ОСОБЕННОСТИ МЕТАБОЛИЗМА ОРГАНИЗМА ХОККЕИСТОВ ВЫСШЕЙ КВАЛИФИКАЦИИ В ХОДЕ СОРЕВНОВАТЕЛЬНОГО ПЕРИОДА | КОЛЕСОВ
1. Rohleder N., Beulen S.E., Chen E., Wolf J. M., Kirschbaum C. Stress on the dance floor: the cortisol stress response to socialevaluative threat in competitive ballroom dancers. Personality and Social Psychology Bulletin. 2007; 33(1): 69-84. DOI: 10.1177/014616720629386.
2. Карузин К.А., Бойцов В.И., Мартусевич А.К., Левушкин С.П. Влияние комплексной программы оптимизации метаболизма на работоспособность спортсменов. Кубанский научный медицинский весник. 2016; 6: 80-88. [Karuzin K.A., Boytsov V.I., Martusevich A.K., Levushkin S.P. Vliyanie kompleksnoy programmy optimizatsii metabolizma na rabotosposobnost’ sportsmenov. Kubanskiy nauchnyy meditsinskiy vesnik. 2016; 6: 80-88 (In Russ.)].
3. Siart B., Nimmerichter A., Vidotto C., Wallner B. Status, Stress and Performance in Track and Field Athletes during the European Games in Baku (Azerbaijan). Sci. rep. 2017; 7(1): 6076. Doi: 10.1038/s41598-017-06461-z.
4. Колесов С.А., Рахманов Р.С., Блинова Т.В., Страхова Л.А., Чумаков Н.В. Сывороточный оксид азота и адаптация к физическим нагрузкам на фоне приема продукта спортивного питания. Медицина труда и экология человека. 2017; 1: 8492. [Kolesov S.A., Rakhmanov R.S., Blinova T.V., Strakhova L.A., Chumakov N.V. Serum nitric oxide and adaptation to physical loads during dietary administration of sport nutrition product. Meditsina truda i ekologiya cheloveka. 2017; 1: 84-92 (In Russ.)].
5. Muhsin H., Aynur O., İlhan O., Mehmet S., Ozan S. Effect of Increasing Maximal Aerobic Exercise on Serum Muscles Enzymes in Professional Field Hockey Players.
6. Афанасьева И.А., Таймазов В.А. Синдром перетренированности у спортсменов: эндогенная интоксикация и факторы врожденного иммунитета. Ученые записки университета им. П.Ф. Лесгафта. 2011; 82(12): 24-30. [Afanas’eva I.A., Taymazov V.A. Sindrom peretrenirovannosti u sportsmenov: endogennaya intoksikatsiya i faktory vrozhdennogo immuniteta. Uchenye zapiski universiteta im. P.F. Lesgafta. 2011; 82(12): 24-30 (In Russ.)].
7. Ипатенко О.Н. Биохимический мониторинг работоспособности футболиста в тренировочных и игровых циклах. Журнал практической спортивной медицины. URL: https://sportdoktor.ru/ genthemes/genthemes_712.html. [Ipatenko O.N. Biokhimicheskiy monitoring rabotosposobnosti futbolista v trenirovochnykh i igrovykh tsiklakh. Zhurnal prakticheskoy sportivnoy meditsiny. URL: https://sportdoktor.ru/genthemes/genthemes_712.html (In Russ.)].
8. Куранов А.А., Балеев М.С., Митрофанова Н.Н., Мельников В.Л. Некоторые аспекты патогенеза атеросклероза и факторы риска развития сердечно-сосудистых заболеваний. Фундаментальные исследования. 2014; 10-6: 12341238. [Kuranov A.A., Baleev M.S., Mitrofanova N.N., Mel’nikov V.L. Nekotorye aspekty patogeneza ateroskleroza i faktory riska razvitiya serdechno-sosudistykh zabolevaniy. Fundamental’nye issledovaniya. 2014; 10-6:1234-1238 (In Russ.)].
9. Nasseu J.R., Ama Moor V.J., Takam R.D.M., Zing-Awona B., Azabji-Kenfack M., Tankeu F., Tchoula C.M., Moukette B.M., Ngogang J.Y. Cameroonian professional soccer players and risk of atherosclerosis. BMC Res Notes. 2017; 10(1): 186. Doi: 10.1186/s13104-017-2508-x.
10. Скальный А.В., Рудаков И.А. Биоэлементы в медицине. М.: Издательский дом «ОНИКС 21 век»: Мир. 2004; 272 с. [Skal’nyy A.V., Rudakov I.A. Bioelementy v meditsine. M.: Izdatel’skiy dom «ONIKS 21 vek»: Mir. 2004; 272 s (In Russ.)].
Скальный А.В., Рудаков И.А. Биоэлементы в медицине. М.: Издательский дом «ОНИКС 21 век»: Мир. 2004; 272 с. [Skal’nyy A.V., Rudakov I.A. Bioelementy v meditsine. M.: Izdatel’skiy dom «ONIKS 21 vek»: Mir. 2004; 272 s (In Russ.)].
11. Долгов В.В., Шевченко О.П. Лабораторная диагностика нарушений обмена белков. М.: ECO-MED-POLL. 1997; 68 с. [Dolgov V.V., Shevchenko O.P. Laboratornaya diagnostika narusheniy obmena belkov. M.: ECO-MED-POLL; 1997. 68 s (In Russ.)].
12. Baird M.F., Graham S.M., Baker J.S., Bickerstaff G.F. Creatine-Kinase- and Exercise-Related Muscle Damage Implications for Muscle Performance and Recovery. J Nutr Metab. 2012; 2012: 96036. Published online. Doi: 10.1155/2012/960363.
13. Brancaccio P., Maffulli N., Limongelli F.M. Creatine kinase monitoring in sport medicine. Br Med Bull. 2007; 81-82: 209-230.
14. Wadenaar F., Brinkmans N., Ceelen I., Van Rooij B., Mensink M., Witkamp R., De Vries J. Micronutrient Intakes in 553 Dutch Elite and Sub – Elite Athletes: Prevalence of Low and High Intakes in Users and Non – Users of Nutritional Supplements. Nutrients. 2017; 9(2): 142. Doi: 10.3390/nu9020142.
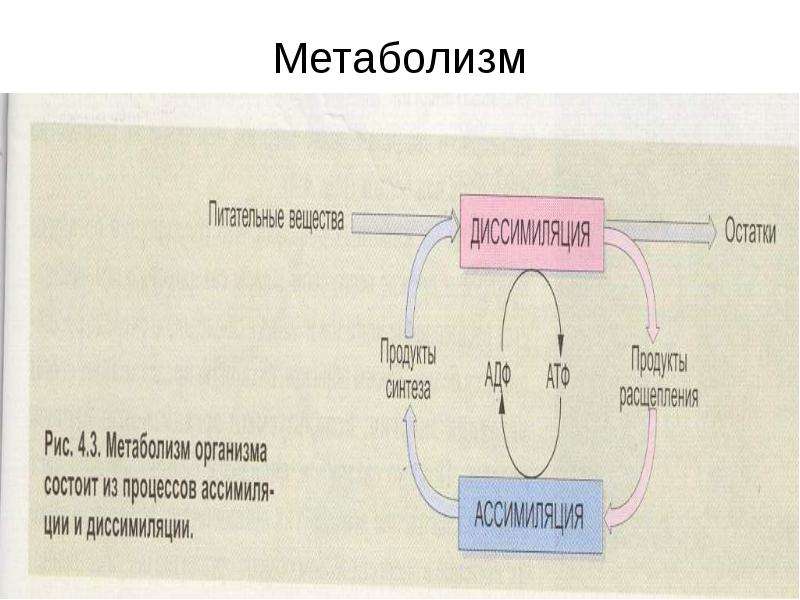
Возрастные особенности метаболизма и клеточного энергообмена у женщин | Пестряева Л.А., Третьякова Т.Б., Дерябина Е.Г., Данькова И.В., Кинжалова С.В., Борисова С.В.
Цель исследования: выявить особенности метаболизма и клеточного энергообмена у женщин в перименопаузе.Материал и методы: обследованы 2 группы женщин: 1-я (контрольная) — 25 женщин репродуктивного возраста;
2-я (основная) — 25 женщин в перименопаузе.
 На момент обследования все пациентки контрольной группы характеризовали менструальный цикл как регулярный. Менопауза у пациенток основной группы — естественная, наступила после 45 лет, длительность — от 1 года до 3-х лет. У 64% пациенток основной группы наблюдались заболевания, обусловленные нарушением нейрогуморальной регуляции: дискинезия желудочно-кишечного тракта и желчевыводящих путей, нейроциркуляторная дистония по гипертоническому типу, избыточная масса тела и ожирение (индекс массы тела >25 кг/м2). На момент обследования обострения хронических экстрагенитальных заболеваний не отмечалось. В основной группе климактерический синдром выявлен у 56% женщин, в т. ч. средней и тяжелой степени. Биохимическое исследование сыворотки венозной крови выполнено на автоматическом анализаторе с помощью унифицированных тест-систем. Активность ферментов клеточного энергообмена сукцинатдегидрогеназы и a-глицерофосфатдегидрогеназы в лимфоцитах периферической крови определяли цитохимическим методом с последующей визуальной морфометрией.
На момент обследования все пациентки контрольной группы характеризовали менструальный цикл как регулярный. Менопауза у пациенток основной группы — естественная, наступила после 45 лет, длительность — от 1 года до 3-х лет. У 64% пациенток основной группы наблюдались заболевания, обусловленные нарушением нейрогуморальной регуляции: дискинезия желудочно-кишечного тракта и желчевыводящих путей, нейроциркуляторная дистония по гипертоническому типу, избыточная масса тела и ожирение (индекс массы тела >25 кг/м2). На момент обследования обострения хронических экстрагенитальных заболеваний не отмечалось. В основной группе климактерический синдром выявлен у 56% женщин, в т. ч. средней и тяжелой степени. Биохимическое исследование сыворотки венозной крови выполнено на автоматическом анализаторе с помощью унифицированных тест-систем. Активность ферментов клеточного энергообмена сукцинатдегидрогеназы и a-глицерофосфатдегидрогеназы в лимфоцитах периферической крови определяли цитохимическим методом с последующей визуальной морфометрией.Выводы: у женщин в перименопаузе активность митохондриальных ферментов ниже, чем в репродуктивном возрасте. В период перименопаузы наблюдается снижение клеточного энергообмена и синтеза половых гормонов, о чем косвенно свидетельствует повышение содержания основных метаболитов (общего белка, альбумина, глюкозы, холестерина, липопротеидов низкой плотности, триглицеридов, мочевой кислоты). Изучение процессов внутриклеточного энергообмена на различных этапах онтогенеза является перспективным направлением научного поиска, крайне важного для сохранения репродуктивного потенциала и профилактики заболеваний старости на популяционном уровне. Ключевые слова: перименопауза, менопауза, метаболизм, энергообмен, сукцинатдегидрогеназа, a-глицерофосфатдегидрогеназа.

Для цитирования: Пестряева Л.А., Третьякова Т.Б., Дерябина Е.Г. и др. Возрастные особенности метаболизма и клеточного энергообмена у женщин. РМЖ. Мать и дитя. 2018;26(5(I)):16-18.
Age features of metabolism and cellular energy exchange in womenPestryaeva L.A., Tretyakova T.B., Deryabina E.G., Dankova I.V., Kinzhalova S.V., Borisova S.V.
Urals Research Institute of Maternity and Childhood, Ekaterinburg
Aim: identification of the features of metabolism and cellular energy exchange in perimenopausal women.
Results: age specific features of metabolism and cell bioenergetics (decrease in the activity of energy metabolism enzymes, an increase in the total protein, albumin, glucose, cholesterol, triglycerides, uric acid) in perimenopausal women were established.

Conclusions: women in perimenopause have a lower activity of mitochondrial enzymes than women in reproductive age. During the period of menopause there is a decrease in cellular energy exchange and synthesis of sex hormones, as evidenced by the increase in the content of the main metabolites (total protein, albumin, glucose, cholesterol, LDL, triglycerides, uric acid). The study of processes of intracellular energy exchange at various stages of ontogenesis is a promising direction of scientific research, it is extremely important to maintain the reproductive potential and prevent old age diseases at the population level.
Key words: perimenopause, menopause, metabolism, energy exchange, succinate dehydrogenase, a-glitserofosfatdegidrogenase.
For citation: Pestryaeva L.A., Tretyakova T.B., Deryabina E.G. et al. Age features of metabolism and cellular energy exchange in women // RMJ. 2018. № 5(I). P. 16–18.
В статье представлены возрастные особенности метаболизма и клеточного энергообмена у женщин.Показано, что в перименопаузе активность митохондриальных ферментов ниже, чем в репродуктивном возрасте. В период перименопаузы наблюдается снижение клеточного энергообмена и синтеза половых гормонов.
Введение
Старение населения во всем мире признано одной из глобальных проблем человечества. Если в 1990 г. во всем мире женщин в возрасте 50 лет и старше было 476 млн, то уже в 2013 г. их стало около 800 млн, а через 15 лет этот показатель увеличится почти в полтора раза [1]. Но наши знания о причинах и механизмах старения все еще ограничены.Существуют различные теории патогенеза старения, в т. ч. митохондриальная, которая впервые была предложена в 1978 г. [2]. Согласно этой теории в основе старения лежит прогрессивная потеря функциональной активности митохондрий в различных тканях организма.
Как известно, митохондрия отвечает за создание энергии в форме аденозинтрифосфата (АТФ) и участвует в механизмах передачи сигналов апоптоза.
 Нарушение клеточной энергетики приводит к полисистемным поражениям, сопровождает практически все патологические процессы, в т. ч. старение, поэтому большой интерес представляют изучение энергетического статуса человека и разработка диагностических критериев митохондриальной дисфункции как типового патологического процесса, не имеющего этиологической и нозологической специфики [3–5].
Нарушение клеточной энергетики приводит к полисистемным поражениям, сопровождает практически все патологические процессы, в т. ч. старение, поэтому большой интерес представляют изучение энергетического статуса человека и разработка диагностических критериев митохондриальной дисфункции как типового патологического процесса, не имеющего этиологической и нозологической специфики [3–5].В пользу такой точки зрения свидетельствуют накопление в митохондриальной ДНК больших делеций и точечных мутаций в тканях пожилых индивидуумов и уменьшение количества копий, снижение с возрастом активности ферментов, обеспечивающих перенос электронов в дыхательной цепи, в лимфоцитах, скелетных мышцах и кардиомиоцитах. Как следствие этих процессов происходит увеличение продукции активных форм кислорода и прогрессивное перекисное окисление липидов и белков мембран клеток, изменения морфологической структуры митохондрий и снижение мембранного потенциала митохондрий, обеспечивающего энергию для синтеза АТФ [2, 6].
С возрастом у человека одной из первых начинает угасать репродуктивная функция. При старении организма понижается чувствительность всех звеньев оси «гипоталамус — гипофиз — гонады» к сигналам прямой и отрицательной обратной связи от уровня управляющих структур мозга и до органов-мишеней [7]. Нереализованность регуляторных сигналов приводит к гиперфункции и истощению гипоталамуса, гипофиза и яичников. Разобщение регуляторных взаимоотношений в оси «гипоталамус–гипофиз–яичники» нарушает ритм менструального цикла и, наконец, овуляторный цикл прерывается и репродуктивная функция утрачивается. Показано, что метаболическая терапия с использованием митохондриальных субстратов способна существенно повышать фертильность [8], имеет «омолаживающий» эффект с симпатотонической активацией нейроэндокринной системы [9].
Разработаны многочисленные способы диагностики митохондриальной дисфункции. Наиболее простыми, доступными и достаточно точными являются методы, основанные на определении активности митохондриальных ферментов в лимфоцитах периферической крови.
 Чаще всего исследуется активность двух окислительно-восстановительных дегидрогеназ: сукцинатдегидрогеназы (СДГ) и α-глицерофосфатдегидрогеназы (α-ГФДГ).
Чаще всего исследуется активность двух окислительно-восстановительных дегидрогеназ: сукцинатдегидрогеназы (СДГ) и α-глицерофосфатдегидрогеназы (α-ГФДГ).Проведенное нами ранее исследование показателей клеточного энергообмена у женщин с нарушением фертильности показало значительное (почти двукратное) снижение активности СДГ и α-ГФДГ по сравнению с фертильными женщинами [10].
Цель настоящего исследования: в сравнении изучить метаболизм и клеточный энергообмен у женщин репродуктивного возраста и в перименопаузе.
Материал и методы
Биохимическое и цитохимическое исследование крови выполнено у 50 небеременных женщин, которые по возрасту были поделены на 2 группы: 1-я (контрольная) группа — 25 женщин раннего репродуктивного возраста (средний возраст 25,7±3,8 года), критерии включения в исследование: возраст 18–44 лет, регулярный менструальный цикл; 2-я (основная) группа — 25 женщин в возрасте 45 лет и старше (средний возраст 48,9±3,1 года), критерии включения: возраст 45 лет и старше, отсутствие менопаузы.
Критерии исключения из исследования: беременность, менопауза, прием препаратов, содержащих половые гормоны.
Для оценки состояния клеточного энергетического обмена определяли активность СДГ и a-ГФДГ в лимфоцитах периферической крови цитохимическим методом в модификации Р.П. Нарциссова, с последующей визуальной морфометрией [11]. Показатели белкового, углеводного, липидного обмена (общий белок, альбумин, билирубин общий и конъюгированный, мочевина, креатинин, мочевая кислота, глюкоза, гликированный гемоглобин, холестерин, липопротеиды высокой плотности, липопротеиды низкой плотности (ЛПНП), триглицериды), активность цитоплазматических ферментов: аспартатаминотрансферазы, аланинаминотрансферазы, лактатдегидрогеназы, щелочной фосфатазы определяли в образцах венозной крови на биохимическом автоматическом анализаторе «Sapphire 400» (Япония) с использованием унифицированных тест-систем.

Статистический анализ полученных данных выполнен с использованием пакета прикладных программ Statistica for Windows 7.0. Описательная статистика включала расчет средней величины с учетом стандартной ошибки среднего (M±m). Равенство выборочных средних проверяли по t-критерию Стьюдента. Для оценки значимости различий между качественными признаками использован критерий хи-квадрат. Критический уровень значимости (р) при проверке гипотез равен 0,05.
Результаты и обсуждение
Период полового созревания характеризовался своевременным менархе у 20 (80%) женщин основной группы и 22 (88%) — контрольной группы. Средний возраст наступления менархе составил 13,2±0,26 года в основной и 12,8±0,43 в контрольной группе. Раннее становление менструальной функции выявлено у 2-х (8%),позднее — у 3-х (12%) пациенток основной группы, в контроле — у 3-х (12%) и 4-х (16%) соответственно. В основной группе менструации установились сразу только у 10 (40%) пациенток, у 15 (60%) — отмечено позднее установление менструального цикла (в течение 1,5–2 лет с момента менархе), тогда как в контрольной группе у 18 (72%) пациенток сразу установился регулярный цикл. На момент обследования все пациентки контрольной группы характеризовали менструальный цикл как регулярный, длительностью 27,6±0,83 дня с колебаниями от 21 до 35 дней. Длительность менструального кровотечения не превышала 7 дней у всех обследованных женщин и составила, в среднем 5,12±0,27 дня.Менопауза у пациенток основной группы — естественная (100%), наступила после 45 лет (49±2,6), длительность — 1–3 года. У 14 (64%) пациенток основной
группы наблюдались заболевания, обусловленные нарушением нейрогуморальной регуляции: дискинезия желудочно-кишечного тракта и желчевыводящих путей —
у 7 (28%) пациенток, нейроциркуляторная дистония
по гипертоническому типу — у 10 (40%) (p<0,01). На момент обследования обострения хронических экстрагенитальных заболеваний не отмечалось.
 Избыточная масса тела и ожирение (индекс массы тела >25 кг/м2) чаще встречались у женщин в основной группе, чем в контрольной (76% (19) против 28% (7), p<0,01).
Избыточная масса тела и ожирение (индекс массы тела >25 кг/м2) чаще встречались у женщин в основной группе, чем в контрольной (76% (19) против 28% (7), p<0,01).В основной группе климактерический синдром был выявлен у 56% (14) женщин, в т. ч. средней и тяжелой степени — у 28% (7).
