаболиты (Tremaroli and Backhed, 2012). Более того, была установлена ключевая связь между микробиомом кишечника и высвобождением нескольких гормонов кишечника, которые являются важными регуляторами периферического метаболизма.
В слизистой оболочке кишечника специализированные энтероэндокринные (ЭЭ) клетки синтезируют и секретируют несколько гормонов, которые способствуют ряду ключевых физиологических процессов.В совокупности клетки ЕЕ составляют самый большой эндокринный орган в организме (Ahlman and Nilsson, 2001), несмотря на то, что они составляют менее 1% от общей популяции эпителиальных клеток кишечника. Было определено большое количество субпопуляций клеток ЕЕ, в основном на основании их профиля экспрессии гормонов (Fothergill et al., 2017). Клетки ЕЕ обладают способностью ощущать питательную среду просвета кишечника и по-разному реагируют на многие пищевые соединения и условия просвета кишечника.Растущее количество доказательств подчеркивает, что микробиом кишечника влияет на высвобождение гормона клеток ЕЕ, что имеет последующие последствия для метаболизма хозяина и прогрессирования метаболических заболеваний (рис. 1). Также есть недавние данные о том, что микробиологическое высвобождение гормонов кишечника может влиять на другие типы клеток ЕЕ (Lund et al., 2018), демонстрируя сложность взаимосвязи между микробиомом кишечника, высвобождением гормонов кишечника и метаболизмом хозяина. Этот обзор направлен на описание того, как кишечная микробиота и их метаболиты могут влиять на метаболизм хозяина посредством регуляции высвобождения гормонов кишечника.
Рисунок 1. Микробная регуляция метаболизма хозяина посредством высвобождения гормона кишечника. Микробиота кишечника передает сигнал соседним энтероэндокринным (ЕЕ) клеткам через ряд микробных метаболитов, включая короткоцепочечные жирные кислоты и вторичные желчные кислоты, а также структурные компоненты. Эти клетки EE выделяют важные метаболически активные гормоны, такие как GLP-1, PYY, GIP, 5-HT и CCK, которые влияют на ключевые метаболические процессы, включая метаболизм глюкозы, чувствительность к инсулину, ожирение и пищевое поведение.В свою очередь, диетические компоненты влияют на состав микробиоты кишечника, что может иметь дальнейшие последующие последствия для секреции гормонов кишечника и метаболизма хозяина.
Сигнал микробных метаболитов с клетками-хозяевами
Жирные кислоты с короткой цепью
Микробиота кишечника производит множество метаболитов за счет расщепления неперевариваемых углеводов (рис. 2). Наиболее распространенными из этих метаболитов являются ацетат, пропионат и бутират короткоцепочечных жирных кислот (SCFAs) (Topping and Clifton, 2001), которые существуют в соотношении приблизительно 3: 1: 1 в просвете кишечника человека, соответственно (Cummings et al. al., 1987; Mowat and Agace, 2014), однако это соотношение, по крайней мере частично, зависит как от диеты, так и от микробного состава. Судьба SCFAs, полученных из этих бактерий, существенно различается: ацетат легко всасывается в кровоток для распределения в периферических тканях. С другой стороны, пропионат метаболизируется в печени при абсорбции (Koh et al., 2016), в то время как большая часть бутирата потребляется локально колоноцитами в качестве основного источника топлива. В то время как большинство бактериальных SCFAs присутствуют в толстой кишке, меньшие количества также были обнаружены в подвздошной кишке свиней в результате цекоилеального рефлюкса (Cuche and Malbert, 1999) и, в меньшей степени, в проксимальном отделе тонкой кишки. .Относительное количество SCFA также, вероятно, будет различаться по длине кишечника в результате специфического для региона микробного состава, воздействия субстрата и абсорбции (Gu et al., 2013). Например, гены, кодирующие пути метаболизма углеводов, обогащены представителями типа Bacteroidetes, тогда как гены, кодирующие пути метаболизма желчных кислот, обогащены устойчивостью к желчным кислотам Firmicutes (David et al., 2014).
Рисунок 2. Микробная передача сигналов энтероэндокринным клеткам.Резидентная микробиота в просвете кишечника передает сигнал энтероэндокринным (ЭЭ) клеткам несколькими путями. Во-первых, микробиота превращает неперевариваемые углеводы в жирные кислоты с короткой цепью (SCFA), которые, в свою очередь, передают сигнал клеткам EE через рецепторы свободных жирных кислот 2 или 3 (FFAR2 / 3) или путем активации ядерных гистоновых деацетилаз (HDAC). Во-вторых, микробиота превращает первичные (1 °) желчные кислоты во вторичные (2 °) желчные кислоты, такие как дезоксихолат, которые затем передают сигнал клеткам EE через мембранный рецептор желчных кислот, связанный с G-белком, TGR5 или ядерный FXR.Наконец, структурные компоненты микробиоты, такие как жгутики и липополисахарид эндотоксина (ЛПС), передают сигнал толл-подобным рецепторам (TLR).
Рецепторы, присутствующие на клетках EE, позволяют этим клеткам ощущать просветные и, возможно, циркулирующие SCFA, что запускает высвобождение нескольких метаболически активных гормонов кишечника. Сигнал SCFA через два преобладающих механизма: (1) ингибирование ядерной гистоновой деацетилазы (HDAC) (Waldecker et al., 2008; Fellows et al., 2018; Larraufie et al., 2018) для изменения транскрипции и экспрессии гена и (2 ) стимуляция рецепторов 2 и 3 свободных жирных кислот, связанных с G-белком (FFAR3, FFAR3), экспрессируемых по всей длине желудочно-кишечного тракта в различных региональных структурах.FFAR2 имеет одинаковое сродство к ацетату, пропионату и бутирату, в то время как сродство FFAR3 к ацетату значительно ниже, чем к пропионату и бутирату (Offermanns, 2014). Таким образом, передача сигналов SCFA через FFAR2 / 3 зависит от комбинации типа рецептора и количества конкретных метаболитов.
Вторичные желчные кислоты
Желчные кислоты представляют собой амфипатические молекулы, синтезируемые гепатоцитами из холестерина и высвобождаемые в просвет желудочно-кишечного тракта, чтобы способствовать солюбилизации и, следовательно, абсорбции пищевых липидов.Давно известно, что кишечная микробиота напрямую участвует в метаболизме желчных кислот хозяина, поскольку эффективная энтерогепатическая рециркуляция солей желчных кислот в значительной степени зависит от деконъюгации и дигидроксилирования желчных кислот гидролазами солей желчных кислот микробного происхождения (BSH). Это приводит к образованию вторичных желчных кислот (Jones et al., 2008), которые являются более гидрофобными и, таким образом, могут реабсорбироваться посредством пассивной диффузии, ограничивая потерю желчных кислот с фекалиями. Специфическая активность BSH в кишечной микробиоте человека различается между разными типами: BSH у Firmicutes и Actinobacteria способна метаболизировать все конъюгированные соли желчных кислот, а активность BSH Bacteroidetes специфична для тауро-конъюгированных желчных кислот (Jones et al., 2008). В дополнение к этой роли в желудочно-кишечном тракте желчные кислоты являются сигнальными молекулами, которые участвуют в периферическом метаболизме. Желчные кислоты играют важную роль в периферическом метаболизме благодаря своему действию на два рецептора желчных кислот, рецептор, связанный с G-белком TGR5 (ранее известный как Gpbar1), и ядерный рецептор FXR, оба из которых экспрессируются в клетках EE (рис. 2). Сродство к рецепторам и их эффективность существенно различаются между разными желчными кислотами. Таким образом, микробиом кишечника может оказывать сильное влияние на метаболизм хозяина, изменяя состав пула желчных кислот за счет изменения синтеза и повторного поглощения желчных кислот.
Клеточное распознавание структурных компонентов микробов
Структурные компоненты микробной мембраны, такие как жгутики и мембраносвязанный липополисахарид (LPS), действуют как сигнальные молекулы через ряд белков распознавания клеточных образов (Gordon, 2002). ЛПС представляет собой компонент клеточной стенки грамотрицательных бактерий, таких как представители типа Bacteroidetes, и является мощным лигандом для толл-подобных рецепторов (TLR), особенно толл-подобных рецепторов 4 (TLR4). Помимо мощных эффектов, вызывающих иммунитет и воспаление (Takeuchi and Akira, 2002; Lancaster et al., 2018), экспрессия TLR была продемонстрирована в ряде клеток EE, и активация этого рецептора запускает секрецию ряда метаболически активных гормонов, таких как GLP-1 (Lebrun et al., 2017), 5-HT (Kidd et al., 2009) и PYY (Larraufie et al., 2017; рисунок 2). Клинически повышенные уровни ЛПС (эндотоксемия) тесно связаны с ожирением и инсулинорезистентностью (Cani et al., 2007a). Механизмы, с помощью которых LPS вносят вклад в нарушенный гликемический контроль и ожирение, вероятно, включают сложные взаимодействия между секрецией кишечных гормонов, целостностью слизистого барьера и воспалением хозяина и иммунными путями.