Исследование активности митохондриальных ферментов в лимфоцитах крови выявило достоверные отличия между группами. Максимальная активность ферментов наблюдалась у женщин репродуктивного возраста (1-я группа): 21,22±1,23 ед. СДГ и 12,6±0,45 ед. α-ГФДГ, в перименопаузе активность ферментов энергообмена снизилась до 15,5±0,94 ед. СДГ (р<0,001) и 8,94±0,98 ед. α-ГФДГ (р<0,05).
Сравнительный анализ показателей белкового обмена (общего белка, альбумина, мочевины, креатинина) выявил повышение средней концентрации всех исследуемых метаболитов с увеличением возраста. Так, например, содержание общего белка и альбумина составило в 1-й (контрольной) группе 75,1±1,1 и 48,3±0,6 г/л, во 2-й (основной) группе — 77,3±2,5 и 51,4±3,1 г/л соответственно, однако отличия были недостоверны.
Содержание мочевой кислоты во 2-й группе (310,9±15,3 мкмоль/л) было достоверно выше, чем у женщин репродуктивного возраста контрольной группы (209,9±10,8 мкмоль/л) (р<0,001).
Мочевая кислота является конечным продуктом метаболизма пуринов человека и высших позвоночных. Однако в настоящее время установлена и другая ее функция в организме — антиоксидантная. Анионы мочевой кислоты обладают способностью захватывать активные формы кислорода [12]. Возможно, повышение содержания мочевой кислоты у женщин во время перименопаузы объясняется компенсаторной реакцией организма на развитие оксидативного стресса и снижение активности антиоксидантной системы в этот период.
Содержание липидов крови (холестерин и триглицериды) с возрастом увеличивались: холестерин в 1-й группе — 4,8±0,2 ммоль/л, во 2-й группе — 6,7±0,3 ммоль/л (р<0,01). Содержание триглицеридов в 1-й группе — 0,64±0,06 ммоль/л, в 2-й группе — 1,86±0,31ммоль/л (р<0,01).

Содержание глюкозы с возрастом также увеличивалось: в 1-й группе — 5,2±0,1 ммоль/л, во 2-й группе — 5,5±0,5 ммоль/л (p>0,05). У обследованных пациенток не было выявлено сахарного диабета. Среди женщин старше 45 лет было отмечено 2 случая предиабета (гликемия натощак ≥6,1 ммоль/л и <7,0 ммоль/л). У более молодых женщин нарушений углеводного обмена не выявлено.
По данным ВОЗ, сахарный диабет является основной причиной развития сердечно-сосудистых заболеваний и занимает 3-е место в мире среди причин смертности. Сахарный диабет 2-го типа развивается постепенно и на первых порах незаметно для пациента. Состояние, предшествующее манифестации заболевания, называют предиабетом. Результаты первого национального эпидемиологического исследования NATION показали, что в России 19,3% населения, или около 20,7 млн человек, живут с предиабетом [13].
Таким образом, с возрастом возникает тенденция к ожирению и атеросклерозу, сахарному диабету 2-го типа, особенно в период перименопаузы, когда происходит естественное снижение синтеза половых гормонов. Как известно, менопауза является пусковым моментом в развитии менопаузального метаболического синдрома. Этот синдром относится к поздним обменным нарушениям менопаузы и представляет собой сочетание метаболических, гормональных и клинических нарушений у женщин в постменопаузе [14, 15]. Синдром объединяет абдоминально-висцеральное ожирение, инсулинорезистентность и гиперинсулинемию, дислипидемию, артериальную гипертензию, нарушение толерантности к глюкозе и сахарный диабет 2-го типа, ранний атеросклероз, ишемическую болезнь сердца, нарушения гемостаза, гиперурикемию, подагру, микроальбуминурию, гиперандрогению яичникового генеза. Клиническая значимость всех этих патологических изменений в конечном итоге заключается в том, что их сочетание в значительной степени ускоряет развитие и прогрессирование атеросклероза и сердечно-сосудистых заболеваний, которые являются самой частой причиной смертности среди населения высокоразвитых стран [16].

Полученные данные позволяют по-новому взглянуть на механизмы старения репродуктивной системы. В настоящее время ввиду увеличения продолжительности жизни женщин это перспективное направление научного поиска крайне важно для сохранения репродуктивного потенциала и профилактики заболеваний старости на популяционном уровне [17].
Выводы
В результате проведенного исследования были установлены возрастные особенности метаболизма и клеточной биоэнергетики.1. Высокая активность митохондриальных ферментов определяется у женщин репродуктивного возраста, в перименопаузе показатели снижаются.
2. Содержание основных метаболитов (общего белка, альбумина, глюкозы, холестерина, ЛПНП, триглицеридов, мочевой кислоты) с возрастом повышается, вероятно, это связано как со снижением процессов синтеза половых гормонов, так и со снижением клеточного энергообмена в период перименопаузы.
3. Изучение процессов внутриклеточного энергообмена на различных этапах онтогенеза является перспективным направлением научного поиска, крайне важного для сохранения репродуктивного потенциала и профилактики заболеваний старости на популяционном уровне.
Литература
1. Lutz W., Butz W.P. World population and human capital in the twenty-first century: an overview // Oxford University Press. 2017. 234 p.
2. Лобачев А.Н. Роль митохондриальных процессов в развитии и старении организма. Старение и рак. М.: ВИНИТИ, 1978. 49 с. [Lobachev A.N. Rol’ mitohondrial’nyh processov v razvitii i starenii organizma. Starenie i rak. M.:VINITI, 1978. 49 s. (in Russian)].
3. Сухоруков В.С., Николаев Е.А. Нарушение клеточного энергообмена у детей: сборник материалов. М.: Атес Медика Софт, 2004. С.4-18 [Suhorukov V.S., Nikolaev E.A. Narushenie kletochnogo ehnergoobmena u detej: sbornik materialov. M.: Ates Medika Soft, 2004. S.4–18 (in Russian)].
M.: Ates Medika Soft, 2004. S.4–18 (in Russian)].
4. Царегородцев А.Д., Сухоруков В.С. Митохондриальная медицина — проблемы и задачи // Российский вестник перинатологии и педиатрии. 2012. №4. С.4–13 [Caregorodcev A.D., Suhorukov V.S. Mitohondrial’naya medicina — problemy i zadachi // Rossijskij vestnik perinatologii i pediatrii. 2012. №4. S.4–13 (in Russian)].
5. Di Mauro S., Schon E.A. Mitochondrial disorders in the nervous system // Annu. Rev. Neurosci. 2008. Vol. 31. P.91–123.
6. Скулачев В.П. Новые сведения о биохимическом механизме запрограммированного старения организмов и антиоксидантной защите митохондрий // Биохимия. 2009. №12. С.1718–1721. [Skulachev V.P. Novye svedeniya o biohimicheskom mekhanizme zaprogrammirovannogo stareniya organizmov i antioksidantnoj zashchite mitohondrij // Biohimiya. 2009. №12. S.1718–1721(in Russian)].
7. Дильман В.М. Большие биологические часы. Л.: Медицина, 1982. 208 с. [Dil’man V.M. Bol’shie biologicheskie chasy. L.: Medicina, 1982. 208 s. (in Russian)].
8. Bentov Y., Esfandiari N. et al. The use of mitochondrial nutrients to improve the outcome of infertility treatment in older patients //Fertility and Sterility. 2010. Vol. 93(1). P.272–275.
9. Васильева А.А. Исследование сукцинатсодержащего препарата как средства облегчения симптомов менопаузы // Medline.ru. 2014. №1. С.61–73 [Vasil’eva A.A. Issledovanie sukcinat-soderzhashchego preparata kak sredstva oblegcheniya simptomov menopauzy // Medline.ru. 2014. №1. S.61–73(in Russian)].
10. Пестряева Л.А., Нигматова Е.А., Шейко Л.Д., Маркова Т.В. Активность ферментов клеточного энергообмена в прогнозе эффективности вспомогательных репродуктивных технологий // Клиническая лабораторная диагностика. 2014. №7. С.14–17 [Pestryaeva L.A., Nigmatova E.A., SHejko L.D., Markova T.V. Aktivnost’ fermentov kletochnogo ehnergoobmena v prognoze ehffektivnosti vspomogatel’nyh reproduktivnyh tekhnologij // Klinicheskaya laboratornaya diagnostika. 2014. №7. S.14–17 (in Russian)].
11. Лабораторные методы исследования в клинике: справочник / под ред. проф. В.В. Меньшикова. М.: Медицина, 1987. 368 с. [Laboratornye metody issledovaniya v klinike: spravochnik / pod red. prof. V.V. Men’shikova. M.: Medicina, 1987. 368 s. (in Russian)].
12. Титов В.Н., Ощепкова Е.В. и др. Гиперурикемия — показатель нарушения биологических функций эндоэкологии и адаптации, биологических реакций экскреции, воспаления и артериального давления // Клиническая лабораторная диагностика. 2012. №4. С.3–14 [Titov V.N., Oshchepkova E.V. i dr. Giperurikemiya-pokazatel’ narusheniya biologicheskih funkcij ehndoehkologii i adaptacii, biologicheskih reakcij ehkskrecii, vospaleniya i arterial’nogo davleniya. Klinicheskaya laboratornaya diagnostika. 2012. №4. S.3–14 (in Russian)].
13. Дедов И.И. Распространенность сахарного диабета 2-го типа у взрослого населения России (исследование NATION) // Сахарный диабет. 2016. №2. С.104–112 [Dedov I.I. Rasprostranennost’ saharnogo diabeta 2 tipa u vzroslogo naseleniya Rossii (issledovanie NATION) // Saharnyj diabet. 2016. №2. S.104–112 (in Russian)].
14. Cobin R.H., Goodman N.F. AACE Reproductive endocrinology scientific committee. American assciation of clinical endocrinologists and american college of endocrinology position statement on menopause // Endocr. Pract. 2017. Vol. 23(7). P.869–880.
15. Pucci G., Alcidi R. et al. Sex- and gender-related prevalence, cardiovascular risk and therapeutic approach in metabolic syndrome: A review of the literature // Pharmacol. Res. 2017. Vol. 120. P.34–42.
16. Reaven G.M. The metabolic syndrome: requiescat in pace // Clin. Chem. 2005. Vol. 51. P.931–938.
17. Сметник В.П. Старение репродуктивной системы женщины: клинико-гормональное обоснование стадий, терминология // Доктор.ру. 2014. №12. С.13–14 [Smetnik V.P. Starenie reproduktivnoj sistemy zhenshchiny: kliniko-gormonal’noe obosnovanie stadij, terminologiya// Doktor.ru. 2014. №12. S.13–14 (in Russian)].
Контент доступен под лицензией Creative Commons «Attribution» («Атрибуция») 4.
 0 Всемирная.
0 Всемирная.Поделитесь статьей в социальных сетях
Порекомендуйте статью вашим коллегам
ОСОБЕННОСТИ МЕТАБОЛИЗМА МЕЖКЛЕТОЧНОГО ВЕЩЕСТВА У ПАЦИЕНТОК С ОСТЕОАРТРИТОМ И ДИСПЛАЗИЕЙ СОЕДИНИТЕЛЬНОЙ ТКАНИ | Тюрин
1. Алексенко Е.Ю., Говорин А.В. Особенности клинических проявлений дисплазий соединительной ткани у больных остеоартрозом. Кубанский научный медицинский вестник 2009;111(6):7–9. [Alexeyenko E.Yu., Govorin A.V. Peculiarities of clinical manifestations of connective tissue dysplasia with patients with osteoarthritis. Kuban Scientific Medical Herald 2009;111(6):7–9. (In Russ.)].
2. Царегородцев А.Г. Сосудистая патология при дисплазии соединительной ткани в танатогенезе при внезапной смерти лиц молодого возраста. Сибирский медицинский журнал 2009;24(1):34–9. [Tsaregorodtsev A.G. Vascular pathology with connective tissue dysplasia in tanatogenesis with sudden death of young people. Siberian Medical Journal 2009;24(1):34–9. (In Russ.)].
3. Яковлев В.М., Нечаева Г.И. Системные дисплазии соединительной ткани: актуальность проблемы в клинике внутренних болезней. Сибирский медицинский журнал 2011; 26(3):9–13. [Yakovlev V.M., Nechayeva G.I. Systemic cases of connective tissue dysplasia: actuality of the problem in internal diseases clinic. Siberian Medical Journal 2011;26(3):9–13.
4. Кадурина Т.И., Горбунова В.Н. Дисплазия соединительной ткани: руководство для врачей. СПб.: Элби, 2009. [Kadurina T.I., Gorbunova V.N. Connective tissue dysplasia: manual for physicians. St. Petersburg: Elbi, 2009. (In Russ.)].
5. http://rheumatology.org.ua/blog/articles/392/.
http://rheumatology.org.ua/blog/articles/392/.
6. Chen H.C., Shah S.H., Li Y.J. et al. Inverse association of general joint hypermobility with hand and knee osteoarthritis and serum cartilage oligomeric matrix protein levels. Arthritis Rheum 2008;58(12):3854–64.
7. Русова Т.В., Баитов В.С. Структура гликозаминогликанов у больных остеоартрозом: влияние топографии коленного сустава. Бюллетень СО РАМН 2012;32(2):54–60. [Rusova T.V., Baitov V.S. The structure of glycosaminoglycans of patients with osteoarthritis: effect of the knee joint topography. Bulletin of SO RAMS 2012;32(2):54–60. (In Russ.)].
8. Stubendorff J.J., Lammentausta E., Struglics A. et al. Is cartilage sGAG content related to early changes in cartilage disease? Implications for interpretation of dGEMRIC. Osteoarthritis Cartilage 2012;20(5):396–404.
9. Altman R., Alarcon G., Appelrouth D. et al. The American College of Rheumatology criteria for the classification and reporting of osteoarthritis of the hand. Arthritis Rheum 1990;33(11):1601–10.
10. Altman R., Alarcon G., Appelrouth D. et al. The American College of Rheumatology criteria for the classification and reporting of osteoarthritis of the hip. Arthritis Rheum 1991;34(5):505–14.
11. Altman R., Asch E., Bloch D. et al. Development of criteria for the classification and reporting of osteoarthritis. Classification of osteoarthritis of the knee. Diagnostic and Therapeutic Criteria Committee of the American Rheumatism Association. Arthritis Rheum 1986;29(8):1039–49.
12. Шараев П.Н., Пишков В.Н., Зубарев О.Н. и др. Биохимические методы анализа показателей обмена биополимеров соединительной ткани. Методические рекомендации. Ижевск, 1990. [Sharayev P.N., Pishkov V.N., Zubarev O.N. et al. Biochemical methods of analysis of indicators of connective tissues metabolism biopolymers. Methodological Recommendations. Izhevsk, 1990. (In Russ.)].
13. Sowers M.F., Karvonen-Gutierrez C.A., Yosef M. et al. Longitudinal changes of serum COMP and urinary CTX-II predict x-ray defined knee osteoarthritis severity and stiffness in women. Osteoarthritis Cartilage 2009;17(12):1609–14.
14. Наследственные нарушения соединительной ткани. Российские рекомендации. Кардиоваскулярная терапия и профилактика 2009;8(6 прил 5):1–24. [Hereditary connective tissue disorders. Russian recommendations. Kardiovaskylyarnaya terapiya i profilaktika = Cardiovascular therapy and prevention 2009;8(6 annex 5): 1–24. (In Russ.)].
15. Нечаева Г.И. Дисплазия соединительной ткани: основные клинические синдромы, формулировка диагноза, лечение. Лечащий врач 2008;(2):22–8. [Nechayeva G.I. Connective tissue dysplasia: main clinical syndromes, diagnosis wording, therapy. Lechashchiy vrach = Attending Physician 2008;(2):22–8. (In Russ.)].
16. Эверт Л.С., Бороздун С.В., Боброва Е.И. и др. Диагностика дисплазии соединительной ткани с использованием биомаркеров. Журнал Сибирского федерального университета. Химия 2009;2(4):385–90. [Evert L.S., Borozdun S.V., Bobrova E.I. et al. Diagnostics of connective tissue dysplasia with the use of biomarkers. Journal of the Siberian Federal University. Chemistry 2009;2(4):385–90. (In Russ.)].
17. Бикбов М.М., Даутова З.А., Саматова P.P. и др. Некоторые биохимические показатели крови при приобретенной близорукости у детей. Вестник Российской военно-медицинской академии 2009; 3):52–4. [Bikbov M.M., Dautova Z.A., Samatova R.R. et al. Certain biochemical blood Indicators with acquired myopia of children. Herald of the Russian Military Medical Academy 2009;(3):52–4. (In Russ.)].
18. Aslam I., Perjar I., Shi X.A. et al. Associations between biomarkers of joint metabolism, hand osteoarthritis, and hand pain and function: the Johnston County Osteoarthritis Project. J Rheumatol 2014; 41(5):938–44.
19. Valdes A.M., Meulenbelt I., Chassaing E. et al. Large scale meta-analysis of urinary C-terminal telopeptide, serum cartilage oligomeric protein and matrix metalloprotease degraded type II collagen and their role in prevalence, incidence and progression of osteoarthritis. Osteoarthritis Cartilage 2014;22(5):683–9.
20. Jordan J.M. Cartilage oligomeric matrix protein as a marker of osteoarthritis. J Rheumatol Suppl 2004;70:45–9.
21. Васильева Л.В., Стародубцева И.А., Барсукова Н.А. Лечение больных остеоартрозом с использованием низкоинтенсивного лазерного излучения. Научно-практическая ревматология 2008;(3):85–9. 21. [Vasiliyeva L.V., Starodubtseva I.A., Barsukova N.A. Therapy for parients with osteoarthritis with the use of low-intensity laser radiation. Nauchno-practicheskaya revmatologiya = Scientific and Practical Rheumatology 2008;(3):85–9. (In Russ.)].
22. Briggs M.D., Brock J., Ramsden S.C., Bell P.A. Genotype to phenotype correlations in cartilage oligomeric matrix protein associated chondrodysplasias. Eur J Hum Genet 2014;22(11): 1278–82.
23. Briggs M.D., Mortier G.R., Cole W.G. et al. Diverse mutations in the gene for cartilage oligomeric matrix protein in the pseudoachondroplasia-multiple epiphyseal dysplasia disease spectrum. Am J Hum Genet 1998;62(2):311–9.
ОСОБЕННОСТИ МЕТАБОЛИЗМА КОСТНОЙ ТКАНИ У ДЕТЕЙ С ПЕРЕЛОМАМИ КОНЕЧНОСТЕЙ | Галятина
1. The Bone and Joint Decade 2000–2010 for prevention and treatment of musculo-skeletal disorders // Acta Orthop. Scand. 1998. V. 69. Suppl. 281. P. 1–80.
2. Садовой М.А., Фаламеева О.В. Остеопороз в детском, подростковом и юношеском возрасте: патогенетические механизмы и основы профилактики. Новосибирск: АНО «Клиника НИИТО», 2009. 280 с.
3. Новиков П.В. Рахит и наследственные рахитоподобные заболевания у детей: диагностика, лечение, профилактика. М.: Триада-Х, 2006. 336 с.
4. Беневоленская Л.И., Лесняк О.М. Остеопороз: диагностика, профилактика и лечение. М.: ГЭОТАР-Медиа, 2005. 293 с.
5. Короткова Т.А. Характеристика костной ткани подростков по оценке показателей минерализации: автореф. дис. … канд. мед. наук. М., 2007. 23 c.
6. Soontrapa S., Bunyaratavei N. // J. Med. Assoc. Thai. 2005. Vol. 99. Suppl. 5. P. S29–S32.
7. Huesa C., Yadav M.C., Finnilä M.A.J., Goodyear S.R., Rob-ins S.P., Tanner K.E., Aspden R.M., Millán J.L., Farquharson C. Phospho1 is essential for mechanically competent mineralization and the avoidance of spontaneous fractures // Bone. 2011. V. 48, № 5. P. 1066–1074.
8. Петеркова В.А, Нагаева Е.В. Опыт применения гормона роста при различных вариантах низкорослости у детей // Вопросы современной педиатрии. 2009. Т. 8, № 2. С. 86–93.
9. Строев Ю.И., Чурилов Л.П. Эндокринология подростков / под ред. А.Ш. Зайчика. СПб.: ЭЛБИ-СПБ, 2004. 384 с.
10. Витебская А.В., Смирнова Г.Е., Ильин А.В. Витамин D и показатели кальций-фосфорного обмена у детей, проживающих в средней полосе России в период максимальной инсоляции // Остеопороз и остеопатии. 2010. № 2. С. 3–6.
11. Ермакова И.П., Бузулина В.П., Пронченко И.А., Дудов Н.С. Содержание маркера формирования кости ами-нотерминального пропептида коллагена первого типа в сыворотке крови здоровых мужчин // Клинич. лаб. диаг-ностика. 2007. № 5. С. 13–15.
12. Храмцова С.Н., Щеплягина Л.А., Моисеева Т.Ю. Маркеры костного ремоделирования у детей // Рос. педиатр. журн. 2006. № 4. С. 17–21.
13. Никонова Т.А., Довгаль Д.А., Хохлова О.И., Устьянцева И.М. Особенности соматотропной регуляции кальциевого обмена у детей с патологией опорно-двигательного аппарата // Политравма. 2012. № 1. С. 52–53.
14. Никонова Т.А., Хохлова О.И., Устьянцева И.М., Довгаль Д.А. Маркеры костного ремоделирования у детей с врожденной и приобретенной патологией опорно-двигательного аппарата // Политравма. 2012. № 3. С. 68–71.
15. Щеплягина Л.А., Моисеева Т.Ю. Проблемы остеопороза в педиатрии: возможности профилактики // Рус. мед. журн. 2003. Т. 11, № 27. С. 1554–1556.
16. Свешников А.А.,. Каминский А.В., Ларионова Т.А. Изменение концентрации остеотропных гормонов, остеокальцина, гормонов стресс-группы при лечении чреc- и межвертельных переломов // Гений ортопедии. 2007. № 3. С. 48–51.
17. Хвостова С.А., Свешников К.А. Роль гормонов эндокринных желез в репаративном костеобразовании // Современные проблемы науки и образования. 2008. № 2. С. 52–56.
18. Ahmad A.M., Hopkins M.T., Thomas J., Durham B.H., Fraser W.D., Vora J.P. Parathyroid responsiveness to hypocalcemic and hypercalcemic stimuli in adult growth hormone deficiency after growth hormone replacement // Am. J. Physiol. Endocrinol. Metab. 2004. V. 286. P. 986–993.
19. Delmas P.D., Eastell R., Garnero P., Seibel M.J., Stepan J. The Use of Biochemical Markers of Bone Turnover in Osteoporosis // Osteoporos. Int. 2000. V. 6, Suppl. P. 2–17 (includes IOF recommendations).
ОСОБЕННОСТИ МЕТАБОЛИЗМА КОСТНОЙ ТКАНИ У ПАЦИЕНТОВ С МОРБИДНЫМ ОЖИРЕНИЕМ | Юдин
1. Anderson J., Konz E., Frederich R. et al. Long-term weight-loss maintenance: a meta-analysis of US studies // Am. J. Clin. Nutr. — 2001. — Vol. 74. — P. 579-584.
2. Angstadt J., Bodziner R. Peripheral polyneuropathy from thiamine defi ciency following laparoscopic Roux-en-Y gastric bypass // Obes. Surg. — 2005. — Vol. 15. — P. 890-892.
3. Avenell A., Brown T., McGee M. et al. What are the long-term bene- fi ts of weight reducing diets in adults? A systematic review of randomized controlled trials // J. Hum. Nutr. Diet. — 2004. — Vol. 17. м P. 317-335.
4. Brechner R., Farris C., Harrison S. et al. A graded evidence-based summary of evidence for bariatric surgery // Surg. Obes. Relat. Dis. — 2005. м Vol. 1. — P. 430-441.
5. Buchwald H., Avidor Y., Braunwald E. et al. Bariatric surgery: a sys- tematic review and meta-analysis // JAMA. — 2004. — Vol. 292. — P. 1724 -1737.
6. Clements R., Yellumahanthi K., Wesley M. et al. Hyperparathy- roidism and vitamin D deficiency after laparoscopic gastric bypass. Am Surg 2008; 74(6): 474-475.
7. Collazo-Clavell M., Jimenez A., Hodson S. et al. Osteomalacia aft er Roux-en-Y gastric bypass // Endocr. Pract. — 2004. — Vol. 10. — P. 195 — 198.
8. Douketis J., Macie C., Th abane L. et al. Systematic review of long- term weight loss studies in obese adults: clinical signifi cance and applicability to clinical practice // Int. J. Obes. (Lond). — 2005. — Vol. 29. — P. 1153 — 1167.
9. Fleischer J., Stein EM, Bessler M, Della Badia M, Restuccia N. et al. The Decline in Hip Bone Density after gastric bypass surgery is associated with extent of weight loss J Clin EndocrinolMetab 2008; 93: 3735-3740.
10. Interdisciplinary European guidelines for durgery for severe (mor- bid) obesity. Obes surg 2007; 17: 260-70.
11. Kalarchian M, Marcus M, Levine M, et al. Relationship of psychi- atric disorders to 6-month outcomes after gastric bypass. Surg Obes Relat Dis 2008; 4: 544-9.
12. Larad-Jimines A, Diaz-Guerra C, de Cuadros Borajjo P, et al. Short-, mid-, and long-term results of Larrad biliopancreatic diversion. Obes Surg 2007; 17: 202-10.
13. Orzano A., Scott J. Diagnosis and treatment of obesity in adults: an applied evidence based review // J. Am. Board. Fam. Pract. — 2004. — Vol. 17. — P. 359-369.
14. Rhode B., Arseneau P., Cooper B. et al. Vitamin B-12 defi ciency aft er gastric surgery for obesity // Am. J. Clin. Nutr. — 1996. — Vol. 63. — P. 103-109.
15. Singh S., Kumar A. Wernicke encephalopathy from thiamine defi ciency aft er obesity surgery: a systematic review // Neurology. — 2007. — Vol. 68. — P. 807-811.
16. Sjostrom L., Lindroos A-K., Peltonen M. et al. Lifestyle, diabetes and cardiovascular risk factors 10 years aft er bariatric surgery // N. Engl. J. Med. — 2004. — Vol. 351. — P. 2683-2693.
17. Summerbell C., Cameron C., Glasziou P. WITHDRAWN: Advice on low-fat diets for obesity // Cochrane Database Syst. Rev. — 2008. — Vol. 16.
Особенности метаболизма дегидроэпиандростерона и современные возможности его использования для коррекции нарушений репродуктивной функции
Цель исследования. Представить характеристику метаболизма дегидроэпиандростерона (ДГЭА), рассмотреть видовые различия в синтезе ДГЭА.
Материал и методы. В обзор включены данные зарубежных и отечественных статей, найденных в Pubmed по данной теме, опубликованных за последние годы.
Результаты. Доказана определяющая роль ДГЭА как источника образования биологически активных половых стероидов – тестостерона и эстрогенов в периферических тканях. Рассматривается биодоступность и возможные механизмы действия ДГЭА. В экспериментальных и клинических исследованиях выявлена более высокая биодоступность ДГЭА при трансдермальном введении по сравнению с его приемом per os.
Заключение. Большинство современных исследований у мужчин и женщин демонстрируют выраженную зависимость биодоступности ДГЭА в организме в виде водного геля водонерастворимых стероидов. В работе представлена разработанная авторами технология трансдермального введения в виде водного геля водонерастворимых стероидов С18, С19 и С21 ряда с использованием в качестве карьера деацетилированного хитина (хитозана), что позволяет существенно снизить дозу вводимого гормона.
1. Leinonen P., Ruokonen A., Kontturi M., Vihko R. Effects of estrogen treatment on human testicular unconjugated steroid and steroid sulfate production in vivo. J. Clin. Endocrinol. Metab. 1981; 53(3): 569-73.
2. McKenna T.J., DiPietro D.L., Brown R.D., Strott C.A., Liddle G.W. Plasma 17-OH-pregnenolone in normal subjects. J. Clin. Endocrinol. Metab. 1974; 39(5): 833-41.
3. Baulieu E.E., Robel P. Dehydroepiandrosterone and dehydroepiandrosterone sulfate as neuroactive neurosteroids. J. Endocrinol. 1996; 150(Suppl.): S221-39.
4. Goncharov N.P., Katsiya G.V., Dzhokua A.A., Barkaya V.S., Kulava Z.V., Mikvabiya Z.Y. Effect of neurosteroid dehydroepiandrosterone on the higher nervous activity of old non_human primates. Hum. Physiol. 2014; 40(2): 149-55.
5. Labrie F., Dupont A., Belanger A., Cusan L., Lacourciere Y., Monfette G. et al. New hormonal therapy in prostatic carcinoma: combined treatment with an LHRH agonist and an antiandrogen. Clin. Invest. Med. 1982;5(4): 267-75.
6. Khalil M.W., Strutt B., Vachon D., Killinger D.W. Effect of dexamethasone and cytochrome P450 inhibitors on the formation of 7alpha- hydroxydehydroepiandrosterone by human adipose stromal cells. J. Steroid Biochem. Mol. Biol. 1994; 48(5-6): 545-52.
7. Luu-The V., Dufort I., Paquet N., Reimnitz G., Labrie F. Structural characterization and expression of the human dehydroepiandrosterone sulfotransferase gene. DNA Cell Biol. 1995; 14(6): 511-8.
8. Rhéaume E., Simard J., Morel Y., Mebarki F., Zachmann M., Forest M.G. et al. Congenital adrenal hyperplasia due to point mutations in the type II 3 beta-hydroxysteroid dehydrogenase gene. Nat. Genet. 1992; 1(4): 239-45.
9. Labrie F., Sugimoto Y., Luu-The V., Simard J., Lachance Y., Bachvarov D. et al. Structure of human type II 5alpha-reductase gene. Endocrinology. 1992; 131(3): 1571-3.
10. Labrie F., Simard J., Luu-The V., Pelletier G., Bélanger A., Lachance Y. et al. Structure and tissue-specific expression of 3 beta-hydroxysteroid dehydrogenase/ 5-ene-4-ene isomerase genes in human and rat classical and peripheral steroidogenic tissues. J. Steroid Biochem. Mol. Biol. 1992; 41(3-8): 421-35.
11. Luu-The V., Zhang Y., Poirier D., Labrie F. Characteristics of human types 1, 2 and 3 17beta-hydroxysteroid dehydrogenase activities: oxidation/reduction and inhibition. J. Steroid Biochem. Mol. Biol. 1995; 55(5-6): 581-7.
12. Labrie Y., Durocher F., Lachance Y., Turgeon C., Simard J., Labrie C., Labrie F. The human type II 17beta-hydroxysteroid dehydrogenase gene encodes two alternatively spliced mRNA species. DNA Cell Biol. 1995; 14(10): 849-61.
13. Labrie F., Simard J., Luu-The V. The 3-beta-hydroxysteroid dehydrogenase / isomerase gene family: lessions from type II 3beta-HSD congenital deficiency. In: Hansson V., Levy F. O., Tasken K., eds. Signal transduction in testicular cells. Ernst Schering Research Foundation Workshop. Berlin, Heidelberg, New York: Springer-Verlag; 1996: 185-218.
14. Stahl F., Schnorr D., Pilz C., Dörner G. Dehydroepiandrosterone (DHEA) levels in patients with prostatic cancer ,heart diseases and under surgery stress. Exp. Clin. Endocrinol. 1992; 99(2): 68-70.
15. Labrie C., Martel C., Dufour J.M., Lévesque C., Mérand Y., Labrie F. Novel compounds inhibit estrogen formation and action. Cancer Res. 1992; 52(3):610-5.
16. Luo S., Sourla A., Labrie C., Gauthier S., Merand Y., Belanger A., Labrie F. Effect of twenty-four-week treatment with the antiestrogen EM-800 on estrogen-sensitive parameters in intact and ovariectomized mice. Endocrinology. 1998; 139(5): 2645-56.
17. Luu-The V., Labrie C., Simard J., Lachance Y., Zhao H.F., Couët J. et al. Structure of two in tandem human 17 beta-hydroxysteroid dehydrogenase genes. Mol. Endocrinol. 1990; 4(2): 268-75.
18. Labrie F., Bélanger A., Simard J., Van Luu-The, Labrie C. DHEA and peripheral androgen and estrogen formation: intracinology. Ann. N. Y. Acad. Sci. 1995; 774: 16-28.
19. Studd J.W., Chakravarti S., Collins W.P. Plasma hormone profiles after the menopause and bilateral oophorectomy. Postgrad. Med. J. 1978; 54(Suppl. 2): 25-30.
20. Longcope C., Hui S.L., Johnston C.C. Jr. Free estradiol, free testosterone, and sex hormone-binding globulin in perimenopausal women. J. Clin. Endocrinol. Metab. 1987; 64(3): 513-8.
21. Steinberg K.K., Freni-Titulaer L.W., DePuey E.G., Miller D.T., Sgoutas D.S., Coralli C.H. et al. Sex steroids and bone density in premenopausal and perimenopausal women. J. Clin. Endocrinol. Metab. 1989; 69(3): 533-9.
22. Labrie C., Flamand M., Bélanger A., Labrie F. High bioavailability of dehydroepiandrosterone administered percutaneously in the rat. J. Endocrinol. 1996; 150(Suppl.): S107-18.
23. Wolkowitz O.M., Reus V.I., Roberts E., Manfredi F., Chan T., Raum W.J. et al. Dehydroepiandrosterone (DHEA) treatment of depression. Biol. Psychiatry. 1997; 41(3): 311-8.
24. Labrie F., Archer D., Bouchard C., Fortier M., Cusan L., Gomez J.L. et al. Intravaginal dehydroepiandrosterone (Prasterone), a physiological and highly efficient treatment of vaginal atrophy. Menopause. 2009; 16(5): 907-22.
25. Labrie F., Archer D., Bouchard C., Fortier M., Cusan L., Gomez J.L. et al. High internal consistency and efficacy of intravaginal DHEA for vaginal atrophy. Gynecol. Endocrinol. 2010; 26(7): 524-32.
26. Labrie F., Archer D.F., Bouchard C., Fortier M., Cusan L., Gomez J.L. et al. Intravaginal dehydroepiandrosterone ( prasterone), a highly efficient treatment of dyspareunia. Climacteric. 2011; 14(2): 282-8.
27. Labrie F., Archer D.F., Koltun W., Vachon A., Young D., Frenette L. et al. Efficacy of intravaginal dehydroepiandrosterone (DHEA) on moderate to severe dyspareunia and vaginal dryness, symptoms of vulvovaginal atrophy, and of the genitourinary syndrome of menopause. Menopause. 2016; 23(3):243-56.
28. Bouchard C., Labrie F., Derogatis L., Girard G., Ayotte N., Gallagher J. et al. Effect of intravaginal dehydroepi-androsterone (DHEA) on the female sexual function in postmenopausal women: ERC-230 open-label study. Horm. Mol. Biol. Clin. Investig. 2016; 25(3): 181-90.
29. Schmid B.C., Rezniczek G.A., Rolf N., Saade G., Gebauer G., Maul H. Uterine packing with chitosan-covered gauze for control of postpartum hemorrhage. Am. J. Obstet. Gynecol. 2013; 209(3): 225. e1-5.
30. Labrie F., Belanger A., Simard J., Luu-The V., Labrie C. DHEA, The precursor of androgens and estrogens in peripheral tissues in the human: intracrinology. In: Kalimi M., Regelson W., eds. Dehydroepiandrosterone (DHEA). Berlin, New York: Walter de Gruyter; 2000: 299-342.
31. Labrie F., Archer D., Bouchard C., Fortier M., Cusan L., Gomez J.L. et al. Serum steroid levels during 12-week intravaginal dehydroepiandrosterone administration. Menopause. 2009; 16(5): 897-906.
32. Trivedi D.P., Khaw K.T. Dehydroepiandrosterone sulfate and mortality in elderly men and women. J. Clin. Endocrinol. Metab. 2001; 80(9):4171-7.
33. Casson P.R., Lindsay M.S., Pisarska M.D., Carson S.A., Buster J.E. Dehydroepiandrosterone supplementation augments ovarian stimulation in poor responders: a case series. Hum. Reprod. 2000; 15(10):2129-32.
34. Fouany M.R., Sharara F.I. Is there a role for DHEA supplementation in women with diminished ovarian reserve? J. Assist. Reprod. Genet. 2013; 30:1239-44.
35. Zhang J., Qiu X., Gui Y., Xu Y., Li D., Wang L. Dehydroepiandrosterone improves the ovarian reserve of women with diminished ovarion reserve and is a potential regulator of the immune response in the ovaries. Biosci. Trends. 2015; 9(6): 350-9.
36. Kotb M.M., Hassan A.M., AwadAllah A.M. Does dehydroepiandrosterone improve pregnancy rate in women undergoing IVF/ICSI with expected poor ovarian response according to the Bologna criteria? Eur. J. Obstet. Gynecol. Reprod. Biol. 2016; 200: 11-5.
37. Qin J.C., Fan L., Qin A.P. The effect of dehydroepiandrosterone (DHEA) supplementation on women with diminished ovarian reserve (DOR) in IVF cycle. J. Gynecol. Obstet. Hum. Reprod. 2017; 46(1): 1-7.
Поступила 03.02.2017
Принята в печать 17.02.2017
Гончаров Николай Петрович, д.м.н., профессор, зав. лаборатории биохимической эндокринологии и гормонального анализа, ФГБУ Эндокринологический научный центр Минздрава России. Адрес: 115478, Россия, Москва, ул. Москворечье, д. 1. Телефон: 8 (499) 324-95-29. E-mail:
Дедов Иван Иванович, д.м.н., профессор, академик РАН, вице-президент РАН, директор ФГБУ Эндокринологический научный центр Минздрава России.
Адрес: 117036, Россия, Москва, ул. Дмитрия Ульянова, д. 11. Телефон: 8 (499) 500-00-90
Кация Гулинара Викторовна, д.м.н., г.н.с. лаборатории биохимической эндокринологии и гормонального анализа ФГБУ Эндокринологический научный центр
Минздрава России. Адрес: 115478, Россия, Москва, ул. Москворечье, д. 1
Гависова Алла Анатольевна, к.м.н., с.н.с. отделения сохранения и восстановления репродуктивной функции ФГБУ НЦАГиП им. академика В.И. Кулакова
Минздрава России. Адрес: 117997, Россия, Москва, ул. Академика Опарина, д. 4. Е-mail:
Сухих Геннадий Тихонович, д.м.н., профессор, академик РАН, директор ФГБУ НЦАГиП им. академика В.И. Кулакова Минздрава России.
Адрес: 117997, Россия, Москва, ул. Академика Опарина, д. 4. Телефон: 8 (495) 438-18-00
Для цитирования: Гончаров Н.П., Дедов И.И., Кация Г.В., Гависова А.А.,
Сухих Г.Т. Особенности метаболизма дегидроэпиандростерона и современные возможности его использования для коррекции нарушений репродуктивной функции.
Акушерство и гинекология. 2017; 7: 18-26.
http://dx.doi.org/10.18565/aig.2017.7.18-26
(PDF) ОСОБЕННОСТИ МЕТАБОЛИЗМА ГРИБОВ МНОГОЛЕТНЕЙ МЕРЗЛОТЫ
Выпуск 2. Экология грибов. Экстремофилы 195
Анализ грибного разнообразия в большом чис-
ле образцов многолетнемерзлых грунтов, криопэгов,
палеосемян, хранившихся в мерзлых грунтах, частей
ископаемой лошади, обнаруженной в мерзлоте, по-
зволяет выявить повторяющиеся таксоны, имею-
щие высокую частоту встречаемости в холодных ме-
стообитаниях. К ним, в первую очередь, относятся
Pseudogymnoascus pannorum и виды рода Penicillium.
Они встречаются в большинстве изученных образцов.