Микробиом регулирует метаболизм хозяина с помощью высвобождения гормона кишечника
Серотонин
Энтерохромаффинные (ЕС) клетки являются источником почти всего (около 95%) серотонина (5-HT; 5-гидрокситриптамина) в организме. Эти клетки составляют почти половину всех клеток EE и рассредоточены по всей длине желудочно-кишечного тракта с различной плотностью (Raghupathi et al., 2013). Давно известно, что ЕС-клетки играют важную роль во многих внутренних механизмах кишечника, связанных с моторикой (Keating and Spencer, 2010; Spencer et al., 2011, 2015; Спенсер и Китинг, 2016; Keating and Spencer, 2018), а клетки ЭК способны ощущать свою локальную питательную среду и реагировать, выделяя 5-HT уникальным образом (Zelkas et al., 2015; Raghupathi et al., 2016; Thorn et al., 2016) . В настоящее время есть твердые доказательства того, что 5-HT, полученный из кишечника, является ключевым фактором нарушения регуляции периферического метаболизма (Sumara et al., 2012; Watanabe et al., 2014; Crane et al., 2015; Young et al., 2015, 2018; Martin et al., 2017c). Отсутствие 5-HT, полученного из кишечника, за счет фармакологического ингибирования или генетического подавления фермента, ограничивающего скорость синтеза 5-HT в кишечнике, триптофангидроксилазы 1 (TPh2), обеспечивает защиту мышей от ожирения, вызванного диетой (Crane et al. al., 2015). Более того, циркулирующий 5-HT увеличивается у людей с ожирением и положительно коррелирует с индексом массы тела (Young et al., 2018) и плохим гликемическим контролем (Takahashi et al., 2002).
Микробиом кишечника влияет на уровни 5-HT в организме хозяина. У мышей, получавших GF и антибиотики, уровень 5-HT, полученного из клеток EC, существенно ниже по сравнению с контрольными животными с обычным повышенным уровнем (CONV-R), который восстанавливается путем колонизации мышей GF микробиотой кишечника доноров (Yano et al., 2015) . Клетки ЭК обладают способностью воспринимать микробные метаболиты, поскольку они экспрессируют как FFAR2, так и FFAR3 (Akiba et al., 2015; Martin et al., 2017b) и ряда обонятельных рецепторов (Bellono et al., 2017; Lund et al., 2018). Однако острое воздействие SCFA на первичные клетки ЕС мыши в культуре не вызывает увеличения секреции 5-HT (Martin et al., 2017a). Скорее, увеличение 5-HT, наблюдаемое в присутствии микробиома кишечника (Reigstad et al., 2015; Yano et al., 2015), вероятно, связано с хроническим воздействием, опосредующим увеличение биосинтеза 5-HT, способствовало за счет увеличения пролиферации клеток EC (Yano et al., 2015). Кроме того, инфузия люминального бутирата восстанавливает перистальтику кишечника у мышей GF, которая притупляется у мышей TPh2-KO, указывая на то, что эффекты бутирата могут опосредоваться клетками 5-HT EC (Vincent et al., 2018). Острые реакции на ароматические метаболиты, такие как изобутират и изовалерат, наблюдались в клетках ЭК в составе кишечных органоидов (Bellono et al., 2017), вероятно, через активацию обонятельных рецепторов. Однако вполне вероятно, что это непрямой ответ из-за перекрестного взаимодействия с другими гормонами кишечного происхождения, такими как GLP-1, которые также увеличиваются после воздействия микробных метаболитов и обладают способностью передавать сигналы клеткам EC (Lund и другие., 2018).
ГПП-1
Глюкагоноподобный пептид 1 (GLP-1), продукт расщепления проглюкагона, секретируется L-клетками, преимущественно расположенными в подвздошной и толстой кишке. GLP-1 представляет собой инкретиновый гормон (Kreymann et al., 1987), высвобождаемый после приема пищи и в ответ на питательные вещества, такие как глюкоза (Sun et al., 2017), для увеличения инсулина и ингибирования секреции глюкагона поджелудочной железой (Grondahl et al., 2017). Кроме того, GLP-1 ингибирует опорожнение желудка и влияет на сытость и прием пищи (Holst, 2007).Считается, что вместе с PYY, GLP-1 лежит в основе некоторых метаболических улучшений, наблюдаемых после операции обходного желудочного анастомоза (Madsbad and Holst, 2014), а действие GLP-1 лежит в основе некоторых глюкозоснижающих способностей терапии диабета, метформина ( Bahne et al., 2018). Таким образом, терапевтические средства, нацеленные на GLP-1, включая аналоги GLP-1 и ингибиторы деградации GLP-1 под действием фермента дипептидилпептидазы IV (DPP-4), широко используются из-за их антидиабетических свойств (Aroda et al., 2012) .Интересно, что микробиота обладает активностью, подобной DPP-4 (Olivares et al., 2018b), которая у мышей снижается при введении ингибитора DPP-4, вилдаглиптина, и сопровождается изменением микробного состава, которое не зависит от прямые эффекты ингибирования DPP-4 на функцию микробиоты (Olivares et al., 2018a). В частности, лечение вилдаглиптином связано с уменьшением численности Oscillibacter и увеличением численности Lactobacillus , с уменьшением количества лигандов TLR и увеличением пропионата (Olivares et al., 2018а). Таким образом, DPP-4-подобная активность кишечных бактерий может потенциально влиять на уровни циркулирующих GLP-1 и PYY, которые, в свою очередь, могут существовать как петля обратной связи, чтобы влиять на микробный состав и изобилие микробных метаболитов.
Между L-клетками и микробиомом кишечника существует динамическая взаимосвязь. Опосредованная желчной кислотой активация TGR5 (Bala et al., 2014), передачи сигналов SCFA (Tolhurst et al., 2012), LPS и других метаболитов, таких как индол (Chimerel et al., 2014), являются мощными стимуляторами секреции GLP-1. .Регуляция GLP-1 вторичными желчными кислотами зависит от задействованного рецепторного сигнального пути, поскольку активация TGR5 увеличивает секрецию GLP-1, тогда как, с другой стороны, активация FXR снижает секрецию GLP-1. Эта динамическая взаимосвязь еще более усложняется перекрестным взаимодействием TGR5-FXR, которое существует между этими двумя рецепторами, особенно в толстой кишке (Pathak et al., 2017). Инфузия желчной кислоты хенодезоксихолевой кислоты в просвет крыс запускает высвобождение GLP-1 в сосудистую сеть, помимо PYY, через TGR5 (Kuhre et al., 2018). Индол, еще один важный бактериальный метаболит, полученный из пищевого триптофана, резко стимулирует GLP-1, увеличивая продолжительность потенциала клеточного действия (Chimerel et al., 2014). Напротив, хроническое воздействие индола дозозависимо снижает секрецию GLP-1 в первичных L-клетках мыши путем ингибирования пути синтеза АТФ (Chimerel et al., 2014). Кроме того, микробный ЛПС запускает секрецию GLP-1 через TLR4 после повреждения слизистого барьера (Lebrun et al., 2017) и, как таковой, стимулированную глюкозой секрецию инсулина на мышиной модели эндотоксемии (Nguyen et al., 2014). Диетические пребиотики, такие как олигофруктоза, которые увеличивают продукцию SCFA бактериями, связаны с усилением дифференцировки L-клеток и содержанием GLP-1 в проксимальном отделе толстой кишки крыс и снижают прибавку в весе при введении до и во время HFD (Cani et al., 2007b) . Увеличение количества L-клеток, посредством чего увеличивается высвобождение GLP-1 после приема пищи, также связано с усилением чувства насыщения и снижением ожирения (Cani et al., 2007b). Механизмы, с помощью которых SCFA увеличивают секрецию GLP-1, зависят от региона, так как передача сигналов в тонком кишечнике осуществляется преимущественно через FFAR3, тогда как опосредованное FFAR2 высвобождение GLP-1 происходит в толстой кишке (Greiner and Backhed, 2016).
Парадоксально, но у мышей, получавших GF и антибиотики, были более высокие уровни циркулирующего GLP-1 во время голодания (Zarrinpar et al., 2018) и пониженное содержание GLP-1 в слизистой оболочке (Duca et al., 2012) по сравнению с генетически идентичными мышами CONV-R . В недавнем исследовании (Arora et al., 2018) сообщается, что профиль экспрессии генов L-клеток подвздошной кишки, полученных от мышей GF, существенно отличается от мышей CONV-R. Примечательно, что многие из генов, регулирующих функциональную способность L-клеток, активируются у мышей GF, а L-клетки имеют большее количество секреторных пузырьков у мышей GF.Что лежит в основе этих различий у мышей GF, неизвестно, но может быть отражением устойчивости к GLP-1, которая наблюдается при ожирении, вызванном диетой, и связана с измененным составом микробиоты, особенно в подвздошной кишке (Grasset et al., 2017).