Грибы этих таксонов вызывают большой интерес у
исследователей. Во многом он связан с поиском актив-
ных при пониженных температурах метаболитов (фер-
ментов и других биологически активных веществ). Из
коллекции палеогрибов ВКМ было исследовано на
способность к образованию вторичных метаболитов
более 80 штаммов рода Penicillium, выделенных из
образцов низкотемпературных экотопов Арктики и
Антарктиды. Было обнаружено 25 вторичных мета-
болитов с уже известной химической структурой и 2
новых для науки метаболита из группы хинолиновых
алкалоидов [10].
Интерес к представителям вида Pseudogymnoascus
pannorum (syn. Geomyces pannorum) определяется тем,
что они являются носителями стратегий выживания в
экстремальных условиях (психротолерантности) [11].
Изучение биологии этих экстремотолерантных
микроскопических грибов, которые являются пер-
спективными моделями функциональной эволюции,
расширяет наши представления об адаптационных
возможностях эукариотов вообще. Не исключено, что
грибы многолетней мерзлоты и, в частности, предста-
вители Pseudogymnoascus pannorum имеют специаль-
ные механизмы репарации клеточных структур, не-
обходимые для выживания при действии различных
стрессоров.
В настоящее время проводится работа по изуче-
нию генетических основ адаптационных способно-
стей этих грибов при воздействии стрессоров, харак-
терных для экотопов Арктики и Антарктиды, – низкой
температуры и пониженной влажности. Анализ пред-
ставленности категорий дифференциально экспрес-
сирующихся генов штамма P. p an no ru m показал суще-
ственное обогащение генов, уменьшающих уровень
экспрессии, генами, входящими в пути контролирую-
щие клеточные деления и пролиферацию, что соответ-
ствует наблюдаемому падению скорости роста этого
организма при действии стрессоров [12].
При этом обращает на себя внимание увеличен-
ное количество генов, понижающих свою экспрессию.
Наиболее интересными для дальнейшего анализа яв-
ляются гены с увеличенным уровнем экспрессии как
в пределах одного, так и нескольких типов условий,
которые могут являться генами, обеспечивающими
метаболизм грибов в неблагоприятных условиях.
Работа была выполнена при поддержке гранта
РФФИ 13-04-02082а.
Список литературы
1. Vishniac HS. e microbiology of Antarctic soils. In:
Friedmann E.I. (ed.) “Antarctic Microbiology”. Wiley-
Liss, New York. 1993: 297-341.
2. Кирцидели И.Ю., Власов Д.Ю., Баранцевич Е.П.,
Крыленков В.А., Соколов В.Т. Комплексы микро-
скопических грибов в почвах и грунтах полярного
острова Известий ЦИК (Карское море). Микол.
фитопатол. 2014; 48(6): 25-31.
3. Власов Д.Ю., Абакумов Е.В., Томашунас В.М., Кры-
ленков В.А., Зеленская М.С. Микобиота почв и ан-
тропогенных субстратов полуострова Ямал. Гигие-
на и санитария. 2014; 5: 49-51.
4. Кочкина Г.А., Иванушкина Н.Е., Карасев С.Г. и др.
Микромицеты и актинобактерии в условиях мно-
голетней естественной криоконсервации. Микро-
биология. 2001; 70(3): 412-20.
5. Gilichinsky DA, Wilson GS, Friedmann EI et all. Mic-
robial populations in Antarctic permafrost: biodiversi-
ty, state, age and implication for astrobiology. Astrobio-
logy. 2007; 2: 275-311.
6. Ozerskaya S, Kochkina G, Ivanushkina N, Gilichinsky
D. Fungi in permafrost. In: Margesin R. (ed.) «Perma-
frost soils». Springer Verlag, Berlin–Heidelberg. 2009:
85-95.
7. Lydolph MC, Jacobsen J, Arctander P et all. Beringian
paleoecology inferred from permafrost-preserved fun-
gal DNA. Appl Environ Microbiol. 2005; 71: 1012-7.
8. Kochkina G, Ivanushkina N, Ozerskaya S et all. An-
cient fungi in Antarctic permafrost environments.
FEMS Microbiol Ecol. 2012; 82(2): 501-9.
9. Ivanushkina NE, Kochkina GA, Ozerskaya S.M. Fun-
gi in ancient permafrost sediments of the Arctic and
Antarctic regions. In: Castello J, Rogers S (eds.) “Life in
ancient ice”. Princeton Press, Princeton. 2005: 127-39.
10. Kozlovsky AG, Zhelifonova VP, Antipova TV. Penicil-
lium fungi from permafrost: biosynthesis of secondary
metabolites, peculiarities of growth and development.
In Pokrovsky O. (Еd.) “Permafrost: distribution, com-
position and impacts on infrastructure and Ecosys-
tems”. CNRS, Toulouse, France and Tomsk State Uni-
versity. Russia. 263-80.
11. Конова И.В., Сергеева Я.Э., Галанина Л.А., Кочки-
на Г.А., Иванушкина Н.Е., Озерская С.М. Липоге-
нез грибов Geomyces pannorum при воздействии
стрессоров. Микробиология. 2009; 78(1): 52-8.
12. Василенко О.В., Пенин А.А., Логачева М.Д. и др.
Анализ экспрессии генов у грибов-экстермофи-
лов Pseudogymnoascus pannorum с использовани-
ем RNA-seq для изучения стратегий выживания в
неблагоприятных условиях среды. Первая Пущин-
ская школа-конференция «Биохимия, физиология
и биосферная роль микроорганизмов». Пущино:
ИБФМ РАН. 2014: 132-34.
Метаболические особенности реакции клетки на опасность
Реакция на опасность клетки (CDR) — это эволюционно консервативный метаболический ответ, который защищает клетки и хозяев от вреда. Это вызвано столкновениями с химическими, физическими или биологическими угрозами, которые превышают способность клетки к гомеостазу. Возникающее в результате метаболическое несоответствие между доступными ресурсами и функциональной способностью вызывает каскад изменений клеточного электронного потока, потребления кислорода, окислительно-восстановительного потенциала, текучести мембран, динамики липидов, биоэнергетики, распределения ресурсов углерода и серы, фолдинга и агрегации белков, доступности витаминов, гомеостаза металлов, метаболизм индола, птерина, 1-углерода и полиамина, а также образование полимеров.Первая волна сигналов опасности состоит из высвобождения промежуточных продуктов метаболизма, таких как АТФ и АДФ, промежуточных продуктов цикла Кребса, кислорода и активных форм кислорода (АФК), и поддерживается пуринергической передачей сигналов. После того, как опасность устранена или нейтрализована, активируется заданная последовательность противовоспалительных и регенеративных путей, чтобы обратить вспять CDR и излечить. Когда CDR сохраняется ненормально, метаболизм всего тела и микробиом кишечника нарушаются, коллективная работа многих систем органов нарушается, поведение изменяется, что приводит к хроническим заболеваниям.Метаболическая память о прошлых стрессовых ситуациях сохраняется в виде измененного содержания митохондриальных и клеточных макромолекул, что приводит к увеличению функциональной резервной емкости посредством процесса, известного как митоцеллюлярный гормезис. Системная форма CDR и ее увеличенная форма, пуринергическая реакция на угрозу жизни (PLTR), находятся под прямым контролем древних путей в мозге, которые в конечном итоге координируются центрами ствола мозга. Хемосенсорная интеграция метаболизма всего тела происходит в стволе мозга и является предпосылкой нормального мозгового, моторного, вестибулярного, сенсорного, социального и речевого развития.Понимание CDR позволяет нам переосмыслить старые концепции патогенеза для широкого спектра хронических, связанных с развитием, аутоиммунных и дегенеративных нарушений. Эти расстройства включают расстройства аутистического спектра (РАС), синдром дефицита внимания и гиперактивности (СДВГ), астму, атопию, глютен и многие другие синдромы пищевой и химической чувствительности, эмфизему, синдром Туретта, биполярное расстройство, шизофрению, посттравматическое стрессовое расстройство (ПТСР). , хроническая травматическая энцефалопатия (ХТЭ), черепно-мозговая травма (ЧМТ), эпилепсия, суицидальные мысли, биология трансплантации органов, диабет, болезни почек, печени и сердца, рак, болезнь Альцгеймера и Паркинсона и аутоиммунные расстройства, такие как волчанка, ревматоидный артрит, рассеянный склероз и первичный склерозирующий холангит.
Метаболические особенности рецидивирующего большого депрессивного расстройства в стадии ремиссии и риск рецидива в будущем
Когорта исследования
В исследование были включены 68 пациентов, не принимавших лекарственные препараты, и наблюдались в течение 2,5 лет (блок-схема: рис. 1). Было также зарегистрировано пятьдесят девять контрольных групп, соответствующих возрасту и полу.
Рис. 1: Блок-схема дизайна исследования.Для анализа были доступны результаты метаболизма 126 из 127 участников. В общей сложности 68 субъектов с историей рецидивирующего большого депрессивного расстройства в стадии ремиссии (rrMDD) прослеживались проспективно в течение 2.5 лет.
Характеристики участников
Пациенты с rrMDD имели длительную историю болезни (25 ± 2,4 года для мужчин и 27 ± 1,9 года для женщин) с высоким общим числом эпизодов MDD (среднее значение = 9,7 ± 2,5 для мужчин; 8,1 ± 1,9 для самок) (таблица 1). Средний балл по шкале HAM-D для субъектов rrMDD составлял 2 и 3 для мужчин и женщин, соответственно. Основным эпидемиологическим различием между мужчинами и женщинами с rrMDD было среднее время до рецидива после включения в исследование (мужчины: 5,5 месяцев IQR = 1.9–7,6; женщины: 10,1 IQR = 4,9–17,2; Таблица 1). Во время наблюдения трое участников возобновили прием антидепрессантов при рецидиве, а двое участников возобновили прием антидепрессантов во время ремиссии во время наблюдения. Это произошло после сбора образцов крови для метаболомики при зачислении и не повлияло на анализ без лекарств.
Таблица 1 Характеристики участников.Обзор метаболизма
Субъекты rrMDD, не принимавшие лекарственные препараты, имели метаболический профиль, который можно было отличить от здорового контроля.Многофакторный анализ показал четкое разделение между двумя группами как у мужчин, так и у женщин (рис. 2A, B). Лучшие различающие метаболиты показаны на рис. 2C, D. Относительное метаболическое воздействие и значимость этих различий показаны на рис. 2E, F. Таблицы S1 и S2 представляют исходные данные, а таблицы S3 и S4 перечисляют порядок ранжирования биохимических путей, которые были нарушены у rrMDD женщин и мужчин, соответственно, по сравнению со здоровыми контрольными группами того же возраста и пола. На рисунке 2G показаны общие и гендерные различия в метаболизме.Анализ основных компонентов (PCA) показал, что метаболомика объясняет до 39,1% фенотипической дисперсии между пациентами с rrMDD и здоровым контролем как у мужчин, так и у женщин (рис. S1AC). Метаболомика объяснила до 43,7% фенотипической дисперсии между женщинами, у которых наблюдались рецидивы депрессивных симптомов, и теми, у которых не рецидивы в течение 2,5 лет проспективного наблюдения (рис. S1B), и до 50,6% у мужчин (рис. S1D).
Рис. 2: Нарушения метаболизма и биохимических путей при рецидивирующем большом депрессивном расстройстве в стадии ремиссии.Самки : A , C , E . Самцы : B , D , F. AB . Многовариантное метаболомическое различение ремиссируемого рецидивирующего большого депрессивного расстройства (rrMDD) от контроля с помощью частичного дискриминантного анализа наименьших квадратов. CD . Порядок ранжирования 25 лучших распознающих метаболитов по переменной важности в оценках прогноза (VIP). EF . График изменения пути следования пузырьков. G . Диаграмма Венна общих и гендерно-зависимых метаболитов для диагностики ррБМП.Красные стрелки указывают на повышенный, а черные стрелки указывают на то, что снижение концентрации было связано с риском rrMDD. Показатели VIP ≥1,5 были значительными. Подопытные rrMDD n = 44 женщины, 23 мужчины. Контроль n = 40 самок, 19 самцов.
В целом, изменения в метаболизме липидов преобладали в метаболической сигнатуре rrMDD. Нарушения липидов составляли 80% из 10 основных нарушений метаболизма у женщин (Таблица S3 и Рис. S1). У мужчин липидные нарушения составили 70% из 10 основных метаболических путей (таблица S4 и рис.S2). Наиболее затронутыми 8 липидными путями были фосфолипиды, жирные кислоты и ацилкарнитины, кардиолипины, два класса сфинголипидов (церамиды и сфингомиелины), эйкозаноиды, желчные кислоты, стерины и негонадные стероиды.
Главным изменением нелипидного пути был метаболизм пуринов (таблицы S3 и S4 и рис. S2). Пиримидины, метаболиты микробиома, ГАМК-глутамат-пирролин-5-карбоксилат-пролин, фолат-1-углерод, инозитол и метаболизм триптофан-серотонина также были изменены у субъектов с rrMDD (рис.2 и S2). Гендерные различия обсуждаются в дополнительных результатах.
Метаболические изменения, общие для мужчин и женщин
При анализе на уровне путей, 16 путей были общими для мужчин и женщин с rrMDD (рис. 2E, F и таблицы S3 и S4). Все 8 липидных путей, обнаруженных как аномальные, были общими как у мужчин, так и у женщин. Остальные 8 общих путей вовлекают различные нелипидные пути.
Ацилкарнитины, кардиолипины и витамин B2
Одним из наиболее устойчивых нарушений липидов, обнаруженных как у мужчин, так и у женщин, было повышение уровня ацил-карнитинов (таблицы S3 и S4), которые являются маркером снижения митохондриального окисления жирных кислот. 30 .У женщин это ограничивалось увеличением ацилкарнитинов со средней длиной цепи (C6 – C10), тогда как у мужчин было повышено содержание ацилкарнитинов с длинной цепью (C12 – C18) и очень длинной цепи (≥C20) (таблицы S5 и S6). Кардиолипины, маркеры сложности и биомассы внутренней митохондриальной мембраны, также были снижены. Фосфатидные кислоты (PA-липиды), предшественники синтеза кардиолипина, были увеличены как у мужчин, так и у женщин (рис. 2G). Метаболизм витамина В2, необходимый для окисления митохондриальных жирных кислот, был снижен как у мужчин, так и у женщин.У мужчин это отражалось в снижении уровня рибофлавина в плазме. У женщин это было связано со снижением уровня флавинадениндинуклеотида в плазме (FAD; рис. 2G, таблицы S5 и S6).
Сфинголипиды
Сфинголипиды (церамиды, сфингомиелины и гликосфинголипиды) являются основными структурными и сигнальными липидами, которые облегчают обмен веществ между лизосомами и плазматической мембраной для регулирования роста и воспаления клеток 31,32 , образуют мембранные липидные плотины 33 , экзосомы высвобождаются из клеток 34 и участвуют в синапсах 35,36,37 .Самцы и самки rrMDD обладали специфической аномалией метаболизма сфинголипидов: снижение 2′-гидроксисфингомиелина SM (d18: 1/26: 0 OH) (рис. 2G). 2′-гидроксилирование (2′-ОН) предшественника жирной кислоты амидаацильной цепи сфинголипидов катализируется пероксисомным ферментом 2′-гидроксилазой жирной кислоты (FA2H) 38 . 2′-гидроксисфингомиелины являются предшественниками 2′-ОН гликосфинголипидов, необходимых для дифференцировки клеток, нейрональной связи, стабильности миелина и обладают противоопухолевыми свойствами 39 .
Эйкозаноиды и оксилипины
И у мужчин, и у женщин были изменения в эйкозаноидных (20-углеродных, полиненасыщенных) липидах, полученных из арахидоновой кислоты. 40 , но конкретные метаболиты и направление изменений различались. У женщин также наблюдалось снижение содержания 18-углеродного оксилипина, полученного из линолевой кислоты 41 , называемого 13 (S) -гидроксиоктадекадиеновой кислотой (13-HODE), которая не изменилась у мужчин. 15 (S) -гидроксиэйкозатетраеновая кислота (15 (S) -HETE) снижалась у женщин, но не у мужчин (рис.2C и таблицы S5 и S6). 13-HODE и 15-HETE являются противовоспалительными и способствующими рассасыванию оксилипинами, которые также обладают противоопухолевым действием 42 . Напротив, у rrMMD-самцов наблюдалось увеличение 3 эйкозаноидов: 11-HETE, 9-HETE и 5-HETE, которые являются провоспалительными медиаторами, вырабатываемыми нейтрофилами, эозинофилами и тучными клетками 43,44 . У мужчин также наблюдалось увеличение сосудорасширяющих и противовоспалительных эпоксиэйкозатриеновых кислот 8,9-EET и 11,12-EET (рис. 2G). Большое количество изменений в метаболизме эйкозаноидов сделало это наиболее статистически значимым изменением пути у мужчин (рис.2F).
Стерины
Стерины необходимы для синтеза холестерина, глюкокортикоидов, стероидных гормонов и желчных кислот. Синтез стеролов требует скоординированной активности ферментов в эндоплазматическом ретикулуме (ER) и митохондриях. Уровень стеринов снижался как у мужчин, так и у женщин, у субъектов с rrMDD (рис. 2G, таблицы S3 и S4). У женщин было снижено содержание 24,25-эпоксихолестерина и холестерилсульфата. У мужчин было снижено содержание предшественников холестерина, 24,25-дигидроланостерола и латостерола (таблицы S5 и S6).
Желчные кислоты
Метаболизм желчных кислот требует скоординированной активности ферментов, расположенных в митохондриях, пероксисомах, ER и микробиоме кишечника 45 . Желчные кислоты — это сигнальные молекулы, которые связываются с несколькими классами ядерных рецепторов (FXR, PXR и CAR) и обеспечивают координацию в реальном времени между приемом пищи, микробиомом, печенью и системами системной детоксикации 46 . Желчные кислоты состоят из холестерина и представляют собой основной путь избавления от избыточного холестерина.И у мужчин, и у женщин снизился уровень желчных кислот в плазме. rrMDD-самки имели пониженные уровни четырех вторичных желчных кислот, конъюгированных с глицином и таурином, включая гликохолевую и таурохолевую кислоты (таблицы S3 – S6 и рис. 2). Нарушения желчных кислот были наиболее статистически значимым изменением единственного пути у женщин (рис. 2E). У мужчин rrMDD снижен уровень вторичной желчной кислоты, дезоксихолевой кислоты, которая образуется в результате дегидроксилирования холевой кислоты нормальными кишечными бактериями.
Пурины
Пуриновые нуклеозиды, аденозин, гуанозин и инозин были снижены, а ксантин повышен как у мужчин, так и у женщин с rrMDD (рис.2G и таблицы S3 и S4). Пуриновые нуклеозиды плазмы образуются путем дефосфорилирования пуриновых нуклеотидов, таких как АТФ, АДФ, АМФ, GMP и IMP. Ксантин представляет собой пуриновое азотистое основание, которое, как было показано, связывает метаболизм пуринов с иммунной системой, памятью и тревогой 47 . Синтез, восстановление и метаболизм пуриновых нуклеотидов de novo зависит от совместной активности внеклеточных, ассоциированных с клеточными мембранами, цитозольных и митохондриальных ферментов.
Метаболомика как диагностический инструмент
Анализ площади под кривой оператора приемника (AUROC) был использован для проверки точности метаболитов, чтобы различать субъектов rrMDD и здоровых людей (рис.S3). Классификатор для женщин использовал 12 метаболитов, что дало AUC 0,83 (95% Cl = 0,68–0,96; чувствительность = 80%, 95% Cl = 0,66–0,89; специфичность = 87%, 95% Cl = 0,74–0,95). Для мужчин в качестве классификатора использовалось семь метаболитов, в результате чего AUC составила 0,83 (95% Cl = 0,64–1,0; чувствительность = 74%, 95% Cl = 0,53–0,87; специфичность = 79%, 95% Cl = 0,57–0,91. ).