PYY
Пептид тирозин-тирозин (PYY) синтезируется и секретируется L-клетками, помимо GLP-1, и преимущественно экспрессируется в нижних отделах тонкой кишки и толстой кишки. PYY регулирует потребление пищи и насыщение посредством активации центральных рецепторов Y2, связанных с G-белком, на нейропептиде Y (NPY) и нейронах AgRP в аркуатном ядре гипоталамуса (Dumont et al., 1995). Это инициирует сигнальный каскад, посредством которого стимулирующие аппетит нейроны NPY подавляются, обеспечивая растормаживание индуцирующего насыщение пути проопиомеланокортина (POMC) / α-MSH (Loh et al., 2015). У людей с ожирением наблюдается снижение циркулирующего PYY (Batterham et al., 2006) в результате ослабленной секреции PYY в толстой кишке (le Roux et al., 2006), а не устойчивости к PYY (Batterham et al., 2003). Циркулирующий PYY существует в двух формах: PYY 1-36 и DPP-4-расщепленный PYY 3-36 , причем последняя является наиболее доминирующей циркулирующей формой после приема пищи (Grandt et al., 1994) и наиболее биологически сильнодействующим в отношении аноректических эффектов (Chelikani et al., 2006).
Таким образом, способность кишечной микробиоты влиять на секрецию PYY имеет важное значение для развития ожирения и нарушений обмена веществ. Микробные SCFA, особенно бутират, вызывают зависящее от дозы и времени увеличение экспрессии гена PYY в двух модельных клеточных линиях EE и в первичных культурах клеток толстой кишки человека (Larraufie et al., 2018). Кроме того, пероральный прием бутирата умеренно увеличивает циркулирующий PYY (Lin et al., 2012). Механизмы, с помощью которых SCFA увеличивают биосинтез PYY, по-видимому, связаны с комбинацией передачи сигналов FFAR2 / 3 всеми SCFA и ингибирования HCAD пропионатом и бутиратом (Larraufie et al., 2018). Хотя эти механизмы кажутся видоспецифичными (Larraufie et al., 2018) и не сопровождались повышением секреции GLP-1, которое наблюдается после воздействия пропионата на первичные культуры толстой кишки мышей (Psichas et al., 2015) . Использование мыши с нокаутом FFAR2 демонстрирует участие передачи сигналов SCFA в увеличении количества PYY-содержащих клеток, особенно у мышей, получавших диету, богатую предшественником SCFA, инулином (Brooks et al., 2017). Изменение микробиоты кишечника человека в результате 4-дневного режима приема антибиотиков широкого спектра действия резко и обратимо увеличило постпрандиальный плазменный PYY (Mikkelsen et al., 2015). Однако точные изменения микробных метаболитов и видов бактерий, лежащие в основе этого изменения, неизвестны. Вторичные желчные кислоты также являются мощными стимулами для секреции PYY, и механизмы, с помощью которых это происходит, согласуются с механизмами секреции GLP-1 (Kuhre et al., 2018). Люминальная перфузия смеси первичных и вторичных желчных кислот в просвет крысы с сосудистой перфузией увеличивает уровни PYY в венозном эффлюенте TGR5-зависимым образом, в то время как такой же эффект наблюдался при инфузии только CDCA вторичной желчной кислоты (Kuhre et al., 2018).
GIP
Глюкозозависимый инсулинотропный пептид (GIP), также известный как желудочный ингибирующий пептид, представляет собой инкретиновый гормон, высвобождаемый после приема пищи в тонком кишечнике из классически определенных К-клеток (Buffa et al., 1975). Активность GIP передается через рецепторы GIP (GIPR), экспрессируемые в β-клетках поджелудочной железы (Gremlich et al., 1995), адипоцитах (Yip et al., 1998), костных клетках (Bollag et al., 2000) и в нейроны ЦНС (Paratore et al., 2011). Подобно GLP-1, биологическая активность GIP быстро ослабляется ферментативным расщеплением DPP-IV (Baggio and Drucker, 2007).Внутри поджелудочной железы GIP вносит значительный вклад в постпрандиальную секрецию инсулина посредством увеличения биосинтеза инсулина (Baggio and Drucker, 2007) и усиления пролиферации β-клеток (Widenmaier et al., 2009). Считается, что дефектная передача сигналов GIP лежит в основе, по крайней мере частично, ослабленной глюкозо-стимулированной секреции инсулина, наблюдаемой у пациентов с T2D (Vilsboll et al., 2002). GIP также широко считается адипогенным гормоном (Thondam et al., 2017), поскольку он способствует захвату и хранению липидов в адипоцитах (Getty-Kaushik et al., 2006).
Повышенный уровень GIP был связан с наблюдаемым повышенным ожирением, вызванным субтерапевтическим режимом приема антибиотиков у мышей при отъеме в течение 7 недель, поскольку лечение не влияло на уровни других гормонов кишечника в плазме (Cho et al., 2012). Лечение значительно увеличило соотношение Firmicutes / Bacteroidetes и уровни SCFA в слепой кишке, что потенциально могло лежать в основе повышенного уровня GIP и, следовательно, повышенного ожирения. Однако недавние противоречивые данные демонстрируют, что углеводы в просвете ингибируют секрецию GIP посредством микробного сигнального пути SCFA-FFAR3 (Lee et al., 2018). Неизвестно, связано ли расхождение, наблюдаемое в этих исследованиях, с дифференциальной передачей сигналов через FFAR2 и FFAR3. В соответствии со специфическими рецепторными путями для SCFA-опосредованного высвобождения гормона кишечника, пероральное введение бутирата натрия мышам, как было показано, временно увеличивает секрецию GIP и GLP-1, в то время как пируват натрия и коктейль SCFA являются селективными для увеличения GIP, но не GLP- 1 или PYY (Lin et al., 2012).
CCK
Холецистокинин (CCK) происходит из классически названных «I-клеток», преимущественно локализованных в верхних отделах тонкой кишки (Dockray, 2012).CCK высвобождается в ответ на потребление жиров и белков с пищей. CCK играет четко определенную роль в регуляции аппетита (Ritter, 2004; Becskei et al., 2007), опорожнении желудка и моторике (Raybould and Tache, 1988; Raybould, 1991; Ellis et al., 2013) и высвобождении желчных кислот. и ферменты поджелудочной железы, которые важны для пищеварения (Li and Owyang, 1994; Li and Owyang, 1996; Owyang and Logsdon, 2004), посредством активации рецепторов CCK (Rogers and Hermann, 2008). О регуляции ХЦК кишечными микробами известно меньше по сравнению с другими гормонами кишечника, в основном из-за воздействия на ХЦК-содержащие клетки микробиоты, ограниченной тонким кишечником.У свиней инфузия в подвздошную кишку ацетата, пропионата и бутирата SCFAs во время кормления повышала уровни CCK в плазме и парадоксальным образом подавляла секрецию поджелудочной железы (Sileikiene et al., 2008). Однако были проведены ограниченные исследования микробной регуляции ХЦК у людей. В одном отчете от пациентов с обходным желудочным анастомозом Roux-en-Y не было выявлено никаких изменений в уровнях циркулирующих ХЦК у людей с нормальным весом и ожирением до и после операции, несмотря на значительное изменение микробного состава после операции (Federico et al., 2016). Однако сниженная экспрессия белка CCK наблюдается в диссоциированных клетках проксимального отдела тонкого кишечника мышей GF, что не было связано с уменьшением количества клеток EE (Duca et al., 2012).
Диета влияет на состав кишечной микробиоты
Разнообразие кишечной микробиоты и относительное количество микробных метаболитов (метаболомный профиль) в значительной степени зависит от конкретных диетических компонентов (David et al., 2014), о чем свидетельствует существенная разница в микробных сообществах с потреблением богатых растениями или белков. богатые диеты.Это происходит из-за селективного давления, оказываемого питательными веществами на микробиоту, в пользу видов бактерий, обогащенных генами, необходимыми для метаболизма определенных субстратов. Например, растительная диета и потребление пробиотиков увеличивает содержание волокон в просвете и сложных углеводов, в результате чего выбираются виды, обогащенные углеводно-активными ферментами (David et al., 2014). Диеты на основе животных, богатые жирами и белками и с низким содержанием клетчатки, увеличивают содержание желчных кислот в просвете, способствуя развитию устойчивых к желчным кислотам микробов, обогащенных генами метаболизма желчных кислот, такими как гидрозазы желчных кислот и сульфитредуктаза (Devkota et al., 2012; Дэвид и др., 2014). Пищевые волокна также оказывают большое влияние на транзит через кишечник (подробно рассмотрено Muller et al., 2018), что, в свою очередь, является важным фактором, определяющим состав и метаболизм фекального микробиома (Roager et al., 2016; Vandeputte et al. , 2016). Однако в отсутствие пищевых волокон наблюдается компенсаторный сдвиг в микробиоме кишечника с увеличением популяций, экспрессирующих ферменты, расщепляющие муцин, что указывает на общее предпочтение микробов субстратам на основе волокон (Desai et al., 2016).