Метаболомика как инструмент прогноза
Предикторы, общие для мужчин и женщин
Среднее время до рецидива в течение 2.Последующее наблюдение в течение 5 лет для всех субъектов rrMDD составило 588 дней для женщин и 291 день для мужчин (анализ Каплана – Мейера; рис. 3A, B). Многофакторный анализ позволил отличить метаболомную сигнатуру субъектов, у которых возник рецидив, от тех, у кого этого не было (рис. 3C, D и таблицы S7 и S8). Основные различающие метаболиты показаны на рис. 3E, F. В целом, наиболее предсказуемыми классами метаболитов для обоих полов были сфингомиелины и фосфолипиды (рис. 3G, H). AUROC-анализ был использован для проверки прогностической точности женского классификатора с использованием 7 метаболитов и мужского классификатора, который использовал 3 метаболита (рис.3I, J). Прогнозирующая точность рецидива у женщин составила 0,90 (95% ДИ = 0,69–1,0; чувствительность = 0,88; специфичность = 0,89; рис. 3I, K). Несмотря на отличие от женщин-субъектов rrMDD, точность прогноза рецидива депрессии у мужчин составляла 0,99 (95% ДИ = 0,9–1,0; чувствительность = 0,91; специфичность = 1,0; рис. 3J, L).
Рис. 3: Метаболомические предикторы рецидива рецидивирующего большого депрессивного расстройства (rrMDD).A Анализ Каплана – Мейера латентного периода до рецидива у субъектов с рецидивирующим рецидивирующим большим депрессивным расстройством (rrMDD), женщины, B Мужчины.Пунктирные границы указывают 95% доверительный интервал. Метаболические предикторы рецидива рецидивирующего большого депрессивного расстройства. Самки: C , E , G . Кобели: D , F , H . CD Многовариантная метаболомическая дискриминация субъектов с rMDD, которые испытали рецидив в следующие 2,5 года, и тех, у кого нет, проанализирована с помощью частичного дискриминантного анализа наименьших квадратов. EF Порядок ранжирования 15 лучших распознающих метаболитов по переменной важности в оценках проекции (VIP). GH Пузырьковый график изменений пути. Анализ кривой характеристики оператора приемника (ROC) мультианалитических диагностических классификаторов для rrMDD. Я . В классификаторе для женщин использовано 7 метаболитов. J В классификаторе для мужчин используются три метаболита. Площадь AUC под кривой, точность повторной двойной перекрестной проверки rdCV. KL 2 × 2 анализ таблицы непредвиденных обстоятельств. Анализ пропорциональной опасности Кокса выбранных метаболитов. M Снижение содержания метилцистеина предсказывает более высокий риск рецидива у женщин. N Снижение содержания 15-гидроксиэйкозатетраеновой кислоты (15-HETE) предсказывает более высокий риск рецидива у мужчин. O Снижение содержания моногексозилцерамида (MHC (d18: 1/20: 0)) предсказывало более высокий риск рецидива у женщин. P Снижение уровня β-каротина предсказывало более высокий риск рецидива у мужчин. За субъектами rrMDD проспективно наблюдали в течение 2,5 лет: n = 42 женщины (24 с рецидивом, 18 без рецидива), 20 мужчин (11 с рецидивом, 9 без рецидива).
Три класса метаболитов предсказывали риск рецидива как у мужчин, так и у женщин с помощью анализа пропорциональных рисков Кокса (рис.4 и таблицы S7 – S10). Это было снижение содержания липидов 2′-гидроксисфингомиелинов (2′-OH SM), тригексозилцерамидов (THC) и фосфатидилхолина (PC). Как у мужчин, так и у женщин почти 80% идентифицированных прогностических метаболитов отрицательно коррелировали с риском рецидива (таблицы S9 и S10). Это означает, что когда уровень в крови был низким по сравнению с другими субъектами rrMDD, риск рецидива был выше, и наоборот.
Рис. 4: Диаграмма Венна общих и гендерно-зависимых предикторов рецидива.Красные стрелки указывают на повышенную, а черные стрелки указывают на то, что снижение концентрации было связано с риском рецидива депрессии. За субъектами rrMDD проспективно наблюдали в течение 2,5 лет: n = 42 женщины (24 с рецидивом, 18 без рецидива), 20 мужчин (11 с рецидивом, 9 без рецидива).
Специфические для женщин предикторы рецидива
Все самые сильные предикторы рецидива были специфичны для пола. У женщин самым сильным предиктором был низкий уровень метилцистеина (рис.3K, M и таблица S9). Было обнаружено, что у женщин-субъектов rrMDD с нижней половиной метилцистеина плазмы среднее время до рецидива составляло 1,1 года (403 ± 63 дня, среднее ± SEM). Женщины, входящие в 50-й перцентиль метилцистеина, гораздо медленнее испытывали рецидив (медиана = 2,3 года; 848 ± 58 дней; бета-коэффициент Кокса = -1,8; p <0,00006; Таблица S9 и рис. 3M). Моногексозилцерамид (MHC (d18: 1/20: 0)) был еще одним примером метаболита, защищающего от рецидива в будущем (рис.30 и таблица S9). Двумя метаболитами, повышающими риск рецидива, были лизофосфатидилхолин 16: 0 (LysoPC (16: 0)) и арахидоновая кислота (20: 4). Эти два липида могут быть получены из одного и того же исходного липида фосфатидилхолина, PC (16: 0/20: 4) за счет действия липопротеин-ассоциированной фосфолипазы A2 (LP-PLA 2 ) и других типов PLA 2 в условиях напряжение 48 . Метаболиты, повышающие и снижающие риск рецидива, перечислены в таблице S9. Из 399 измеренных метаболитов 33 были значимыми предикторами риска рецидива у женщин, и у 82% этих метаболитов (27/33) более высокие уровни снижали риск рецидива (таблицы S7 и S9).Корректировка количества предыдущих эпизодов и остаточных симптомов не изменила отношения рисков этих метаболитов.
Мужские предикторы рецидива
Из 399 измеренных метаболитов 17 были значимыми предикторами риска рецидива у мужчин (таблицы S8 и S10). Липид эйкозаноидов, 15-гидроксиэйкозатетраеновая кислота (15-HETE), был основным предиктором рецидива у мужчин. Когда мужчины были разделены на верхний и нижний 50-й процентили, у субъектов rrMDD в нижней половине для этого плазменного 15-HETE было обнаружено, что среднее время до рецидива составляет 0.7 лет (245 ± 85 дней). Мужчины, входящие в верхний 50-й процентиль этого противовоспалительного эйкозаноида, намного медленнее испытывали рецидив (медиана = 1,7 года; 614 ± 108 дней; рис. 3N; бета-коэффициент Кокса = -1,5; p <0,0006; таблица S10). Бета-каротин также был защитным. У мужчин с самым низким 50-м процентилем бета-каротина в плазме среднее время до рецидива составляло 0,5 года (175 ± 103 дней), в то время как у мужчин верхнего 50-го процентиля среднее время до рецидива составляло 1,9 года (677 ± 104 дней; рис.3N и P; Таблица S8 и S10). Повышенный уровень аланина и аллантоина был связан с повышенным риском рецидива у мужчин. Повышенный уровень аланина продуцируется как продукт трансаминирования пирувата и является маркером митохондриальной дисфункции 49 . Аллантоин неферментативно вырабатывается из мочевой кислоты под воздействием активных форм кислорода (АФК) и является маркером окислительного экранирования 50 и стресса 51 . Повышенный уровень 14 из 18 (78%) предикторов у мужчин снизил риск рецидива.Рисунки S4 и S5 иллюстрируют профили рецидивов в стиле Каплана-Мейера для основных прогнозируемых метаболитов, определенных анализом пропорциональных рисков Кокса у женщин и мужчин, соответственно. Корректировка количества предыдущих эпизодов и остаточных симптомов не изменила отношения рисков этих метаболитов, за исключением бета-каротина, SM (d18: 1/20: 2 OH), PS (18: 0/18: 1), PE. (34: 1) или SM (d18: 1/20: 1), где поправка на особенно предыдущие жизненные эпизоды депрессии увеличивала отношения рисков.
Расчет размера выборки для валидационных исследований
Для оценки размера выборки, необходимого в будущих исследованиях для подтверждения результатов этого пилотного исследования, использовались три метода (см. Дополнительные методы).Медиана Pearson r для метаболитов у женщин с оценкой VIP ≥1,5 составила r = 0,2. Средняя разница по Z-баллам составила 0,39 (таблицы S5, S11 – S14). Используя корреляционный анализ и требование для Pearson r ≥ 0,2, общее количество субъектов одного пола (случаи плюс контроль) составило 194. С использованием множественного регрессионного анализа и порогового значения не менее 35 значимых метаболитов и f Коэна. 2 = 0,15, размер исследования 201.Используя пороговое значение Z-балла ≥0,4 (значимые метаболиты в случаях должны отличаться от контроля не менее чем на 0,4 стандартных отклонения), размер исследования составил 198. Средняя оценка составила 198 ± 3,5 (среднее ± стандартное отклонение). Эти результаты показали, что для подтверждения результатов потребуется исследование по меньшей мере 198 женщин и 198 мужчин (99 случаев и 99 контрольных).
Дополнительные результаты см. В дополнительных результатах.
Метаболические характеристики субпопуляций CD8 + Т-клеток у молодых и пожилых людей не позволяют прогнозировать функциональность
T
VM и клетки старых мышей обладают повышенной резервной дыхательной способностьюМы пытались понять, является ли базальный метаболический фенотип T Клетки VM более тесно связаны с популяцией T N или имеют общие характеристики с обычными клетками T MEM , такие как повышенная митохондриальная нагрузка и SRC, а также для понимания того, как эти профили меняются с возрастом.Поэтому мы провели комплексный митохондриальный и метаболический анализ для каждой из этих подгрупп, выделенных из селезенки наивных молодых и старых мышей, свободных от специфических патогенов (SPF).
Базальный митохондриальный метаболический профиль молодых и старых субпопуляций Т-лимфоцитов CD8 + мышей определяли путем проведения мито-стрессового теста с использованием биоанализатора Seahorse XFe96 на отсортированном T N (CD44 lo ), T VM (CD44 hi CD49d lo ) и T MEM (CD44 hi CD49d hi ) CD8 + клетки непосредственно ex vivo.В этом анализе скорость потребления кислорода (OCR) отслеживается в базальном состоянии (OCR Bas ), а затем во время лечения различными ингибиторами митохондрий, чтобы обеспечить измерение максимального OCR (OCR Max ), с разницей между OCR . Bas и OCR Max , представляющие SRC. Клетки T N от молодых и старых мышей имели сопоставимые профили OCR, что приводило к сопоставимым SRC для этих подмножеств (рис. 1a, b). Клетки T VM от молодых мышей имели значительно более высокий OCR, чем клетки T N , как при OCR Bas , так и, что более заметно, при OCR Max , что приводило к значительному увеличению SRC (рис.1а, б), что соответствует фенотипу памяти. Клетки T VM от старых мышей также имели стабильно более высокий OCR, чем клетки от молодых мышей, что приводило к значительно более высокому SRC (рис. 1a, b). Поразительно, что клетки T MEM от молодых мышей не обладают более высоким SRC, чем клетки T N , а скорее имеют самый низкий SRC из всех подмножеств (рис. 1a, b), в отличие от предыдущих отчетов 4 . В то время как значительное увеличение SRC также наблюдалось в клетках T MEM с возрастом (рис.1а, б), он оставался значительно ниже, чем SRC, наблюдаемый для клеток Т VM от старых мышей. Эти данные предполагают, что высокий SRC не является отличительной чертой клеток T MEM , а вместо этого определяет клетки T VM , и что старение приводит к увеличению SRC во всех Т-клетках с фенотипом памяти CD8 + .
Рис. 1: Клетки T VM имеют высокий SRC и CIV, который увеличивается с возрастом.a Скорость потребления кислорода (OCR) во времени для отсортированных клеток T N , T VM и T MEM из селезенки наивных молодых и старых мышей SPF.Стрелки указывают добавление митохондриальных ингибиторов (олигомицин; FCCP; антимицин A / ротенон) или временные точки для оценки SRC (OCR Bas , OCR Max ) ( n = 2–4, 5 экспериментальных повторностей). b Изменение OCR с OCR Bas на OCR Max (SRC) для каждого отсортированного подмножества ( n = 2–4, 5 экспериментальных повторов). c Электронно-микроскопические изображения отсортированных клеток непосредственно ex vivo, масштабная линейка показывает 0,2 мкм (1 экспериментальная повторность). d Конфокальная микроскопия отсортированных клеток непосредственно ex vivo, зеленая флуоресценция — окрашивание цитохромом C, масштабная линейка указывает 2 мкм, что использовалось для определения ( e ) преобладающей морфологии митохондрий (слитых, промежуточных или фрагментированных) для 140 клеток на подмножество и f средний митохондриальный след на клетку, рассчитанный из конфокальных изображений (3 экспериментальных повтора). г BN-PAGE и блот для Cox5a из ETC CIV, с указанием полос для CIV, CII, CIII 2 , CV и суперкомплекса CI / CIII 2 / CIV, рядом с блотом, окрашенным Кумасси (3 экспериментальных повтора) . ч Скорость потребления кислорода (OCR) во времени для отсортированных клеток T VM от молодых мышей с высокой (200 мкМ) или низкой (5 мкМ) дозой этомоксира ( n = 3, 3 экспериментальных повтора). Показано среднее значение ± стандартная ошибка среднего (SEM). NS указывает на незначительность, * указывает на p ≤ 0,05, ** указывает на p ≤ 0,01, непарный тест t .
Корреляция характеристик митохондрий SRC
Часто считается, что увеличение SRC отражает количественные и / или качественные изменения в самих митохондриях, включая (i) более плотные митохондриальные кристы, способствующие процессивности и эффективности потребления кислорода электронно-транспортной цепью (ETC) 3 , (ii) слитная морфология митохондрий 6 и (iii) увеличенная митохондриальная нагрузка или объем 5 .Для оценки этих митохондриальных характеристик клетки T N , T VM и T MEM от молодых и старых мышей были отсортированы, и митохондрии были визуализированы непосредственно ex vivo. По данным электронной микроскопии, не было очевидных различий в размере, плотности или морфологии митохондриальных крист между типами клеток или возрастом (рис. 1c). Когда слияние митохондрий оценивалось с помощью конфокальной микроскопии (рис. 1d), промежуточная или слитая морфология наблюдалась только в меньшинстве клеток, даже для клеток T MEM (рис.1e), предполагая, что слияние митохондрий не требуется для поддержания клеток T MEM . Кроме того, не наблюдались клетки T VM от молодых мышей со слитой морфологией (рис. 1e), несмотря на их высокий SRC. Поразительно, что с возрастом наблюдалось значительное увеличение митохондриального слияния во всех типах клеток, причем этот эффект наиболее очевиден в клетках T VM (рис. 1e). Наконец, митохондриальный след на клетку как показатель митохондриальной нагрузки был самым высоким в клетках T VM и T MEM и значительно увеличивался с возрастом в клетках T N и T VM (рис.1е).
Как правило, морфология, слияние или объем митохондриальных крист плохо коррелировали с высоким SRC, избирательно наблюдаемым в субпопуляции клеток T VM . Чтобы получить более прямую оценку емкости электронной транспортной цепи (ETC), уровни митохондриального ETC Complex IV (CIV; Cox5a) из определенного количества каждой клеточной субпопуляции количественно определяли непосредственно с помощью синего PAGE и иммуноблоттинга. Хотя это не абсолютная корреляция, количество CIV, по-видимому, лучше коррелирует с SRC, чем с митохондриальной нагрузкой или морфологией; а именно CIV был увеличен в клетках T VM по сравнению с клетками T N от молодых мышей, и возрастное увеличение CIV было наиболее заметно в популяции T VM (рис.1г). В совокупности наши анализы митохондриальной нагрузки и морфологии показали, что они в целом позволяют прогнозировать возрастное увеличение SRC. Уровни экспрессии ETC CIV, по-видимому, наиболее точно предсказывают клеточный SRC в зависимости от возраста и подгрупп.
Чтобы обеспечить механистическую основу для увеличения митохондриальной нагрузки / активности в клетках T VM от старых мышей, данные RNA-Seq, ранее полученные из поднаборов T N , T VM и T MEM от молодых и старых мышей 17 опрашивали на предмет транскриптов, участвующих в митохондриальном биогенезе, митофагии и митохондриальном слиянии или делении.Во всех подмножествах Т-клеток наблюдалось зависимое от возраста снижение транскриптов Atg101 и Ulk1 , которые являются критическими для митофагии (дополнительный рис. 1a). Наблюдалось соответствующее увеличение количества транскриптов PGC-1α ( Ppargcla ) с возрастом в популяциях обоих фенотипов памяти, что особенно заметно в подмножестве T VM (дополнительный рис. 1a). Анализ уровней транскриптов, связанных со слиянием митохондрий ( Mfn1 , Mfn2 , Opa1 ) или делением ( Dnm1l ), выявил минимальные изменения или их отсутствие в субпопуляциях Т-клеток или со старением (дополнительный рис.1в). В совокупности, хотя для истинного определения влияния митохондриальной динамики на митохондриальную нагрузку требуется более подробный биохимический анализ, эти транскрипционные данные подчеркивают, что с возрастом может происходить уменьшение деградации митохондрий и увеличение биогенеза, особенно в подгруппе T VM , что может способствовать наблюдаемому увеличению митохондриальной нагрузки и SRC.
Нет доказательств того, что ФАО способствует высокому SRC в клетках T
VMРанее считалось, что высокий SRC в клетках T MEM подпитывается окислением жирных кислот (FAO), механизмом, в значительной степени определенным с использованием этогооксира для ингибирования карнитин-пальмитоилтрансферазы I. (Cpt1), который является ферментом, ограничивающим скорость для FAO 3 .Однако недавно было продемонстрировано, что высокая концентрация этогооксира, использованная в этих исследованиях, также ингибировала другие компоненты OXPHOS, снижая SRC 26,27,28 . Чтобы изучить влияние этогооксира на высокий уровень SRC в клетках T VM , препарат был включен в тест Mito Stress с клетками T VM молодых мышей либо в высокой концентрации (200 мкМ), либо в низкой концентрации (5 мкМ). , последний из которых, как предполагается, сохраняет специфичность для Cpt1 (ref. 27 ).Существенное снижение OCR Max наблюдалось при высокой концентрации этомоксира, но было только очень умеренное снижение OCR Max при низкой концентрации (рис. 1h). Это предполагает, что FAO через Cpt1 не способствует высокому SRC, наблюдаемому в клетках T VM .
Затем мы исследовали возможность того, что гликолиз необходим для подпитки высокого SRC, наблюдаемого в клетках T VM , скорее всего, через производство пирувата 29 .Тест Mito Stress проводили на клетках T VM от наивных мышей с добавлением 2-дезокси-D-глюкозы (2-DG), аналога глюкозы, который ингибирует гликолиз. Добавление 2-DG имело минимальный эффект на OCR (дополнительный рис. 2a), аналогично добавлению низких доз этогооксира (рис. 1h). Напротив, добавление 2-DG резко снижает ECAR, подтверждая его эффективное ингибирование гликолиза (дополнительный рис. 2b). Эти данные предполагают, что высокий базальный SRC, наблюдаемый в клетках T VM , не зависит исключительно от FAO или гликолиза, но может подпитываться субстратом, генерируемым независимо от обоих путей.
Вирусная инфекция приводит к увеличению SRC в клетках T
VMВысокий SRC не наблюдался в клетках T MEM в этом исследовании (рис. 1a, b), но эти клетки T MEM были выделены из наивного SPF мышей и, вероятно, были созданы в ответ на комменсальные или низкопатогенные организмы и в условиях слабого воспаления. Чтобы оценить SRC в вызванных инфекцией клетках T MEM , мышей инфицировали вирусом гриппа A (IAV) и клетками T MEM , специфичными для тетрамерного H-2D b , нагруженных NP 366 , PA 224 и PB1-F2 62 эпитопов (клетки T MEM (IAV)) были выделены 20 дней спустя.Клетки T MEM и T MEM (IAV) показали одинаково низкий SRC (рис. 2a, b). Интересно, что клетки T MEM (IAV) постоянно демонстрировали значительно более высокие базальные и максимальные скорости внеклеточного закисления (ECAR Bas и ECAR Max ) по сравнению с клетками T N и T MEM (фиг. 2c). Эти данные показывают, что клетки T MEM и T MEM (IAV) метаболически различаются в отношении гликолитической, но не OXPHOS, способности.