Повышенное потребление непитательных подсластителей (NNS), таких как сахарин, сукралоза и аспартам, хотя и очень полезно для снижения потребления калорий и отклонений уровня глюкозы в крови, также имеет долгосрочные последствия для состава микробиома и непереносимости глюкозы. Примечательно, что потребление обычных NNS у мышей усугубляет развитие непереносимости глюкозы (Suez et al., 2014, 2015) и увеличение веса у самцов (Bian et al., 2017), что опосредуется различными функциональными изменениями в организме. микробиом кишечника (Suez et al., 2014). В частности, сахарин NNS и ацесульфам-K увеличивают численность представителей Bacteroidetes (Suez et al., 2014; Bian et al., 2017) и снижают численность Firmicutes (Suez et al., 2014). Влияние NNS на метаболизм и микробный состав у людей в значительной степени продиктовано природным микробным составом до воздействия NNS (Suez et al., 2014), в то время как у мышей реакция, по-видимому, также зависит от пола (Bian et al. , 2017). Тем не менее, изменения микробного состава, вызванные NNS, наблюдаемые в этих исследованиях, согласуются с микробным составом, наблюдаемым при ожирении и метаболических заболеваниях (Ley et al., 2006; Тернбо и Гордон, 2009; Turnbaugh et al., 2009).
Богатство и разнообразие микробиома кишечника человека коррелируют с метаболической функцией (Le Chatelier et al., 2013; David et al., 2014; Dao et al., 2016), и был установлен основной микробиом, вызывающий ожирение, который характеризуется высоким отношение Firmicutes к Bacteroidetes (Ley et al., 2006; Turnbaugh, Gordon, 2009; Turnbaugh et al., 2009). Изменение состава микробиома для уменьшения численности Bacteroidetes, как видно при краткосрочном ограничении калорийности, приводит к улучшению метаболических исходов у мышей в результате снижения продукции LPS и снижения передачи сигналов TLR4 (Fabbiano et al., 2018). Существуют также крупномасштабные обсервационные исследования, которые связывают использование антибиотиков с риском развития диабета 2 типа (Boursi et al., 2015; Mikkelsen et al., 2015), хотя при интерпретации этих результатов следует избегать осторожности, поскольку гипергликемия является фактором риска. на инфекцию (Falagas and Kompoti, 2006) и, таким образом, может служить основанием для увеличения использования антибиотиков. Кроме того, специфические клинически используемые антибиотики, в частности ванкомицин-имипенем и ципрофлоксацин, вызывают различное воздействие на состав микробиома и количество микробных метаболитов после возобновления роста (Choo et al., 2017), что может иметь последствия для специфического воздействия антибиотиков на метаболизм. Проглатывание ксенобиотиков, таких как фармацевтические препараты и химические вещества, относящиеся к окружающей среде, может изменить микробный состав кишечника с последующими последствиями для метаболизма. Об этом свидетельствует лекарство от диабета метформин, терапевтический эффект которого частично зависит от изменения микробного состава кишечника (Wu et al., 2017). Метаболизм ксенобиотиков микробиотой кишечника также химически различен (Koppel et al., 2017), в котором микробиом кишечника рассматривается как возможный инструмент для целенаправленной разработки и доставки лекарств. Недавняя работа Vangay et al. (2018) элегантно показали, что миграция из незападной культуры в западную культуру быстро и межпоколенческая влияет на разнообразие кишечной микробиоты. Эта потеря микробной сложности и биоразнообразия привела к потере ключевых микробных ферментов, необходимых для переваривания растительных волокон, что частично объясняется изменением состава рациона и уменьшением разнообразия пищи, и может предрасполагать людей к метаболическим заболеваниям (Vangay et al., 2018). Таким образом, вмешательства по изменению состава микробиоты кишечника могут быть мощным терапевтическим инструментом для лечения ожирения и метаболических нарушений.
Авторские взносы
Все перечисленные авторы внесли существенный, прямой и интеллектуальный вклад в работу и одобрили ее к публикации.
Финансирование
DK был поддержан исследовательской стипендией Национального совета по здравоохранению и медицинским исследованиям (NHMRC) Австралии.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Редактор обработки объявил о прошлом сотрудничестве с одним из авторов DK.
Список литературы
Aguirre, M., Eck, A., Koenen, M. E., Savelkoul, P.H., Budding, A.E., and Venema, K. (2016). Диета приводит к быстрым изменениям метаболической активности и состава кишечной микробиоты человека на проверенной модели кишечника in vitro. Res. Microbiol. 167, 114–125. DOI: 10.1016 / j.resmic.2015.09.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Акиба, Ю., Иноуэ, Т., Кадзи, И., Хигасияма, М., Наримацу, К., Ивамото, К. и др. (2015). Определение короткоцепочечных жирных кислот в двенадцатиперстной кишке крысы. J. Physiol. 593, 585–599. DOI: 10.1113 / jphysiol.2014.280792
CrossRef Полный текст | Google Scholar
Арода, В. Р., Генри, Р. Р., Хан, Дж., Хуанг, В., Де Янг, М. Б., Дарсо, Т. и др. (2012). Эффективность агонистов рецептора GLP-1 и ингибиторов DPP-4: метаанализ и систематический обзор. Clin. Ther. 34, 1247.e22–1258.e22. DOI: 10.1016 / j.clinthera.2012.04.013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Арора, Т., Акрами, Р., Пайс, Р., Бергквист, Л., Йоханссон, Б. Р., Шварц, Т. В. и др. (2018). Микробная регуляция транскриптома L-клеток. Sci. Отчет 8: 1207. DOI: 10.1038 / s41598-017-18079-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Bahne, E., Sun, E. W. L., Young, R. L., Hansen, M., Sonne, D. P., Hansen, J. S., и другие. (2018). Вызванная метформином секреция глюкагоноподобного пептида-1 способствует действию метформина при диабете 2 типа. JCI Insight 3: 93936. DOI: 10.1172 / jci.insight.93936
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бала В., Раджагопал С., Кумар Д. П., Налли А. Д., Махавади С., Саньял А. Дж. И др. (2014). Высвобождение GLP-1 и PYY в ответ на активацию связанного с G-белком рецептора желчных кислот TGR5 опосредуется путем Epac / PLC-эпсилон и модулируется эндогенным h3S. Фронт. Physiol. 5: 420. DOI: 10.1007 / s00125-018-4647-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Баттерхэм, Р. Л., Коэн, М. А., Эллис, С. М., Ле Ру, К. В., Уизерс, Д. Дж., Фрост, Г. С. и др. (2003). Подавление приема пищи у пациентов с ожирением пептидом YY3-36. N. Engl. J. Med. 349, 941–948. DOI: 10.1056 / NEJMoa030204
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Баттерхэм, Р. Л., Хеффрон, Х., Капур, С., Чиверс, Дж. Э., Чандарана, К., Херцог, Х. и др. (2006). Критическая роль пептида YY в опосредованном белком насыщении и регуляции массы тела. Cell Metab. 4, 223–233. DOI: 10.1016 / j.cmet.2006.08.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беккей, К., Граблер, В., Эдвардс, Г. Л., Ридигер, Т., и Лутц, Т. А. (2007). Поражение латерального парабрахиального ядра ослабляет аноректический эффект периферического амилина и ХЦК. Brain Res. 1162, 76–84. DOI: 10.1016 / j.brainres.2007.06.016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беллоно, Н. В., Байрер, Дж. Р., Лейтч, Д. Б., Кастро, Дж., Чжан, К., О’Доннелл, Т. А. и др. (2017). Энтерохромаффинные клетки — это хемосенсоры кишечника, которые связаны с сенсорными нервными путями. Ячейка 170, 185.e16–198.e16. DOI: 10.1016 / j.cell.2017.05.034
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Биан, Х., Чи, Л., Гао, Б., Ту, П., Ру, Х. и Лу, К. (2017). Искусственный подсластитель ацесульфам калий влияет на микробиом кишечника и прирост массы тела мышей CD-1. PLoS One 12: e0178426. DOI: 10.1371 / journal.pone.0178426
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Боллаг, Р. Дж., Чжун, К., Филлипс, П., Мин, Л., Чжун, Л., Камерон, Р. и др. (2000). Клетки, происходящие из остеобластов, экспрессируют функциональные глюкозозависимые рецепторы инсулинотропных пептидов. Эндокринология 141, 1228–1235.DOI: 10.1210 / endo.141.3.7366
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Brooks, L., Viardot, A., Tsakmaki, A., Stolarczyk, E., Howard, J. K., Cani, P. D., et al. (2017). Ферментируемый углевод стимулирует FFAR2-зависимую экспансию PYY-клеток толстой кишки, повышая чувство насыщения. Мол. Метаб. 6, 48–60. DOI: 10.1016 / j.molmet.2016.10.011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Буффа, Р., Полак, Дж. М., Пирс, А.Г., Сольча, Э., Гримелиус, Л., и Капелла, К. (1975). Идентификация кишечных клеток, хранящих желудочный ингибирующий пептид. Гистохимия 43, 249–255. DOI: 10.1007 / BF00499706
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Cani, P. D., Amar, J., Iglesias, M. A., Poggi, M., Knauf, C., Bastelica, D., et al. (2007a). Метаболическая эндотоксемия вызывает ожирение и инсулинорезистентность. Diabetes Metab. Res. Ред. 56, 1761–1772.
Google Scholar
Кани, П.Д., Хост, С., Гио, Ю., Дельзенн, Н. М. (2007b). Пищевые неперевариваемые углеводы способствуют дифференцировке L-клеток в проксимальном отделе толстой кишки крыс. руб. J. Nutr. 98, 32–37. DOI: 10.1017 / S0007114507691648
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Челикани, П. К., Хейвер, А. К., Рив, Дж. Р. мл., Кейре, Д. А., и Рейдельбергер, Р. Д. (2006). Ежедневная периодическая внутривенная инфузия пептида YY (3-36) снижает ежедневное потребление пищи и ожирение у крыс. Am. J. Physiol. Regul. Интегр. Комп. Physiol. 290, R298 – R305. DOI: 10.1152 / ajpregu.00674.2005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чимерель, К., Эмери, Э., Саммерс, Д. К., Кейзер, У., Гриббл, Ф. М., и Рейманн, Ф. (2014). Бактериальный метаболит индол модулирует секрецию инкретина кишечными энтероэндокринными L-клетками. Cell Rep. 9, 1202–1208. DOI: 10.1016 / j.celrep.2014.10.032
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чо, И., Яманиши, С., Кокс, Л., Мете, Б.А., Завадил, Дж., Ли, К. и др. (2012). Антибиотики в раннем возрасте изменяют микробиом толстой кишки и ожирение мыши. Природа 488, 621–626. DOI: 10.1038 / природа11400
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чу, Дж. М., Канно, Т., Зайн, Н. М., Леонг, Л. Е., Абелл, Г. К., Кибл, Дж. Е. и др. (2017). Дивергентные отношения между фекальной микробиотой и метаболомом после явных нарушений, вызванных антибиотиками. мСфера 2: e00005-17.
PubMed Аннотация | Google Scholar
Крейн, Дж. Д., Паланивел, Р., Моттилло, Э. П., Буяк, А. Л., Ван, Х., Форд, Р. Дж. И др. (2015). Подавление периферического синтеза серотонина снижает ожирение и метаболическую дисфункцию, способствуя термогенезу коричневой жировой ткани. Nat. Med. 21, 166–172. DOI: 10,1038 / нм.3766
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Куч, Г., и Мальберт, К. Х.(1999). Короткоцепочечные жирные кислоты, присутствующие в подвздошной кишке, подавляют моторику желудочно-кишечного тракта натощак у находящихся в сознании свиней. Нейрогастроэнтерол. Мотил. 11, 219–225. DOI: 10.1046 / j.1365-2982.1999.00149.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Каммингс, Дж. Х., Помаре, Э. У., Бранч, У. Дж., Нейлор, К. П., и Макфарлейн, Г. Т. (1987). Короткоцепочечные жирные кислоты в толстой кишке, воротной, печеночной и венозной крови человека. Кишечник 28, 1221–1227. DOI: 10.1136 / гут.28.10.1221
CrossRef Полный текст | Google Scholar
Дао, М. К., Эверард, А., Арон-Вишневски, Дж., Соколовска, Н., Прифти, Э., Вергер, Э. О. и др. (2016). Akkermansia muciniphila и улучшение метаболического здоровья во время диетического вмешательства при ожирении: взаимосвязь с богатством микробиома кишечника и экологией. Кишечник 65, 426–436. DOI: 10.1136 / gutjnl-2014-308778
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дэвид, Л.А., Морис, К.Ф., Кармоди, Р. Н., Гутенберг, Д. Б., Баттон, Дж. Э., Вулф, Б. Е. и др. (2014). Диета быстро и воспроизводимо изменяет микробиом кишечника человека. Природа 505, 559–563. DOI: 10.1038 / природа12820
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Десаи, М.С., Сикатц, А.М., Коропаткин, Н.М., Камада, Н., Хики, К.А., Уолтер, М. и др. (2016). Микробиота кишечника, лишенная пищевых волокон, разрушает слизистый барьер толстой кишки и повышает восприимчивость к патогенам. Ячейка 167, 1339.e21–1353.e21. DOI: 10.1016 / j.cell.2016.10.043
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Девкота, С., Ван, Ю., Муш, М. В., Леоне, В., Фелнер-Пич, Х., Надимпалли, А., и др. (2012). Таурохолевая кислота, индуцированная пищевым жиром, способствует размножению патобионтов и колиту у мышей Il10 — / -. Природа 487, 104–108. DOI: 10.1038 / природа11225
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Duca, F.А., Шварц, Т. Д., Сакар, Ю., Коваса, М. (2012). Повышенное оральное обнаружение, но снижение кишечной сигнализации жиров у мышей с недостаточной кишечной микробиотой. PLoS One 7: e39748. DOI: 10.1371 / journal.pone.0039748
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дюмон Ю., Фурнье А., Сен-Пьер С. и Квирион Р. (1995). Характеристика сайтов связывания нейропептида Y в препаратах мембран головного мозга крысы с использованием [125I] [Leu31, Pro34] пептида YY и [125I] пептида YY3-36 в качестве селективных радиолигандов Y1 и Y2. J. Pharmacol. Exp. Ther. 272, 673–680.
Google Scholar
Эллис, М., Чемберс, Дж. Д., Гвинн, Р. М., и Борнштейн, Дж. К. (2013). Серотонин и холецистокинин опосредуют индуцированную питательными веществами сегментацию в тонкой кишке морской свинки. Am. J. Physiol. Гастроинтест. Liver Physiol. 304, G749 – G761. DOI: 10.1152 / ajpgi.00358.2012
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фаббиано, С., Суарес-Заморано, Н., Шевалье, К., Лазаревич В., Кизер С., Риго Д. и др. (2018). Ремоделирование функциональной микробиоты кишечника способствует метаболическим улучшениям, вызванным ограничением калорийности. Cell Metab. 28, 907.e7–921.e7. DOI: 10.1016 / j.cmet.2018.08.005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фалагас, М. Э. и Компоти, М. (2006). Ожирение и инфекция. Lancet Infect. Dis. 6, 438–446. DOI: 10.1016 / S1473-3099 (06) 70523-0
CrossRef Полный текст | Google Scholar
Федерико, А., Даллио, М., Толоне, С., Гравина, А.Г., Патроне, В., Романо, М. и др. (2016). Желудочно-кишечные гормоны, кишечная микробиота и метаболический гомеостаз у пациентов с ожирением: эффект бариатрической хирургии. In Vivo 30, 321–330.
PubMed Аннотация | Google Scholar
Fellows, R., Denizot, J., Stellato, C., Cuomo, A., Jain, P., Stoyanova, E., et al. (2018). Полученные из микробиоты короткоцепочечные жирные кислоты способствуют кротонилированию гистонов в толстой кишке через гистоновые деацетилазы. Nat. Commun. 9: 105. DOI: 10.1038 / s41467-017-02651-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фотергилл, Л. Дж., Каллаган, Б., Хунн, Б., Браво, Д. М., и Фернесс, Дж. Б. (2017). Совместное хранение энтероэндокринных гормонов оценивается на клеточном и субклеточном уровнях у самцов мышей. Эндокринология 158, 2113–2123. DOI: 10.1210 / en.2017-00243
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гетти-Кошик, Л., Сонг, Д. Х., Бойлан, М. О., Корки, Б. Э. и Вулф, М. М. (2006). Глюкозозависимый инсулинотропный полипептид модулирует липолиз и реэтерификацию адипоцитов. Ожирение 14, 1124–1131. DOI: 10.1038 / oby.2006.129
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гордон, С. (2002). Рецепторы распознавания образов: удвоение для врожденного иммунного ответа. Ячейка 111, 927–930. DOI: 10.1016 / S0092-8674 (02) 01201-1
CrossRef Полный текст | Google Scholar
Грандт, Д., Schimiczek, M., Beglinger, C., Layer, P., Goebell, H., Eysselein, V.E., et al. (1994). Две молекулярные формы пептида YY (PYY) широко распространены в крови человека: характеристика радиоиммуноанализа, распознающего PYY 1-36 и PYY 3-36. Регул. Pept. 51, 151–159. DOI: 10.1016 / 0167-0115 (94)
-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Grasset, E., Puel, A., Charpentier, J., Collet, X., Christensen, J.E., Terce, F., et al. (2017). Специфический дисбиоз кишечной микробиоты мышей с диабетом 2 типа индуцирует резистентность к GLP-1 через энтеральный NO-зависимый механизм и механизм оси кишечник-мозг. Cell Metab. 25, 1075.e5–1090.e5. DOI: 10.1016 / j.cmet.2017.06.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гремлих, С., Поррет, А., Хани, Э. Х., Шериф, Д., Вионнет, Н., Фрогель, П. и др. (1995). Клонирование, функциональная экспрессия и хромосомная локализация рецептора глюкозозависимого инсулинотропного полипептида островков поджелудочной железы человека. Diabetes Metab. Res. Ред. 44, 1202–1208.