Рис. 2: Клетки T VM увеличивают SRC при недавней инфекции.a OCR для отсортированных клеток T N и T MEM из селезенок неинфицированных молодых мышей SPF или клеток Tetramer + T MEM (IAV) от мышей, инфицированных IAV (20 дней после заражения) и b изменение OCR для каждого отсортированного подмножества ( n = 4–2, 3 экспериментальных повтора). c ECAR для отсортированных клеток T N и T MEM от молодых неинфицированных мышей или клеток Tetramer + T MEM (IAV) от мышей, инфицированных IAV (20 дней после заражения), с ECAR Bas и ECAR Max указаны ( n = 2–4, 3 экспериментальных повтора). d OCR для отсортированных клеток T N и T VM от молодых неинфицированных мышей или клеток T VM (IAV) от IAV-инфицированных мышей (20 дней после заражения) и e изменение OCR для каждой отсортированной подмножество ( n = 4–5, 3 экспериментальных повтора). f ECAR для отсортированных клеток T N и T VM от молодых неинфицированных мышей или мышей, инфицированных IAV (20 дней после заражения) ( n = 5, 3 экспериментальных повтора). Показано среднее значение ± SEM.NS указывает на незначительность, * указывает на p ≤ 0,05, ** указывает на p ≤ 0,01, непарный тест t .
Поразительно, недавняя инфекция IAV вызвала существенное повышение SRC клеток T VM (рис. 2d, e) без какого-либо изменения гликолитической способности (рис. 2f). Эти данные демонстрируют, что инфекция, как и старение, приводит к созданию среды, которая избирательно увеличивает SRC в клетках T VM (и, таким образом, антиген-независимым образом), и подтверждают, что высокий SRC не является каноническим признаком клеток T MEM . , даже вызванные инфекцией.
Традиционно определенные клетки T
CM представляют собой преимущественно клетки T VMНедавно было показано, что высокий SRC преимущественно разделяется с подмножеством долгоживущей центральной памяти (T CM ; CD44 hi CD62L hi ). T MEM , а не короткоживущей эффекторной памяти (T EM ; CD44 hi CD62L lo ) клеток 7 . В этом исследовании клетки T CM , по-видимому, были определены как CD44 hi CD62L hi CD8 + T-клетки, полученные от мышей после инфицирования вирусом острого лимфоцитарного хориоменингита (LCMV), которые будут включать клетки T VM . 20 .Чтобы определить степень, в которой метаболические характеристики клеток T CM были объединены с характеристиками клеток T VM , мы оценили долю классически определенных клеток T CM (CD44 hi CD62L hi ), которые были фактически клетки T VM (CD44 hi CD62L hi CD49d lo ) у наивных молодых, наивных старых мышей или мышей, инфицированных LCMV. У молодых и старых наивных мышей подавляющее большинство (≥85%) клеток T CM оказались клетками CD49d lo и, следовательно, клетками T VM (рис.3а). Даже через 40 дней после острой инфекции LCMV, которая вызывает значительный ответ Т-лимфоцитов CD8 + и создает устойчивые антиген-специфические популяции памяти 30 , более 60% клеток T CM оказались клетками T VM ( Рис. 3а). Это подчеркивает возможность того, что популяции Т-клеток CD8 + , ранее определенные как клетки T CM , от молодых, старых или инфицированных мышей, могли преимущественно состоять из клеток T VM .
Фиг.3: Клетки T VM составляют большую часть популяции клеток CD44 hi CD62L hi T CM .a Гистограммы экспрессии CD49d на CD44 hi CD62L hi CD8 + Т-клетки (T CM клетки) от наивных молодых мышей, наивных старых мышей и молодых мышей после заражения LCMV (через 40 дней после инфекция), с гистограммами, показывающими долю клеток T CM , которые являются клетками T VM ( n = 4–5). b Наложения CD44 hi CD49d lo T VM клеток от общего количества CD8 + Т-клеток с T EM / T CM гейтинг клеток (CD44 hi CD62L lo и CD44 привет CD62L привет соответственно). c Наложение CD44 hi CD49d hi T MEM клеток на общее количество CD8 + T-клеток. d Типичные точечные диаграммы, идентифицирующие IAV-специфический тетрамер + CD8 + Т-клетки, которые представляют собой CD62L hi (клетки T CM ) или CD62L lo (клетки T EM ) (60 дней после заражения). ), с гистограммами средней частоты тетрамера + CD8 + Т-клеток, которые находятся в каждой подгруппе ( n = 5). e OCR для отсортированных клеток T N и T VM от молодых неинфицированных мышей и отсортированных клеток T CM и T EM от мышей, инфицированных LCMV (60 дней после заражения), и изменение OCR f для каждого отсортированного подмножества ( n = 5–6, 3 экспериментальных повтора). Данные для a — d являются репрезентативными по крайней мере для 2 отдельных экспериментов. Показано среднее значение ± SEM. * обозначает p ≤ 0,05, ** обозначает p ≤ 0.01, непарный т проба.
Обратный анализ также был выполнен для определения распределения отсортированных клеток T VM (CD44 hi CD49d lo ) и T MEM (CD44 hi CD49d hi ) от наивных молодых, наивных пожилых или инфицированных LCMV мышей через классические ворота T CM (CD44 hi CD62L hi ) и эффекторную память (T EM ; CD44 hi CD62L lo ). Когда клетки T VM накладывались на графики CD44 / CD62L, они распределялись преимущественно по воротам T CM (~ 80%; рис.3б). Клетки T MEM преимущественно распределялись в воротах T EM (49-95%; рис. 3c), хотя значительная часть была обнаружена в воротах T CM у наивных молодых мышей. Мы также обнаружили, что клетки T MEM (IAV) были преимущественно клетками T EM (88,93 ± 6,67%) с некоторыми клетками T CM (11,07 ± 6,67%) (рис. 3d).
Чтобы окончательно определить относительный SRC клеток T CM и T EM наряду с клетками T VM , мы провели тест Mito Stress для морских коньков на клетках T N и T VM , выделенных от наивных мышей SPF и Клетки T EM и T CM , выделенные от мышей, инфицированных LCMV, через 60 дней после инфицирования.Как наблюдалось ранее, клетки T VM имели значительно более высокий SRC, чем все другие подмножества (рис. 3e, f). Кроме того, мы обнаружили, что клетки T EM имели значительно более низкий SRC, чем все другие подмножества, а клетки T CM имели умеренно более высокий SRC, чем клетки T N (рис. 3e, f). Это всесторонне демонстрирует, что высокий SRC не является определяющим признаком обычных ячеек T MEM , в частности подмножества ячеек T EM , а вместо этого является характеристикой ячеек T VM .
SRC коррелирует с маркерами выживаемости, а не функциональностью.
Высокий SRC был предложен в качестве характеристики клеток T MEM , способствующей как их расширенной функциональности, так и долгосрочному выживанию. 4,5 . Соответственно, мы стремились определить, коррелирует ли высокий SRC в клетках T VM и увеличение SRC с возрастом с улучшенной функциональностью CD8 + T-клеток или выживаемостью.
Три ключевых функции CD8 + Т-клеток во время иммунного ответа — это пролиферация, продукция цитокинов (особенно IFN-γ) и цитотоксичность.Недавно мы обнаружили, что клетки T VM от пожилых людей с самым высоким SRC обладают очень низкой TCR-управляемой пролиферативной способностью, хотя те немногие, которые могут отвечать, все еще продуцируют IFN-γ 17 . Чтобы дополнительно оценить функциональность субпопуляций CD8 + Т-клеток, оценивали их цитотоксическую способность непосредственно ex vivo. Отсортированные клетки T N , T VM и T MEM от молодых и старых мышей OT-I использовали в анализе цитотоксичности in vitro с нагруженными овальбумином спленоцитами в качестве мишеней.У молодых мышей клетки T VM были значительно более цитотоксичными, чем клетки T N , и эквивалентны клеткам T MEM (фиг. 4a). С возрастом цитотоксичность клеток T N оставалась низкой, в то время как цитотоксичность клеток T VM и T MEM существенно снижалась (рис. 4а). Эти данные, наряду с нашей предыдущей работой 17 , демонстрируют, что клетки T VM и T MEM снижают цитотоксическую и пролиферативную способность с возрастом, несмотря на их увеличение SRC.
Рис. 4: SRC коррелирует с экспрессией Bcl-2, но не с функциональностью Т-клеток CD8 + .a Процентный лизис OVA-нагруженных мишеней отсортированными клетками T N , T VM и T MEM от молодых или старых мышей OT-I ( n = 3–8). * Обозначает p ≤ 0,05, ** обозначает p ≤ 0,01, непарный тест t , данные являются репрезентативными по крайней мере для 3 отдельных экспериментов. Простой линейный регрессионный анализ среднего SRC из рис.1b против b — среднее число делений отсортированных Т-клеток CD8 + после 60-часовой стимуляции TCR из исх. 17 , c средний процент лизиса из ( a ), d средняя доля отсортированных CD8 + Т-клеток, продуцирующих IFN-γ через 36 часов после стимуляции TCR из ссылки. 17 и e средняя интенсивность флуоресценции (MFI) экспрессии Bcl-2 из ссылки. 17 .
Чтобы формально проверить, коррелирует ли способность любой функции CD8 + Т-клеток (пролиферация, продукция цитокинов или цитотоксичность) с SRC, был проведен линейный регрессионный анализ по типам клеток и возрасту.Не наблюдалось значительной корреляции ни для одной из функций (рис. 4b – d), что всесторонне демонстрирует, что высокий SRC не обязательно указывает на усиление функции Т-клеток CD8 + , особенно в Т-клетках CD8 + от пожилых людей.
Наша предыдущая работа показала, что клетки T VM от пожилых людей, которые демонстрируют самый высокий SRC, имели преимущество в выживаемости после адоптивного переноса 17 , который, по-видимому, был параллельным экспрессии антиапоптотического белка Bcl-2.Линейный регрессионный анализ экспрессии Bcl-2 по сравнению с SRC для разных типов клеток и возраста выявил значительную положительную корреляцию (рис. 4e), подчеркивая, что SRC связан с экспрессией Bcl-2 в качестве суррогата выживаемости в Т-клетках CD8 + .
Повышенный T
VM клетки передачи сигналов IL-15 увеличили SRCУчитывая сильную корреляцию Bcl-2 с SRC, были оценены медиаторы экспрессии Bcl-2 для определения драйверов высокого SRC в клетках T VM с возраст.Цитокин, IL-15 (обзор в ссылке 31 ), был сильным кандидатом, поскольку было показано, что он способствует выживанию Т-клеток с фенотипом памяти CD8 + за счет индукции фосфорилирования STAT5 (pSTAT5) и экспрессии Bcl- 2 (исх. 32 ). Чтобы понять, как передача сигналов IL-15 может изменяться с возрастом в различных подгруппах, мы оценили экспрессию субъединиц цитокиновых рецепторов и передачу сигналов ниже по течению в клетках T N , T VM и T MEM от молодых и старых мышей.
Экспрессия IL-15Rβ была низкой на клетках T N , умеренно выше на клетках T MEM и заметно и значительно выше на клетках T VM от молодых мышей (5-кратная, p <0,0001; Рис. 5а). Кроме того, в то время как ни T N , ни T MEM клетки не проявляли существенных возрастных изменений экспрессии, клетки T VM демонстрировали заметное увеличение экспрессии IL-15Rβ с возрастом (фиг. 5a). Когда каждое подмножество было отсортировано и стимулировано растворимым IL-15, интенсивность pSTAT5 отслеживалась с уровнями экспрессии рецептора, с клетками T VM от молодых мышей, демонстрирующих высокий pSTAT5, который еще больше увеличивался с возрастом (рис.5б). Вместе эти данные показывают, что из всех подмножеств Т-клеток CD8 + клетки Т VM проявляют наибольшую чувствительность к передаче сигналов IL-15, и эта чувствительность увеличивается с возрастом.
Рис. 5: Высокая чувствительность к IL-15 в клетках T VM увеличивается с возрастом и опосредует увеличение SRC.a IL-15Rβ MFI непосредственно ex vivo от отдельных мышей ( n = 5-7) или b pSTAT5 MFI через 15 минут стимуляции IL-15 in vitro на отсортированных T N , T Клетки VM и T MEM от молодых или старых мышей ( n = 3). c Репрезентативные точечные диаграммы для CD8 + Т-клеток, гейтированных по T N (CD44 lo ), T VM (CD44 hi CD49d lo ) и T MEM (CD44 hi ). CD49d hi ) клеток (указана частота CD8 + Т-клеток) и d частота CD8 + Т-клеток в каждой подгруппе мышей WT или IL-15 KO ( n = 5 для WT и 7 для КО). e OCR для отсортированных T N , T VM и T MEM клеток от молодых неинфицированных, IAV-инфицированных и IAV-инфицированных / нейтрализующих IL-15 мышей, обработанных mAb, и f изменение OCR для каждого отсортированного подмножество.Показано среднее значение ± SEM. * Указывает p ≤ 0,05, ** обозначает p ≤ 0,01, непарный тест t , данные для a , b , e , f являются репрезентативными по крайней мере для 3 отдельных экспериментов.
IL-15 считается критическим цитокином как для клеток T VM , так и для клеток T MEM , о чем свидетельствует тот факт, что мыши с нокаутом IL-15 (KO) теряют примерно половину своего фенотипа памяти (CD44 + ). ) клетки 33 и популяция клеток T VM не может развиваться у молодых мышей в отсутствие IL-15 или IL-15Rα 14,15 .Чтобы установить, отражает ли экспрессия IL-15Rβ и чувствительность к IL-15 субпопуляций Т-клеток CD8 + их относительную зависимость от IL-15, мы оценили развитие и устойчивость каждой подгруппы у молодых и старых мышей IL-15 KO. Клетки T VM отсутствовали у молодых мышей IL-15 KO, что согласуется с предыдущими исследованиями, а клетки T VM также отсутствовали у старых мышей (фиг. 5c, d). Следует отметить, что популяция клеток T MEM , по-видимому, остается относительно нетронутой у старых мышей (рис.5в, г). Это подчеркивает, что IL-15 абсолютно необходим для развития клеток T VM , но он незаменим для генерации и поддержания многих клеток T MEM у старых мышей.
IL-15 продуцируется на низких уровнях в устойчивом состоянии, но может резко повышаться в DC во время инфекции в ответ на передачу сигналов IFN типа I 34 . Чтобы проверить, приводит ли продукция IL-15 во время инфекции к наблюдаемому увеличению SRC в клетках T VM (рис.2d, e) мы вводили нейтрализующие IL-15 моноклональные антитела (mAb) молодым мышам, инфицированным IAV. MAb вводили на 0 и 3 дни после инфицирования, а SRC оценивали на 14 день. Прекращения передачи сигнала IL-15 во время инфекции было достаточно для отмены вызванного инфекцией увеличения SRC в клетках T VM (фиг. 5e, f). . В совокупности эти данные подчеркивают, что метаболический профиль и, в частности, повышенный уровень SRC, который ранее был связан с функцией и долголетием клеток T MEM , по-видимому, является прямой функцией воздействия IL-15 и не зависит от фенотипа T MEM .Кроме того, чувствительность клеток T VM к IL-15, по-видимому, объясняет высокий SRC, наблюдаемый в этих клетках непосредственно ex vivo от молодых мышей, и увеличение SRC в клетках T VM с возрастом.
Повышенный уровень IL-15Rβ и SRC в CD8 пожилого человека
+ Т-клеткиЧтобы определить, актуальны ли наши результаты на мышах для человека, мы сначала проанализировали экспрессию IL-15Rβ у молодых людей (20–30 лет) и старше. (60–80 лет) CD8 + Т-клетки человека. Мы обнаружили значительно более высокую экспрессию IL-15Rβ на клетках T VM по сравнению с клетками T N у молодых людей, и эта экспрессия значительно увеличивалась с возрастом во всех подгруппах (рис.6а, б), аналогично нашим наблюдениям на мышах. Более того, мы наблюдали тенденцию к увеличению SRC в общем количестве CD8 + Т-клеток с возрастом ( p = 0,1) (рис. 6c, d). Учитывая наше предыдущее описание сниженной пролиферативной способности Т-клеток CD8 + от пожилых людей 35 , эти данные предполагают аналогичное отсутствие корреляции между SRC и функциональностью в Т-клетках CD8 + человека. Более того, эти данные указывают на корреляцию между повышенным уровнем SRC и возрастным повышением чувствительности к IL-15 Т-клеток CD8 + человека.
Фиг. 6. Повышенная экспрессия IL-15Rβ и SRC в старых человеческих CD8 + Т-клетках.a IL-15Rβ MFI непосредственно ex vivo на субпопуляциях CD8 Т-клеток от отдельных молодых (20-30 лет) или старше (60-80 лет) взрослых доноров-людей ( n = 6-7) и b Репрезентативные гистограммы экспрессии IL-15Rβ с экспрессией на Т-клетках CD4 + , используемых в качестве контроля. c OCR для обогащенных CD8 + Т-клеток от молодых или пожилых людей-доноров и d изменение OCR для молодых или пожилых доноров-людей.Этот эксперимент проводился один раз. Столбцы или точки данных представляют собой среднее значение ± SEM. * Обозначает p ≤ 0,05, ** обозначает p ≤ 0,01, непарный тест t .
Метаболические особенности синдрома хронической усталости
Метаболические особенности синдрома хронической усталости — новаторская статья Naviaux Lab по метаболическому исследованию пациентов с синдромом хронической усталости (CFS), которая демонстрирует важные химические признаки крови. Он был опубликован в журнале Proceedings of the National Academy of Sciences of the Unites Stats of America (PNAS) 24 августа 2016 года.Эта статья имеет открытые данные и открытый доступ.
Синдром хронической усталости — это мультисистемное заболевание, которое вызывает длительную боль и инвалидность. Его сложно диагностировать из-за разрозненных симптомов и отсутствия диагностических лабораторных тестов. Мы сообщаем, что целенаправленная метаболомика плазмы широкого спектра не только выявила характерную химическую подпись, но и выявила неожиданную основную биологию. Метаболомика показала, что синдром хронической усталости представляет собой высоко согласованный гипометаболический ответ на стресс окружающей среды, который прослеживается до митохондрий и похож на классически изученное состояние развития Дауэра.Это открытие открывает новый путь для рациональной разработки новых терапевтических средств и определяет метаболомику как мощный инструмент для выявления химических различий, которые способствуют здоровью и болезням. [1]
Рональд Дэвис (редактор), Роберт Навио, Джейн К. Навио, Кефенг Ли, А. Тейлор Брайт, Уильям А. Алейник, Лин Ван, Аша Бакстер, Нил Натан, Уэйн Андерсон, Эрик Гордон
Ответы на тринадцать вопросов по
Метаболические особенности синдрома хронической усталости [править | править источник]РЕЗЮМЕ: Dr.Роберт Навьо отвечает на вопросы о своих выводах. Исходные вопросы и ответы: 1-6 [2] Обновленные вопросы и ответы: 1.1 и 7-12 [3]
- 1. СХУ — это реальное заболевание с химической сигнатурой крови и метаболическое, а не все в сознании
- 1.1 Глубокая личная потеря, горе, депрессия, страх, хроническая боль, тревога и посттравматическое стрессовое расстройство — все это вызывает химические изменения в крови, которые мы можем измерить с помощью метаболомики. Наука, стоящая за расширенным исследованием метаболомики CFS, требует, чтобы мы спросили как субъектов CFS, так и нормальных контролей о психологической травме, чтобы увидеть, может ли это увеличить восприимчивость к CFS в более позднем возрасте, и посмотреть, как предыдущая травма может повлиять на текущую метаболомику.
- 2. Реакция на опасность клетки (CDR) не выключается и приводит к «осадному метаболизму»
- 3. Люди не впадают в спячку, CFS имеет метаболическую подпись dauer
- 4. Это заболевание отличается от мужчин и женщин, не связанных с тестостероном или эстрогеном
- 5. Эпигенетика и пути метилирования ДНК, имеющие отношение к этому заболеванию
- 6. Этот документ не имеет отношения к лечению, хотя Metabolomics будет иметь отношение к любому клиническому испытанию CFS .
- 7.Доктор Навио согласен с доктором Дэвисом в том, что хроническое употребление антибиотиков может подавлять функцию митохондрий. Поэтому длительное использование антибиотиков без признаков активной инфекции приносит больше вреда, чем пользы.