PubMed Аннотация | Google Scholar
Грондаль, М.Ф., Китинг Д. Дж., Вилсболл Т. и Кноп Ф. К. (2017). Современные методы лечения, которые изменяют секрецию глюкагона: каков терапевтический эффект таких модификаций? Curr. Диабетический отчет 17: 128. DOI: 10.1007 / s11892-017-0967-z
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гу, С., Чен, Д., Чжан, Дж. Н., Львов, X., Ван, К., Дуань, Л. П. и др. (2013). Картирование бактериального сообщества желудочно-кишечного тракта мышей. PLoS One 8: e74957. DOI: 10.1371 / journal.pone.0074957
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хартстра А. В., Боутер К. Э., Бэкхед Ф. и Ньивдорп М. (2015). Понимание роли микробиома в ожирении и диабете 2 типа. Уход за диабетом 38, 159–165. DOI: 10.2337 / dc14-0769
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джонс, Б. В., Бегли, М., Хилл, К., Гахан, К. Г. и Марчези, Дж. Р. (2008). Функциональный и сравнительный метагеномный анализ активности гидролаз желчных солей в микробиоме кишечника человека. Proc. Natl. Акад. Sci. USA 105, 13580–13585. DOI: 10.1073 / pnas.0804437105
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Китинг, Д. Дж., И Спенсер, Н. Дж. (2010). Высвобождение 5-гидрокситриптамина из слизистой оболочки не требуется для образования или распространения двигательных комплексов, мигрирующих по толстой кишке. Гастроэнтерология 138, 659–670, 670.e1-2. DOI: 10.1053 / j.gastro.2009.09.020
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Китинг, Д.Дж. И Спенсер Н. Дж. (2018). Какова роль эндогенного серотонина кишечника в контроле моторики желудочно-кишечного тракта? Pharmacol. Res. 140, 50–55. DOI: 10.1016 / j.phrs.2018.06.017
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кидд М., Густафссон Б. И., Дроздов И. и Модлин И. М. (2009). Секреция серотонина, индуцированная IL1бета и ЛПС, повышена в клетках ЭК, полученных из болезни Крона. Нейрогастроэнтерол. Мотил. 21, 439–450.DOI: 10.1111 / j.1365-2982.2008.01210.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кох А., Де Ваддер Ф., Ковачева-Датчари П. и Бакхед Ф. (2016). От пищевых волокон к физиологии хозяина: короткоцепочечные жирные кислоты как ключевые бактериальные метаболиты. Cell 165, 1332–1345. DOI: 10.1016 / j.cell.2016.05.041
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Коскинен, К., Паусан, М. Р., Перрас, А. К., Бек, М., Банг, К., Мора М. и др. (2017). Первые сведения о разнообразных археомах человека: специфическое обнаружение архей в желудочно-кишечном тракте, легких, носу и на коже. мBio 8, e824 – e817. DOI: 10.1128 / mBio.00824-17
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Крейманн Б., Уильямс Г., Гатеи М.А. и Блум С.Р. (1987). Глюкагоноподобный пептид-1 7-36: физиологический инкретин в человеке. Ланцет 2, 1300–1304. DOI: 10.1016 / S0140-6736 (87)
-9
CrossRef Полный текст | Google Scholar
Кухре, Р.E., Wewer Albrechtsen, N.J., Larsen, O., Jepsen, S.L., Balk-Moller, E., Andersen, D. B., et al. (2018). Желчные кислоты являются важными прямыми и косвенными регуляторами секреции гормонов, регулирующих аппетит и метаболизм, из кишечника и поджелудочной железы. Мол. Метаб. 11, 84–95. DOI: 10.1016 / j.molmet.2018.03.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Laforest-Lapointe, I., and Arrieta, M.C. (2018). Микробные эукариоты: недостающее звено в исследованиях микробиома кишечника. mSystems 3: e00201-17.
Google Scholar
Лам, Ю. Ю., Ха, К. В., Кэмпбелл, К. Р., Митчелл, А. Дж., Динудом, А., Оскарссон, Дж. И др. (2012). Повышенная проницаемость кишечника и изменение микробиоты связаны с воспалением мезентериального жира и метаболической дисфункцией у мышей с ожирением, вызванным диетой. PLoS One 7: e34233. DOI: 10.1371 / journal.pone.0034233
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ланкастер, Г. И., Лэнгли, К.Г., Берглунд, Н. А., Каммун, Х. Л., Рейбе, С., Эстевес, Э. и др. (2018). Доказательства того, что TLR4 не является рецептором для насыщенных жирных кислот, а опосредует индуцированное липидами воспаление, перепрограммируя метаболизм макрофагов. Cell Metab. 27, 1096.e5–1110.e5. DOI: 10.1016 / j.cmet.2018.03.014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ларроуфи П., Дор Дж., Лапак Н. и Блоттьер Х. М. (2017). Лиганды TLR и бутират увеличивают экспрессию Pyy двумя различными, но взаимно регулируемыми путями. Cell Microbiol. 19: e12648. DOI: 10,1111 / cmi.12648
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Larraufie, P., Martin-Gallausiaux, C., Lapaque, N., Dore, J., Gribble, F.M., Reimann, F., et al. (2018). SCFAs сильно стимулируют продукцию PYY в энтероэндокринных клетках человека. Sci. Реп. 8:74. DOI: 10.1038 / s41598-017-18259-0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ле Шателье, Э., Нильсен, Т., Qin, J., Prifti, E., Hildebrand, F., Falony, G., et al. (2013). Богатство микробиома кишечника человека коррелирует с метаболическими маркерами. Природа 500, 541–546. DOI: 10.1038 / nature12506
PubMed Аннотация | CrossRef Полный текст | Google Scholar
le Roux, C. W., Batterham, R. L., Aylwin, S.J., Patterson, M., Borg, C.M., Wynne, K. J., et al. (2006). Ослабленное высвобождение пептида YY у субъектов с ожирением связано со снижением насыщения. Эндокринология 147, 3–8.DOI: 10.1210 / en.2005-0972
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Lebrun, L.J., Lenaerts, K., Kiers, D., Pais, de Barros, J.P., Le Guern, N., et al. (2017). Энтероэндокринные L-клетки ощущают ЛПС после повреждения кишечного барьера, чтобы усилить секрецию ГПП-1. Cell Rep. 21, 1160–1168. DOI: 10.1016 / j.celrep.2017.10.008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ли, Э. Ю., Чжан, X., Миямото, Дж., Кимура, И., Такнака, Т., Furusawa, K., et al. (2018). Углеводы кишечника подавляют секрецию GIP через путь микробиоты / SCFA / FFAR3. J. Endocrinol. 239, 267–276. DOI: 10.1530 / JOE-18-0241
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ли Ю. и Оуян К. (1994). Эндогенный холецистокинин стимулирует секрецию ферментов поджелудочной железы через блуждающий афферентный путь у крыс. Гастроэнтерология 107, 525–531. DOI: 10.1016 / 0016-5085 (94)
-5
CrossRef Полный текст | Google Scholar
Ли, Ю., и Owyang, C. (1996). Секреция поджелудочной железы, вызванная холецистокинином и холецистокинин-зависимыми дуоденальными стимулами через афферентные волокна блуждающего нерва у крысы. J. Physiol. 494 (Pt 3), 73–82. DOI: 10.1113 / jphysiol.1996.sp021531
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лин, Х. В., Фрассетто, А., Ковалик, Э. Дж. Мл., Навроцки, А. Р., Лу, М. М. и др. (2012). Бутират и пропионат защищают от ожирения, вызванного диетой, и регулируют гормоны кишечника посредством независимых от рецептора 3 свободных жирных кислот механизмов. PLoS One 7: e35240. DOI: 10.1371 / journal.pone.0035240
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Lund, M. L., Egerod, K. L., Engelstoft, M. S., Dmytriyeva, O., Theodorsson, E., Patel, B.A., et al. (2018). Энтерохромаффинные 5-HT клетки — основная мишень для GLP-1 и метаболитов кишечных микробов. Мол. Метаб. 11, 70–83. DOI: 10.1016 / j.molmet.2018.03.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мадсбад, С.и Холст Дж. Дж. (2014). GLP-1 как медиатор в ремиссии диабета 2 типа после желудочного обходного анастомоза и рукавной гастрэктомии. Diabetes Metab. Res. Ред. 63, 3172–3174. DOI: 10.2337 / db14-0935
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мартин А. М., Ламсден А. Л., Янг Р. Л., Джессап К. Ф., Спенсер Н. Дж. И Китинг Д. Дж. (2017a). Региональные различия в секреции серотонина кишечника, вызванной питательными веществами. Physiol. Реп. 5: e13199.DOI: 10.14814 / phy2.13199
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мартин А. М., Ламсден А. Л., Янг Р. Л., Джессап К. Ф., Спенсер Н. Дж. И Китинг Д. Дж. (2017b). Репертуар чувствительных к питательным веществам энтерохромаффинных клеток мышей различается для двенадцатиперстной и толстой кишки. Нейрогастроэнтерол. Мотил. 29: e13046. DOI: 10.1111 / nmo.13046
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мартин, А.М., Янг, Р.Л., Леонг, Л., Роджерс, Г. Б., Спенсер, Н. Дж., Джессап, К. Ф. и др. (2017c). Разнообразные метаболические роли периферического серотонина. Эндокринология 158, 1049–1063. DOI: 10.1210 / en.2016-1839
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Миккельсен, К. Х., Фрост, М., Баль, М. И., Лихт, Т. Р., Йенсен, США, Розенберг, Дж. И др. (2015). Влияние антибиотиков на микробиоту кишечника, гормоны кишечника и метаболизм глюкозы. PLoS One 10: e0142352. DOI: 10,1371 / журнал.pone.0142352
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Молинаро А., Цезарь Р., Холм Л. М., Тремароли В., Кани П. Д. и Бакхед Ф. (2017). Взаимодействие хозяина и микробиоты вызывает двухфазное воспаление и непереносимость глюкозы у мышей. Мол. Метаб. 6, 1371–1380. DOI: 10.1016 / j.molmet.2017.08.016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мюллер М., Канфора Э. Э. и Блаак Э. Э. (2018). Время прохождения через желудочно-кишечный тракт, гомеостаз глюкозы и метаболическое здоровье: модуляция с помощью пищевых волокон. Питательные вещества 10: 275. DOI: 10.3390 / nu10030275
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Nguyen, A. T., Mandard, S., Dray, C., Deckert, V., Valet, P., Besnard, P., et al. (2014). Опосредованное липополисахаридами увеличение секреции инсулина, стимулированной глюкозой: участие пути GLP-1. Diabetes Metab. Res. Ред. 63, 471–482. DOI: 10.2337 / db13-0903
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ньивдорп, М., Гилийамсе, П. В., Пай, Н., и Каплан, Л. М. (2014). Роль микробиома в регуляции энергии и метаболизма. Гастроэнтерология 146, 1525–1533. DOI: 10.1053 / j.gastro.2014.02.008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Оливарес М., Нейринк А. М., Потгенс С. А., Бомонт М., Салазар Н., Кани П. Д. и др. (2018a). Ингибитор ДПП-4 вилдаглиптин воздействует на микробиоту кишечника и предотвращает нарушение гомеостаза кишечника, вызванного западной диетой у мышей. Diabetologia 61, 1838–1848. DOI: 10.1007 / s00125-018-4647-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Оливарес, М., Шуппель, В., Хассан, А. М., Бомонт, М., Нейринк, А. М., Биндельс, Л. Б. и др. (2018b). Потенциальная роль дипептидилпептидазы-4-подобной активности кишечной микробиоты на здоровье хозяина. Фронт. Microbiol. 9: 1900. DOI: 10.3389 / fmicb.2018.03313
CrossRef Полный текст | Google Scholar
Овьянг, К., и Логсдон, К. Д. (2004). Новые взгляды на нейрогормональную регуляцию секреции поджелудочной железы. Гастроэнтерология 127, 957–969. DOI: 10.1053 / j.gastro.2004.05.002
CrossRef Полный текст | Google Scholar
Параторе, С., Чиотти, М. Т., Базиль, М., Водри, Д., Джентиле, А., Паренти, Р. и др. (2011). Желудочный ингибирующий полипептид и его рецептор экспрессируются в центральной нервной системе и поддерживают выживание нейронов. Cent. Nerv. Syst. Agents Med. Chem. 11, 210–222. DOI: 10.2174 / 187152411798047771
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Патак П., Лю Х., Беме С., Се К., Краус К. В., Гонсалес Ф. и др. (2017). Рецептор Фарнезоида X индуцирует перекрестное взаимодействие рецептора 5 G-белка Takeda для регулирования синтеза желчных кислот и метаболизма в печени. J. Biol. Chem. 292, 11055–11069. DOI: 10.1074 / jbc.M117.784322
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Педерсен, Х.K., Gudmundsdottir, V., Nielsen, H.B., Hyotylainen, T., Nielsen, T., Jensen, B.A., et al. (2016). Микробы кишечника человека влияют на метаболом сыворотки хозяина и чувствительность к инсулину. Природа 535, 376–381. DOI: 10.1038 / природа18646
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Психас А., Слит М. Л., Мерфи К. Г., Брукс Л., Бьюик Г. А., Ханьялоглу А. С. и др. (2015). Пропионат короткоцепочечных жирных кислот стимулирует секрецию GLP-1 и PYY через рецептор 2 свободных жирных кислот у грызунов. Внутр. J. Obes. 39, 424–429. DOI: 10.1038 / ijo.2014.153
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рагхупати Р., Даффилд М. Д., Зелкас Л., Мидения А., Брукс С. Дж., Сиа Т. С. и др. (2013). Идентификация уникальной кинетики высвобождения серотонина из энтерохромаффинных клеток морской свинки и человека. J. Physiol. 591, 5959–5975. DOI: 10.1113 / jphysiol.2013.259796
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рагхупати, Р., Джессап, К. Ф., Ламсден, А. Л., и Китинг, Д. Дж. (2016). Размер пор слияния ограничивает высвобождение 5-HT из везикул отдельных энтерохромафинных клеток. J. Cell. Physiol. 231, 1593–1600. DOI: 10.1002 / jcp.25256
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рейбоулд, Х. Э. (1991). Чувствительные к капсаицину афференты блуждающего нерва и CCK в ингибировании моторной функции желудка, вызванной питательными веществами кишечника. Пептиды 12,1279–1283. DOI: 10.1016 / 0196-9781 (91)
-6PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рейбоулд, Х.Э., и Таш Ю. (1988). Холецистокинин подавляет перистальтику желудка и опорожнение через капсаицин-чувствительный вагусный путь у крыс. Am. J. Physiol. 255, G242 – G246. DOI: 10.1152 / ajpgi.1988.255.2.G242
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рейгстад, К. С., Салмонсон, К. Э., Рейни, Дж. Ф. III, Шуршевски, Дж. Х., Линден, Д. Р., Зонненбург, Дж. Л. и др. (2015). Микробы кишечника способствуют выработке серотонина в толстой кишке за счет воздействия короткоцепочечных жирных кислот на энтерохромаффинные клетки. FASEB J. 29, 1395–1403. DOI: 10.1096 / fj.14-259598
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ридаура, В. К., Фейт, Дж. Дж., Рей, Ф. Э., Ченг, Дж., Дункан, А. Е., Кау, А. Л. и др. (2013). Микробиота кишечника близнецов, не согласных с ожирением, модулирует метаболизм у мышей. Наука 341, 1241214. DOI: 10.1126 / science.1241214
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Роагер, Х. М., Хансен, Л. Б. С., Bahl, M. I., Frandsen, H. L., Carvalho, V., Gøbel, R. J., et al. (2016). Время прохождения через толстую кишку связано с метаболизмом бактерий и обновлением слизистой оболочки кишечника. Nat. Microbiol. 1: 16093. DOI: 10.1038 / nmicrobiol.2016.93
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Родригес, Р. Р., Грир, Р. Л., Донг, X., DSouza, К. Н., Гурунг, М., Ву, Дж. Ю. и др. (2017). Вызванные антибиотиками изменения в микробиоте кишечника связаны с изменениями метаболизма глюкозы у здоровых мышей. Фронт. Microbiol. 8: 2306. DOI: 10.3389 / fmicb.2017.02306
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Роджерс, Г. Б., Китинг, Д. Дж., Янг, Р. Л., Вонг, М. Л., Лисинио, Дж., И Весселинг, С. (2016). От дисбактериоза кишечника до нарушения функции мозга и психических заболеваний: механизмы и пути. Мол. Психиатрия 21, 738–748. DOI: 10.1038 / mp.2016.50
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Скарпеллини, Э., Яниро, Г., Аттили, Ф., Бассанелли, К., Де Сантис, А., и Гасбаррини, А. (2015). Микробиота кишечника человека и виром: потенциальные терапевтические последствия. Dig. Liver Dis. 47, 1007–1012. DOI: 10.1016 / j.dld.2015.07.008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сили Р. Дж., Чемберс А. П. и Сандовал Д. А. (2015). Роль адаптации кишечника в мощном влиянии нескольких бариатрических операций на ожирение и диабет. Cell Metab. 21, 369–378. DOI: 10.1016 / j.cmet.2015.01.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Силейкиене В., Мозентин Р., Бауэр Э., Пиефо Х. П., Тафай М., Крушевска Д. и др. (2008). Влияние подвздошной инфузии короткоцепочечных жирных кислот на прандиальную секрецию поджелудочной железы и желудочно-кишечные гормоны у свиней. Поджелудочная железа 37, 196–202. DOI: 10.1097 / MPA.0b013e31816386f4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Спенсер, Н.Дж. И Китинг Д. Дж. (2016). Есть ли роль эндогенного 5-HT в моторике желудочно-кишечного тракта? как недавние исследования изменили наше понимание. Adv. Exp. Med. Биол. 891, 113–122. DOI: 10.1007 / 978-3-319-27592-5_11
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Спенсер, Н. Дж., Николас, С. Дж., Робинсон, Л., Кайло, М., Флэк, Н., Брукс, С. Дж. И др. (2011). Механизмы, лежащие в основе перистальтики дистального отдела толстой кишки морских свинок, вызванной растяжением: роль энтерохромаффинных клеток? Am.J. Physiol . Gastrointestinal Liver Physiol. 301, G519 – G527. DOI: 10.1152 / ajpgi.00101.2011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Спенсер, Н. Дж., Сиа, Т. К., Брукс, С. Дж., Коста, М., и Китинг, Д. Дж. (2015). Противоположная точка зрения CrossTalk: 5-HT не нужен для перистальтики. J. Physiol. 593, 3229–3231. DOI: 10.1113 / JP270183