- 8. Митохондрии выполняют две основные функции в клетке — энергетический метаболизм и клеточную защиту, и одна из них может быть сверхактивной за счет другой. Митохондрии не могут постоянно выполнять одновременно энергию и защиту (от простуды, гриппа и т. Д.) На 100%.
- 9. Означает ли тот факт, что некоторые антибиотики могут ингибировать митохондрии, что лечение болезни Лайма, которое длится слишком долго, может фактически преобразовать острую инфекцию Лайма в хронический постлаймский синдром и синдром хронической усталости? Да.
- 10. Существуют ли митококтейли или добавки, которые могут помочь, пока ученые не получат более точные методы лечения? Нет простого ответа. При употреблении митококтейлей могут возникать обострения и толчки систем организма у пациентов с СХУ. Начните с малого и продолжайте медленно, поскольку некоторые дозы B6 и магния могут вызвать учащенное сердцебиение.
- 11. Давая противовирусные препараты, врачи не только подавляют вирусы, но и подавляют многие метаболические функции клетки-хозяина. Иногда ингибирование функций клетки-хозяина может на время ослабить симптомы ME / CFS, но в других случаях использование сильнодействующих противовирусных препаратов подавляет митохондриальные реакции и реакции метилирования и может задержать полное выздоровление от ME / CFS.
- 12. Наши исследования показывают, что метаболизм может быть последним общим знаменателем для ME / CFS. Важно помнить, что «диета» и «метаболизм» — это не одно и то же. Диета — это то, что мы едим. Метаболизм — это функциональное состояние матрицы — динамическое состояние потока в сети, которое составляет все биохимические реакции, которые наши клетки используют для ведения жизненного дела.
Вирусы и CFS: Заявления Рона Дэвиса и Боба Навио [править | править источник]
Ниже приводится часть их заявлений.
Рон Дэвис: «Существует множество доказательств того, что различные вирусы могут инициировать ME / CFS, но менее ясно, что вирус участвует в поддержании болезни. Однако у некоторых пациентов может быть постоянное проблема с вирусами, особенно с вирусами, которые мы всегда носим, например, EBV и HHV6.
Эти вирусы обычно контролируются иммунной системой. Любое подавление иммунной системы может вызвать реактивацию этих вирусов (например,г., битумная черепица). Возможно (у нас есть новые подтверждающие данные), что иммунная система несколько ослаблена при ME / CFS, что затруднит подавление этих вирусов. Если мы сможем найти причину этой болезни и вылечить ее, проблема с вирусом должна исчезнуть ». [4]
Bob Naviaux: Вопрос — Многие эксперты ME / CFS улучшили симптомы у некоторых пациентов с помощью лечения противовирусными препаратами и амплигеном (двухцепочечная РНК polyIC).Я думаю, это доказывает, что наши симптомы вызывают продолжающиеся вирусные инфекции. Это не просто «уставшие пациенты», которые застряли в состоянии пониженного метаболизма из-за прошлых триггеров (которые сейчас исчезли).
«Прежде всего, важно, чтобы люди действительно читали нашу газету, прежде чем делать выводы из новостных сообщений и блогов и критиковать то, о чем мы никогда не говорили. Я видел ряд обобщений, которые начали появляться в блогах и репортажах журналистов в даже хорошие газеты и журналы, которые начинают отходить слишком далеко от реальной науки в нашей газете.
Мы посвятили часть статьи этому и другим связанным с ним вопросам об инфекциях. Название раздела было «Однородный метаболический ответ на гетерогенные триггеры». Он завершился предложением: «Несмотря на неоднородность триггеров, клеточная реакция на эти факторы стресса окружающей среды у пациентов, у которых развился СХУ, была однородной и статистически устойчивой». В качестве основы для этого вывода я рекомендую прочитать нашу статью по этой теме под названием «Метаболические особенности реакции клетки на опасность» (PMID 23981537).
Во-вторых, многие люди не понимают, что первая реакция нашего организма на вирусную, бактериальную или любую инфекцию — метаболическая. Да, наша химия — наша первая линия защиты. Наша химия отражает мгновенное состояние нашего здоровья. Врожденный иммунитет координируется митохондриями и является важным первым шагом в развитии адаптивного иммунитета к любому инфекционному агенту. Без врожденного иммунитета не может быть ни антител, ни активации NK-клеток, ни активации тучных клеток, ни иммунитета, опосредованного Т-клетками.
Кроме того, все противовирусные препараты обладают метаболическими эффектами, которые не имеют ничего общего с непосредственным подавлением синтеза вирусной ДНК или РНК. Многие противовирусные препараты ингибируют ключевой метаболический фермент SAdenosylhomocysteine Hydrolase (SAHH). Ингибирование SAHH вызывает повышение внутриклеточных уровней SAH. SAH — мощный ингибитор метилирования ДНК, РНК, белков и малых молекул. Это влияет на эпигенетику как вирусов, так и клеток-хозяев, экспрессию генов, трансляцию мРНК и стабильность белка.» [4]
Анализ
Метаболические особенности синдрома хронической усталости [править | править источник]Для тех, кто интересуется результатами и их последствиями, вкратце исследование Naviaux утверждает, что клетки пациентов с ME находятся в своего рода защитной спячке, ограничивая потребление ресурсов и переходя в гипометаболическое состояние в ответ на инфекция или другие факторы стресса. При детальном изучении метаболитов пациентов было обнаружено, что эта степень защитной гибернации напрямую коррелирует с клинической тяжестью.
Naviaux также утверждает, что клетки в ME / CFS — это клетки, находящиеся под огромным стрессом, для которого они создают серию защит, образно устанавливая превосходную систему блокировки и сигнализации и скрывая все ценности. Однако некоторые патогены знают код для входа, и когда ресурсы скрыты, хост также не может их использовать. Оба эти аспекта этого способа клеточной защиты имеют большое значение для симптоматики. [5]
Naviaux полагает, что митохондрии способны ощущать любую опасность — от патогенов до изменений pH до токсичных элементов от пестицидов, тяжелых металлов и т. Д.к воспалению. Они ощущают проблему в виде инфекции, когда обнаруживают падение напряжения, вызванное отвлечением электронов (НАДН / НАДФН) на выработку вирусных компонентов или реакцию на широкий спектр токсинов.
В ответе на опасность клетки (CDR) митохондрии мгновенно реагируют на эту потерю, уменьшая потребление кислорода, тем самым препятствуя патогенам использовать строительные блоки клетки для репликации. Поскольку кислород больше не используется, он накапливается в клетках, создавая окислительно-заряженную среду, которая прерывает вирусный синтез.CDR также укрепляет мембрану клетки, чтобы предотвратить выход патогенов из нее, и предупреждает другие клетки об опасности и испускает АТФ, чтобы предупредить другие клетки о необходимости повышения их защиты. [6]
Письмо: Ответ в VOGT и др. [Править | править источник]
СМИ и медицинские журналы [править | править источник]
- ↑ Naviaux, Robert K .; Naviaux, Jane C .; Ли, Кефэн; Брайт, А. Тейлор; Alaynick, William A .; Ван, Линь; Бакстер, Аша; Натан, Нил; Андерсон, Уэйн (25 августа 2016 г.).«Метаболические особенности синдрома хронической усталости». Труды Национальной академии наук : 201607571. DOI: 10.1073 / pnas.1607571113. ISSN 0027-8424. PMID 27573827.
- ↑ «Метаболические особенности синдрома хронической усталости» (PDF). naviauxlab.ucsd.edu . 31 августа 2016г.
- ↑ «ОБНОВЛЕНО: метаболические особенности синдрома хронической усталости — вопросы и ответы с Робертом Навио, доктором медицины | Фонд открытой медицины». Фонд открытой медицины .9 сентября, 2016. Проверено 19 августа, 2018.
- ↑ 4,0 4,1 «Вирусы и CFS: заявления Рона Дэвиса и Боба Навио | Фонд открытой медицины». Фонд открытой медицины . 9 сентября, 2016. Проверено 19 августа, 2018.
- ↑ 5,0 5,1 Зельцер, Хайме (30 августа 2016 г.). «Газета Naviaux о метаболизме настолько обширна, насколько вы думаете — #MEAction». # ИЗМЕРЕНИЕ . Проверено 19 августа 2018 года.
- ↑ 6,0 6,1 Джонсон, Корт (1 сентября 2016 г.). «Выявлена основная проблема синдрома хронической усталости? Исследование Метаболомики Навьё открывает новые горизонты — здоровье растет». Здоровье . Проверено 19 августа 2018. .
- ↑ Naviaux, Robert K .; Naviaux, Jane C .; Ли, Кефэн; Брайт, Тейлор А .; Alaynick, William A .; Ван, Линь; Бакстер, Аша; Натан, Нил; Андерсон, Уэйн (2 ноября 2016 г.). «Ответ на VOGT ET AL: Метаболомика и синдром хронической усталости» (PDF). pnas.org . Гордон, Эрик.
Информация, представленная на этом сайте, не предназначена для диагностики или лечения каких-либо заболеваний.
Из MEpedia, краудсорсинговой энциклопедии науки и истории ME и CFS.
Что такое метаболизм?
Метаболизм — это термин, который используется для описания всех химических реакций, участвующих в поддержании жизненного состояния клеток и организма. Обмен веществ можно условно разделить на две категории:
- Катаболизм — распад молекул для получения энергии
- Анаболизм — синтез всех соединений, необходимых клеткам
Метаболизм тесно связан с питанием и доступностью питательных веществ.Биоэнергетика — это термин, который описывает биохимические или метаболические пути, с помощью которых клетка в конечном итоге получает энергию. Формирование энергии — один из жизненно важных компонентов обмена веществ.
Изображение предоставлено: VectorMine / Shutterstock.com
Питание, обмен веществ и энергия
Питание — это ключ к обмену веществ. Пути метаболизма зависят от питательных веществ, которые они расщепляют, чтобы произвести энергию. Эта энергия, в свою очередь, требуется организму для синтеза таких молекул, как новые белки и нуклеиновые кислоты (ДНК, РНК).
Питательные вещества, связанные с метаболизмом, включают такие факторы, как потребности организма в различных веществах, индивидуальные функции в организме, необходимое количество и уровень, ниже которого ухудшается состояние здоровья.
Основные питательные вещества поставляют энергию (калории) и поставляют необходимые химические вещества, которые сам организм не может синтезировать. Пища содержит множество веществ, которые необходимы для построения, содержания и восстановления тканей тела, а также для его эффективного функционирования.
Диета нуждается в основных питательных веществах, таких как углерод, водород, кислород, азот, фосфор, сера и около 20 других неорганических элементов. Основные элементы представлены углеводами, липидами и белком. Кроме того, необходимы витамины, минералы и вода.
Углеводы в обмене веществ
Продукты питания содержат углеводы в трех формах: крахмал, сахар и целлюлозу (клетчатку). Крахмал и сахар являются основными и необходимыми источниками энергии для человека. Волокна увеличивают объем рациона.
Ткани организма зависят от глюкозы во всех сферах деятельности. Углеводы и сахара производят глюкозу в результате пищеварения или метаболизма.
Общая реакция горения глюкозы записывается как:
C 6 H 12 O 6 + 6 O 2 ——> 6 CO 2 + 6 H 2 O + энергия
Большинство людей потребляют около половины своего рациона в виде углеводов. Это происходит из таких продуктов, как рис, пшеница, хлеб, картофель и макаронные изделия.
Белки в обмене веществ
Белки являются основными строителями тканей в организме. Они являются частью каждой клетки тела. Белки помогают в структуре клеток, функциях, формировании гемоглобина для переноса кислорода, ферментах для выполнения жизненно важных реакций и множестве других функций в организме. Белки также жизненно важны для снабжения азотом генетического материала ДНК и РНК и производства энергии.
Белки необходимы для питания, поскольку содержат аминокислоты. Из 20 или более аминокислот человеческий организм не может синтезировать 8, и они называются незаменимыми аминокислотами.
Незаменимые аминокислоты включают:
- Лизин
- Триптофан
- метионин
- лейцин
- Изолейцин
- Фенилаланин
- Валин
- Треонин
Продукты с высоким содержанием белка: яйца, молоко, соевые бобы, мясо, овощи и зерновые.
Жир в обмене веществ
Жиры — это концентрированные источники энергии. Они производят в два раза больше энергии, чем углеводы или белки, в пересчете на вес.
Функции жиров включают:
- Помогает формировать клеточную структуру;
- Образует защитную подушку и изоляцию вокруг жизненно важных органов;
- Помогает усвоить жирорастворимые витамины,
- Обеспечение резервного хранилища энергии
Незаменимые жирные кислоты включают ненасыщенные жирные кислоты, такие как линолевая, линоленовая и арахидоновая кислоты. Их нужно принимать с пищей. Насыщенные жиры, наряду с холестерином, участвуют в артериосклерозе и сердечных заболеваниях.
Минералы и витамины в обмене веществ
Минералы, содержащиеся в пищевых продуктах, не вносят прямого вклада в энергетические потребности, но важны как регуляторы организма и играют роль в метаболических путях организма. В организме человека содержится более 50 элементов. Было обнаружено, что незаменимыми являются около 25 элементов, а это означает, что их дефицит вызывает определенные симптомы дефицита.
Важные минералы включают:
- Кальций
- фосфор
- Утюг
- Натрий
- Калий
- Хлорид-ионы
- Медь
- Кобальт
- Марганец
- цинк
- Магний
- фтор
- Йод
Витамины — это важные органические соединения, которые человеческий организм не может синтезировать сам по себе, и поэтому они должны присутствовать в рационе.Витамины, особенно важные для обмена веществ, включают:
- Витамин А
- B2 (рибофлавин)
- Ниацин или никотиновая кислота
- Пантотеновая кислота
Кредит изображения: Siberian Art / Shutterstock.com
Метаболические пути
Химические реакции метаболизма организованы в метаболические пути. Они позволяют преобразовать основные химические вещества из пищевых продуктов с помощью последовательности ферментов через ряд этапов в другое химическое вещество.
Ферменты имеют решающее значение для метаболизма, потому что они позволяют организмам вызывать желательные реакции, требующие энергии. Эти реакции также связаны с реакциями, высвобождающими энергию. Поскольку ферменты действуют как катализаторы, они позволяют этим реакциям протекать быстро и эффективно. Ферменты также позволяют регулировать метаболические пути в ответ на изменения в клеточной среде или сигналы от других клеток.
Список литературы
Дополнительная литература
Изучение тканеспецифической метаболической физиологии человека
Часто бывает сложно провести на людях достаточно специфические исследования метаболической физиологии, чтобы получить информацию о функциях органа или ткани.Например, поскольку переход от состояния натощак к состоянию сытости отражает ключевые адаптации для поддержания жизни, может произойти существенное изменение потока метаболических субстратов. Действительно, в некоторых органах может происходить полный сдвиг в предпочтении метаболического субстрата. Таким образом, изучение метаболизма органов / тканей человека в голодном состоянии может продемонстрировать только один аспект метаболической физиологии.
Здесь мы описываем методы, которые мы использовали для улучшения понимания тканеспецифического метаболизма человека, уделяя особое внимание жировым депо.
АРТЕРИО-ВЕНОЗНАЯ (A-V) ПРОБКА КРОВИ
Это классический метод выделения органа как метаболической системы. Он может быть очень информативным для определенных органов, но его трудно применить к системам, в которых отбор проб затруднен или когда орган имеет клеточную или метаболическую неоднородность. Техника A-V основана на принципе Фика и использовалась для исследования ноги и предплечья (приближения к скелетным мышцам) 1–3 и сердца, 4 с взятием образцов из печеночной вены, используемым для исследования печени.5,6 Образцы нижней эпигастральной вены и подкожной вены использовались для исследования подкожной брюшной и бедренной жировой ткани соответственно. 2,3,7–10
«Быстрый прогресс в визуализации обеспечивает менее инвазивные методы для тканеспецифичных исследований метаболической физиологии»
В тех случаях, когда невозможно получить артериальную кровь, использовался метод артериализации вен (нагретая рука). В этом методе необходимо учитывать потенциальные эффекты температуры тела и кровотока в коже, и он может не подходить при изучении потребления кислорода тканями. 11,12 Положение венозных катетеров также требует тщательного рассмотрения, так как некоторые вены, например бедренная вена, получают кровь от мышц ног, а также от жировой ткани и кожи. 12
Техника разности A-V в сочетании с измерением кровотока в интересующем органе / ткани позволяет измерять чистый поток вещества, а также клиренс вещества тканью. Мы использовали методологию различия A-V с селективной венозной катетеризацией подкожной жировой ткани брюшной полости и скелетных мышц предплечья в исследованиях со стабильными изотопами для описания метаболических характеристик этих тканей после ночного голодания и после смешанного пробного завтрака. 3 Хиломикрон (диетический / экзогенный) и липопротеин очень низкой плотности (ЛПОНП; эндогенный) триацилглицерин (ТГ) были очищены через жировую ткань и мышцы, причем фракционная экстракция хиломикрон-ТГ была значительно выше, чем у ЛПОНП-ТГ. . 3
Используя разницу A-V в подкожной жировой ткани брюшной полости и кормление смешанными порциями с интервалом в 5 часов, мы обнаружили, что жировая ткань худых мужчин была более метаболически активна в течение 24-часового периода, чем у субъектов с абдоминальным ожирением. 13 В частности, мужчины с абдоминальным ожирением накапливают значительно меньшую долю диетического жира в подкожной жировой ткани брюшной полости, чем худые мужчины, что может увеличить количество жира, хранящегося в печени и скелетных мышцах. 13
Мы также использовали методику A-V для описания метаболических характеристик подкожной брюшной и ягодично-бедренной жировой ткани при голодании и после приема пищи. 14 и после адренергической стимуляции. 15 Мы обнаружили явные метаболические различия в потоке жирных кислот и кровотоке между жировыми депо, при этом ягодично-бедренные депо более метаболически неактивны по сравнению с абдоминальными. 14,15
ИСПОЛЬЗОВАНИЕ МЕТАБОЛИЧЕСКИХ МЕТЕЛЕЙ
Метаболические индикаторы (стабильные и радиоизотопы) дают возможность специально изучать метаболизм тканей или исследовать определенные метаболические пути. Факторы, которые следует учитывать, включают способ доставки, естественное содержание индикатора, метаболическое обращение с индикатором, маркируемые молекулы, фоновые эффекты приема пищи и время между повторными посещениями для исследования.
СПОСОБЫ ИЗОБРАЖЕНИЯ
Быстрый прогресс в области визуализации обеспечивает создание менее инвазивных методов для тканеспецифических исследований метаболической физиологии.Методы метаболической визуализации включают позитронно-эмиссионную томографию (ПЭТ), магнитно-резонансную томографию / спектроскопию (MRI / S), компьютерную томографию (CT) и ультразвук (US). Наряду с оценкой метаболизма субстрата, методы визуализации могут измерять перфузию органов (ПЭТ), плотность ткани и тип жировой ткани (КТ), эктопическое отложение жира, содержание метаболитов и уровни оксигенации крови в головном мозге (МРТ / С) и жесткость печени. / жир (США). 16
Измерители были использованы в комбинации ПЭТ и МРТ / С для исследования метаболизма жирных кислот и глюкозы in vivo у людей.Например, при кормлении людей едой, содержащей 13C-жирные кислоты, и последующем измерении сигнала 13C в печени с помощью МРТ / S через определенные промежутки времени после приема пищи было обнаружено, что поток пищевых жирных кислот через печень заметно медленнее у людей с высоким по сравнению с с низким содержанием жира в печени. 17
Гиперполяризованная МРТ (HP MRI) — это новая технология, позволяющая обнаруживать молекулы, обогащенные 13C, со значительно увеличенным сигналом по сравнению с традиционными методами.HP MRI позволит отслеживать единичные и множественные метаболические пути с использованием одиночных или множественных гиперполяризованных зондов, наряду с отслеживанием превращения субстрата в продукты его метаболизма в реальном времени, 18 , но технические платформы дороги, а опыт работы с людьми ограничен .