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Суарес-Заморано, Н., Fabbiano, S., Chevalier, C., Stojanovic, O., Colin, D. J., Stevanovic, A., et al. (2015). Истощение микробиоты способствует потемнению белой жировой ткани и снижает ожирение. Nat. Med. 21, 1497–1501. DOI: 10,1038 / нм. 3994
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Suez, J., Korem, T., Zeevi, D., Zilberman-Schapira, G., Thaiss, C.A., Maza, O., et al. (2014). Искусственные подсластители вызывают непереносимость глюкозы, изменяя микробиоту кишечника. Природа 514, 181–186.DOI: 10.1038 / природа13793
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Суэц, Дж., Корем, Т., Зильберман-Шапира, Г., Сегал, Э. и Элинав, Э. (2015). Некалорийные искусственные подсластители и микробиом: выводы и проблемы. Кишечные микробы 6, 149–155. DOI: 10.1080 / 194
.2015.1017700
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сумара, Г., Сумара, О., Ким, Дж. К., и Карсенти, Г. (2012). Серотонин кишечного происхождения является многофункциональным детерминантом адаптации к голоданию. Cell Metab. 16, 588–600. DOI: 10.1016 / j.cmet.2012.09.014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Sun, E. W., de Fontgalland, D., Rabbitt, P., Hollington, P., Sposato, L., Due, S. L., et al. (2017). Механизмы контроля индуцированной глюкозой секреции GLP-1 в тонком кишечнике человека. Diabetes Metab. Res. Ред. 66, 2144–2149. DOI: 10.2337 / db17-0058
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Такахаши, Т., Яно, М., Минами, Дж., Харагути, Т., Кога, Н., Хигаши, К. и др. (2002). Сарпогрелат гидрохлорид, антагонист рецепторов серотонина 2А, снижает альбуминурию у больных диабетом с ранней стадией диабетической нефропатии. Diabetes Res. Clin. Практик. 58, 123–129. DOI: 10.1016 / S0168-8227 (02) 00105-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тондам, С. К., Дауси, К., Уилдинг, Дж. П., Холст, Дж. Дж., Амин, Г. И., Янг, К. и др. (2017). Глюкозозависимый инсулинотропный полипептид способствует отложению липидов в подкожных адипоцитах у больных диабетом 2 типа с ожирением: неадаптивный ответ. Am. J. Physiol. Эндокринол. Метаб. 312, E224 – E233. DOI: 10.1152 / ajpendo.00347.2016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Tolhurst, G., Heffron, H., Lam, Y. S., Parker, H.E., Habib, A.M., Diakogiannaki, E., et al. (2012). Короткоцепочечные жирные кислоты стимулируют секрецию глюкагоноподобного пептида-1 через рецептор FFAR2, связанный с G-белком. Diabetes Metab. Res. Rev. 61,364–371. DOI: 10.2337 / db11-1019
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Топпинг, Д.Л. и Клифтон П. М. (2001). Короткоцепочечные жирные кислоты и функция толстой кишки человека: роль резистентных крахмальных и некрахмальных полисахаридов. Physiol. Ред. 81, 1031–1064. DOI: 10.1152 / Physrev.2001.81.3.1031
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тернбо, П. Дж., И Гордон, Дж. И. (2009). Микробиом кишечника, энергетический баланс и ожирение. J. Physiol. 587, 4153–4158. DOI: 10.1113 / jphysiol.2009.174136
CrossRef Полный текст | Google Scholar
Тернбо, П.Дж., Хамади, М., Яцуненко, Т., Кантарел, Б. Л., Дункан, А., Лей, Р. Э. и др. (2009). Основной микробиом кишечника у тучных и худых близнецов. Природа 457, 480–484. DOI: 10.1038 / nature07540
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тернбо, П. Дж., Лей, Р. Э., Маховальд, М. А., Магрини, В., Мардис, Е. Р., и Гордон, Дж. И. (2006). Микробиом кишечника, связанный с ожирением, с повышенной способностью собирать энергию. Nature 444, 1027–1031. DOI: 10.1038 / природа05414
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вандепутте, Д., Фалони, Г., Виейра-Силва, С., Тито, Р. Ю., Йоосенс, М., и Раес, Дж. (2016). Консистенция стула тесно связана с богатством и составом кишечной микробиоты, энтеротипами и скоростью роста бактерий. Кишечник 65, 57–62. DOI: 10.1136 / gutjnl-2015-309618
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вангай П., Джонсон А. Дж., Уорд Т.Л., Аль-Галит, Г. А., Шилдс-Катлер, Р. Р., Хиллманн, Б. М. и др. (2018). Иммиграция в США вестернизирует микробиом кишечника человека. Ячейка 175, 962.e10–972.e10. DOI: 10.1016 / j.cell.2018.10.029
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вилсболл, Т., Краруп, Т., Мадсбад, С., и Холст, Дж. Дж. (2002). Дефектное усиление поздней фазы инсулинового ответа на глюкозу с помощью GIP у пациентов с диабетом II типа с ожирением. Диабетология 45, 1111–1119.DOI: 10.1007 / s00125-002-0878-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Винсент, А. Д., Ван, X. Y., Парсонс, С. П., Хан, В. И., и Хейзинга, Дж. Д. (2018). Аномальная абсорбционная моторная активность толстой кишки у свободных от микробов мышей устраняется бутиратом, эффект, возможно, опосредованный серотонином слизистой оболочки. Am. J. Physiol. Гастроинтест. Liver Physiol. 315, G896 – G907. DOI: 10.1152 / ajpgi.00237.2017
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вризе, А., Out, C., Fuentes, S., Jonker, L., Reuling, I., Kootte, R. S., et al. (2014). Влияние перорального ванкомицина на микробиоту кишечника, метаболизм желчных кислот и чувствительность к инсулину. J. Hepatol. 60, 824–831. DOI: 10.1016 / j.jhep.2013.11.034
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вризе, А., Ван Нуд, Э., Холлеман, Ф., Салоярви, Дж., Кутте, Р. С., Бартельсман, Дж. Ф. и др. (2012). Перенос кишечной микробиоты от худых доноров увеличивает чувствительность к инсулину у людей с метаболическим синдромом. Гастроэнтерология 143, 913.e7–916.e7. DOI: 10.1053 / j.gastro.2012.06.031
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вальдекер, М., Каутенбургер, Т., Дауман, Х., Буш, К., и Шренк, Д. (2008). Подавление активности гистон-деацетилазы короткоцепочечными жирными кислотами и некоторыми метаболитами полифенолов, образующимися в толстой кишке. J. Nutr. Biochem. 19, 587–593. DOI: 10.1016 / j.jnutbio.2007.08.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ватанабэ, Х., Сайто, Р., Накано, Т., Такахаши, Х., Такахаши, Ю., Сумиёси, К. и др. (2014). Влияние периферического 5-HT на метаболизм глюкозы и липидов у овец. PLoS One 9: e88058. DOI: 10.1371 / journal.pone.0088058
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Виденмайер, С. Б., Ао, З., Ким, С. Дж., Уорнок, Г., и Макинтош, К. Х. (2009). Подавление p38 MAPK и JNK посредством Akt-опосредованного ингибирования киназы 1, регулирующей сигнал апоптоза, составляет основной компонент эффектов выживания бета-клеток глюкозозависимого инсулинотропного полипептида. J. Biol. Chem. 284, 30372–30382. DOI: 10.1074 / jbc.M109.060178
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wu, H., Esteve, E., Tremaroli, V., Khan, M. T., Caesar, R., Mannerås-Holm, L., et al. (2017). Метформин изменяет микробиом кишечника людей с диабетом 2 типа, ранее не получавшим лечения, что способствует терапевтическому эффекту препарата. Nat. Med. 22, 850–858. DOI: 10,1038 / нм. 4345
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Яно, Дж.М., Ю., К., Дональдсон, Г. П., Шастри, Г. Г., Энн, П., Ма, Л. и др. (2015). Аборигенные бактерии микробиоты кишечника регулируют биосинтез серотонина хозяина. Cell 161, 264–276. DOI: 10.1016 / j.cell.2015.02.047
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Янг Р. Л., Ламсден А. Л., Мартин А. М., Шобер Г., Пезос Н., Тажат С. С. и др. (2018). Повышенная способность к высвобождению периферического серотонина при ожирении у человека. Внутр. J. Obes. 42, 1880–1889. DOI: 10.1038 / s41366-018-0047-8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Зарринпар А., Чаикс А., Сюй З. З., Чанг, М. В., Мароц, К. А., Сагателян, А. и др. (2018). Вызванное антибиотиками истощение микробиома изменяет метаболический гомеостаз, влияя на передачу сигналов в кишечнике и метаболизм в толстой кишке. Nat. Commun. 9: 2872. DOI: 10.1038 / s41467-018-05336-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Зелкас, Л., Рагхупати, Р., Ламсден, А. Л., Мартин, А. М., Сан, Э., Спенсер, Н. Дж. И др. (2015). Энтероэндокринные клетки, секретирующие серотонин, через различные механизмы реагируют на острые и хронические изменения доступности глюкозы. Nutr. Метаб. 12:55. DOI: 10.1186 / s12986-015-0051-0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
.