ПЭТ — это альтернативный неинвазивный подход к визуализации, который использовался для исследования метаболизма органов или тканей. Обычно это делается в состоянии голодания.При использовании ПЭТ и 11С-пальмитата у субъектов с ожирением было показано повышенное окисление жирных кислот печени без разницы в уровнях поглощения и этерификации жирных кислот печени по сравнению с контрольной группой, не страдающей ожирением, в состоянии натощак. 19
Совсем недавно метаболический зонд 18F-FTHA и ПЭТ использовался для изучения метаболических изменений в жировой ткани у лиц с ожирением до и после потери веса. 20 После потери веса не было изменений в поглощении неэтерифицированных жирных кислот висцеральной и подкожной жировой тканью брюшной полости, но поглощение жировой тканью бедренной кости значительно снизилось.
Короткий период полураспада радиоизотопов, подходящих для ПЭТ, ограничивает этот метод изучением метаболических путей с короткими временными рамками, таких как поглощение глюкозы или жирных кислот. Метаболические превращения, такие как гликолиз или окисление жирных кислот, не могут быть изучены. Последнее соображение заключается в том, что ПЭТ может обнаруживать включение радиоактивно меченых индикаторов в ткань, но не может отличить исходное соединение от образованного радиоактивно меченного продукта. 18
Таким образом, исследования тканеспецифической метаболической функции классически основывались на сложных методах баланса A-V, предпочтительно с использованием меченых метаболических индикаторов.Однако новые методы визуализации метаболизма открывают новые возможности для таких исследований на людях.
Линн Ходсон и Фредрик Карпе
Оксфордский центр диабета, эндокринологии и метаболизма, Оксфордский университет
ССЫЛКИ
- Olsen DB et al. 2005 Физиологический журнал 565 555–562.
- Coppack SW et al. 1990 Метаболизм 39 384–390.
- Bickerton AS et al. 2007 Диабет 56 168–176.
- Funada J et al. 2009 PLoS One 4 e7533.
- Havel RJ et al. 1970 Журнал клинических исследований 49 2017–2035.
- Westerbacka J et al. 2010 Гастроэнтерология 139 1961–1971 e1.
- Фрейн К.Н. и Коппак С.В. 2001 Методы молекулярной биологии 155 269–279.
- Frayn KN et al. 1993 Международный журнал ожирения и связанных с ним метаболических нарушений 17 Дополнение 3 S18 – S21, S22.
- Frayn KN et al. 1994 Американский журнал физиологии 226 E308 – E317.
- McQuaid SE et al. 2010 Ожирение (Сильвер-Спринг) 18 1055–1058.
- Фрейн К.Н. 1999 Труды Общества питания 58 877–886.
- Macdonald IA 1999 Proceedings of the Nutrition Society 58 871–875.
- McQuaid SE et al. 2011 Диабет 60 47–55.
- McQuaid SE et al. 2010 Диабет 59 2465–2473.
- Manolopoulos KN et al. 2012 Диабетология 55 3029–3037.
- Iozzo P 2015 Анналы Нью-Йоркской академии наук 1353 21–40.
- Ravikumar B et al. 2005 Американский журнал физиологии — эндокринология и метаболизм 288 E789 – E797.
- Di Gialleonardo V et al. 2016 Семинары по ядерной медицине 46 28–39.
- Iozzo P et al. 2010 Гастроэнтерология 139 846–856, 856.e1–856.e6.
- Bucci M et al. 2015 Диабетология 58 158–164.
Какие физические данные характерны для метаболического синдрома?
Олуфади Р., Бирн CD. Клинико-лабораторная диагностика метаболического синдрома. Дж. Клин Патол . 2008 июнь 61 (6): 697-706. [Медлайн].
Эрнандес-Байшаули Дж., Кесада-Васкес С., Марин-Касадо Р. и др. Выявление ранних факторов риска заболеваний, связанных с метаболическим синдромом: новая эра с оценкой метаболомики ЯМР. Питательные вещества .2020 18 марта. 12 (3): [Medline]. [Полный текст].
[Рекомендации] Alberti KG, Eckel RH, Grundy SM, et al. Гармонизация метаболического синдрома: совместное промежуточное заявление Целевой группы Международной федерации диабета по эпидемиологии и профилактике; Национальный институт сердца, легких и крови; Американская Ассоциация Сердца; Всемирная федерация сердца; Международное общество атеросклероза; и Международная ассоциация изучения ожирения. Тираж . 2009 20 октября.120 (16): 1640-5. [Медлайн].
Tasali E, IP MS. Обструктивное апноэ сна и метаболический синдром: изменения метаболизма глюкозы и воспаление. Proc Am Thorac Soc . 2008 15 февраля. 5 (2): 207-17. [Медлайн].
Йонеда М., Ямане К., Джитсуики К. и др. Распространенность метаболического синдрома у коренных японцев и американцев японского происхождения. Диабет Рес, Клиническая Практика . 2008 Март 79 (3): 518-22. [Медлайн].
Гранди С.М., Климан Дж. И., Дэниэлс С. Р. и др.Диагностика и лечение метаболического синдрома: научное заявление Американской кардиологической ассоциации / Национального института сердца, легких и крови. Тираж . 2005, 25 октября. 112 (17): 2735-52. [Медлайн].
Хэнли А.Дж., Картер А.Дж., Уильямс К. и др. Прогнозирование сахарного диабета 2 типа с альтернативными определениями метаболического синдрома: Исследование инсулинорезистентного атеросклероза. Тираж . 2005 декабрь 13, 112 (24): 3713-21. [Медлайн].
Gami AS, Witt BJ, Howard DE, et al.Метаболический синдром и риск сердечно-сосудистых событий и смерти: систематический обзор и метаанализ продольных исследований. Джам Колл Кардиол . 2007 30 января. 49 (4): 403-14. [Медлайн].
Джованнуччи Э. Метаболический синдром, гиперинсулинемия и рак толстой кишки: обзор. Am J Clin Nutr . 2007 сентябрь 86 (3): s836-42. [Медлайн].
Xu H, Li X, Adams H, Kubena K, Guo S. Этиология метаболического синдрома и диетическое вмешательство. Int J Mol Sci . 2018 31 декабря 20 (1): [Medline]. [Полный текст].
Sattar N, McConnachie A, Shaper AG и др. Может ли метаболический синдром предсказать сердечно-сосудистые заболевания и диабет? Данные результатов двух проспективных исследований. Ланцет . 7 июня 2008 г. 371 (9628): 1927-35. [Медлайн].
Нильссон П.М. Сердечно-сосудистый риск при метаболическом синдроме: факт или вымысел ?. Curr Cardiol Rep . 2007 ноября, 9 (6): 479-85.[Медлайн].
Гранди С.М. Метаболический синдром: фактор риска множественных сердечно-сосудистых заболеваний. J Clin Endocrinol Metab . 2007 февраль 92 (2): 399-404. [Медлайн].
Cuspidi C, Sala C, Zanchetti A. Метаболический синдром и поражение органов-мишеней: роль артериального давления. Эксперт Rev Cardiovasc Ther . 2008 июн. 6 (5): 731-43. [Медлайн].
Serne EH, de Jongh RT, Eringa EC, et al. Микрососудистая дисфункция: потенциальная патофизиологическая роль в метаболическом синдроме. Гипертония . 2007 июл.50 (1): 204-11. [Медлайн].
Алесси М.С., Юхан-Ваг I. Метаболический синдром, гемостаз и тромбоз. Тромб Хемост . 2008 июнь 99 (6): 995-1000. [Медлайн].
Stehouwer CD, Генри Р.М., Феррейра И. Артериальная жесткость при диабете и метаболическом синдроме: путь к сердечно-сосудистым заболеваниям. Диабетология . 2008 апр. 51 (4): 527-39. [Медлайн].
Элнакиш М.Т., Хассанаин Х.Х., Янссен П.М., Ангелос М.Г., Хан М.Возникающая роль окислительного стресса в метаболическом синдроме и сердечно-сосудистых заболеваниях: важная роль Rac / NADPH-оксидазы. Дж. Патол . 2013 ноябрь 231 (3): 290-300. [Медлайн].
Юберо-Серрано Е.М., Дельгадо-Листа Дж., Пена-Ориуэла П. и др. Окислительный стресс связан с рядом компонентов метаболического синдрома: исследование LIPGENE. Exp Mol Med . 2013 21 июня. 45: e28. [Медлайн]. [Полный текст].
Ogden CL, Carroll MD, Fryar CD, Flegal KM.Распространенность ожирения среди взрослых и молодежи: США, 2011-2014 гг. Краткий обзор данных NCHS . 2015 1-8 ноября. [Медлайн].
Саклаен МГ. Глобальная эпидемия метаболического синдрома. Curr Hypertens Rep . 2018 26 февраля. 20 (2): 12. [Медлайн].
Goossens GH. Роль дисфункции жировой ткани в патогенезе инсулинорезистентности, связанной с ожирением. Physiol Behav . 2008 г. 23 мая. 94 (2): 206-18. [Медлайн].
Густафсон Б., Хаммарштедт А., Андерссон С.Х. и др. Воспаление жировой ткани: причина метаболического синдрома и атеросклероза. Артериосклер тромб Vasc Biol . 2007 27 ноября (11): 2276-83. [Медлайн].
Ланн Д., Леройт Д. Инсулинорезистентность как основная причина метаболического синдрома. Med Clin North Am . 2007 ноябрь 91 (6): 1063-77, viii. [Медлайн].
Despres JP, Lemieux I, Bergeron J, et al.Абдоминальное ожирение и метаболический синдром: вклад в глобальный кардиометаболический риск. Артериосклер тромб Vasc Biol . 2008 июн.28 (6): 1039-49. [Медлайн].
Туркоглу С., Думан Б.С., Гунай Д., Чагатай П., Озджан Р., Буюкдеврим А.С. Влияние абдоминального ожирения на инсулинорезистентность и компоненты метаболического синдрома: данные, подтверждающие, что ожирение является центральным признаком. Obes Surg . 13 октября 2003 г. (5): 699-705. [Медлайн].
Гольдбахер Э.М., Мэтьюз К.А.Связаны ли психологические характеристики с риском метаболического синдрома? Обзор литературы. Энн Бихав Мед . 2007 ноябрь-декабрь. 34 (3): 240-52. [Медлайн].
Сардинья А, Нарди АЕ. Роль тревоги в метаболическом синдроме. Medscape. Доступно на http://www.medscape.com/viewarticle/755474_4. Доступ: 4 декабря 2013 г.
Гранди С.М. Пандемия метаболического синдрома. Артериосклер тромб Vasc Biol . 2008 апр.28 (4): 629-36. [Медлайн].
Ford ES, Giles WH, Mokdad AH. Растущая распространенность метаболического синдрома среди взрослых в США. Уход за диабетом . 2004, 27 октября (10): 2444-9. [Медлайн].
Mozaffarian D, Benjamin EJ, Go AS, et al. Обновление статистики сердечных заболеваний и инсульта за 2016 г .: отчет Американской кардиологической ассоциации. Тираж . 2015 16 декабря. 133: [Medline]. [Полный текст].
Lovre D, Mauvais-Jarvis F.Тенденции распространенности метаболического синдрома. JAMA . 2015 г., 13 сентября. 314 (9): 950. [Медлайн].
Hoang KC, Le TV, Wong ND. Метаболический синдром у жителей Восточной Азии. J Cardiometab Syndr . Осень 2007 года. 2 (4): 276-82. [Медлайн].
Hwang LC, Bai CH, Chen CJ. Распространенность ожирения и метаболического синдрома на Тайване. J Formos Med Assoc . 2006 августа 105 (8): 626-35. [Медлайн].
Nestel P, Lyu R, Low LP, et al.Метаболический синдром: недавняя распространенность среди населения Восточной и Юго-Восточной Азии. Азия Пак Дж. Клин Нутр . 2007. 16 (2): 362-7. [Медлайн].
Kohro T., Furui Y, Mitsutake N, et al. Японская национальная программа проверки здоровья и вмешательства, направленная на предотвращение обострения метаболического синдрома. Инт Сердце J . 2008 Март 49 (2): 193-203. [Медлайн].
Коловоу Г.Д., Анагностопулу К.К., Салпеа К.Д. и др. Распространенность метаболического синдрома в различных популяциях. Am J Med Sci . 2007 июн. 333 (6): 362-71. [Медлайн].
Hu G, Lindstrom J, Jousilahti P, et al. Растущая распространенность метаболического синдрома среди финских мужчин и женщин за последние десять лет. J Clin Endocrinol Metab . 2008 Март 93 (3): 832-6. [Медлайн].
Erem C, Hacihasanoglu A, Deger O, et al. Распространенность метаболического синдрома и связанных с ним факторов риска среди взрослого населения Турции: исследование Trabzon MetS. Эндокринная .2008 Февраль 33 (1): 9-20. [Медлайн].
Махадик SR, Deo SS, Mehtalia SD. Повышенная распространенность метаболического синдрома у азиатских индийцев, не страдающих ожирением — сравнение между городом и деревней. Расстройство отношений синдрома метаболизма . 2007 июн. 5 (2): 142-52. [Медлайн].
Мокан М, Галайда П, Придавкова Д и др. Распространенность сахарного диабета и метаболического синдрома в Словакии. Диабет Рес, Клиническая Практика . 2008 августа 81 (2): 238-42. [Медлайн].
Малик М, Разиг С.А.Распространенность метаболического синдрома среди многонационального населения Объединенных Арабских Эмиратов: отчет национального опроса. Расстройство отношений синдрома метаболизма . Осень 2008 г. 6 (3): 177-86. [Медлайн].
Банерджи Д., Мисра А. Повышает ли польза термина «метаболический синдром» использование этнических критериев? Споры и предложения. Инт Дж. Обес (Лондон) . 2007 сентября 31 (9): 1340-9. [Медлайн].
Кларк LT, Эль-Атат Ф.Метаболический синдром у афроамериканцев: значение для предотвращения ишемической болезни сердца. Клин Кардиол . 2007 Апрель 30 (4): 161-4. [Медлайн].
Ford ES, Giles WH, Dietz WH. Распространенность метаболического синдрома среди взрослых в США: результаты третьего Национального исследования здоровья и питания. JAMA . 2002 16 января. 287 (3): 356-9. [Медлайн].
Ukegbu UJ, Castillo DC, Knight MG, et al. Метаболический синдром не обнаруживает метаболического риска у африканских мужчин, живущих в США.С. Уход за диабетом . 2011 Октябрь 34 (10): 2297-9. [Медлайн]. [Полный текст].
Bentley-Lewis R, Koruda K, Seely EW. Метаболический синдром у женщин. Нат Клин Практик Метаб эндокринола . 2007 октября 3 (10): 696-704. [Медлайн].
Essah PA, Wickham EP, Nestler JE. Метаболический синдром при синдроме поликистозных яичников. Clin Obstet Gynecol . 2007 марта 50 (1): 205-25. [Медлайн].
Кассонс А.Дж., Стаки Б.Г., Уоттс Г.Ф.Метаболический синдром и кардиометаболический риск при СПКЯ. Curr Diab Rep . 2007 февраля, 7 (1): 66-73. [Медлайн].
Сюэ Ф, Михельс КБ. Диабет, метаболический синдром и рак груди: обзор имеющихся данных. Am J Clin Nutr . 2007 сентябрь 86 (3): s823-35. [Медлайн].
Рамос Р.Г., Олден К. Распространенность метаболического синдрома среди американских женщин детородного возраста. Am J Public Health . 2008 июнь 98 (6): 1122-7.[Медлайн].
Bhasin S, Jasjua GK, Pencina M, et al. Глобулин, связывающий половые гормоны, но не тестостерон, проспективно и независимо связан с возникающим метаболическим синдромом у мужчин: исследование сердца Фрамингема. Уход за диабетом . 2011 ноябрь 34 (11): 2464-70. [Медлайн]. [Полный текст].
Tsujimura A, Miyagawa Y, Takezawa K, et al. Является ли низкая концентрация тестостерона фактором риска метаболического синдрома у здоровых мужчин среднего возраста? Урология . 2013 Октябрь 82 (4): 814-9. [Медлайн].
De Ferranti SD, Osganian SK. Эпидемиология детского метаболического синдрома и сахарного диабета 2 типа. Diab Vasc Dis Res . 2007 декабрь 4 (4): 285-96. [Медлайн].
Zimmet P, Alberti KG, Kaufman F, et al. Метаболический синдром у детей и подростков — консенсус IDF. Детский диабет . 2007 октября 8 (5): 299-306. [Медлайн].
Моррисон Дж. А., Форд Э. С., Штейнбергер Дж.Детский метаболический синдром. Минерва Мед . 2008 июнь 99 (3): 269-87. [Медлайн].
Уметани К., Кодама Ю., Накамура Т. и др. Высокая распространенность пароксизмальной фибрилляции предсердий и / или трепетания предсердий при метаболическом синдроме. Циркуляр J . 2007 февраль 71 (2): 252-5. [Медлайн].
Watanabe H, Tanabe N, Watanabe T., Darbar D, Roden DM, Sasaki S. Метаболический синдром и риск развития фибрилляции предсердий: исследование профилактической медицины Ниигата. Тираж . 2008 11 марта. 117 (10): 1255-60. [Медлайн].
Марон Б.А., Леопольд Дж. А., Хемнес, АР. Метаболический синдром, нейрогуморальная модуляция и легочная артериальная гипертензия. Br J Pharmacol . 2020 Апрель 177 (7): 1457-71. [Медлайн]. [Полный текст].
Briand M, Lemieux I, Dumesnil JG, et al. Метаболический синдром отрицательно влияет на прогрессирование заболевания и прогноз при стенозе аорты. Джам Колл Кардиол .6 июня 2006 г. 47 (11): 2229-36. [Медлайн].
Обунай К., Яни С., Дангас Г.Д. Сердечно-сосудистая заболеваемость и смертность от метаболического синдрома. Med Clin North Am . 2007 ноябрь 91 (6): 1169-84, x. [Медлайн].
Mraovic B, Hipszer BR, Epstein RH, et al. Метаболический синдром увеличивает риск тромбоэмболии легочной артерии после артропластики тазобедренного и коленного суставов. Croat Med J . 2013 Август 54 (4): 355-61. [Медлайн].
Towfighi A, Ovbiagele B.Метаболический синдром и инсульт. Curr Diab Rep . 2008 Февраль 8 (1): 37-41. [Медлайн].
Air EL, Kissela BM. Диабет, метаболический синдром и ишемический инсульт: эпидемиология и возможные механизмы. Уход за диабетом . 2007 декабря 30 (12): 3131-40. [Медлайн].
Каллаган Б., Фельдман Э. Метаболический синдром и невропатия: терапевтические проблемы и возможности. Энн Нейрол . 2013 Сентябрь 74 (3): 397-403. [Медлайн].[Полный текст].
Лидофский СД. Неалкогольная жировая болезнь печени: диагностика и связь с метаболическим синдромом и подходы к лечению. Curr Diab Rep . 2008 г. 8 (1): 25-30. [Медлайн].
Котронен А., Ики-Ярвинен Х. Жирная печень: новый компонент метаболического синдрома. Артериосклер тромб Vasc Biol . 2008, 28 января (1): 27-38. [Медлайн].
Tarantino G, Finelli C. Как насчет неалкогольной жировой болезни печени как нового критерия для определения метаболического синдрома ?. Мир Дж. Гастроэнтерол . 2013 14 июня. 19 (22): 3375-84. [Медлайн]. [Полный текст].
Beaulieu LM, Whitley BR, Wiesner TF, et al. Рак груди и метаболический синдром связаны через цикл ингибитора активатора плазминогена-1. Биологические исследования . 2007 октября, 29 (10): 1029-38. [Медлайн].
Hsing AW, Sakoda LC, Chua S. Jr. Ожирение, метаболический синдром и рак простаты. Am J Clin Nutr . 2007 сентябрь 86 (3): s843-57.[Медлайн].
Готтлиб А.Б., Данн Ф., Ментер А. Псориаз и метаболический синдром. J Лекарства Дерматол . 2008 июн. 7 (6): 563-72. [Медлайн].
Коэн А.Д., Шерф М., Видавский Л. и др. Связь между псориазом и метаболическим синдромом. Поперечное исследование. Дерматология . 2008. 216 (2): 152-5. [Медлайн].
Harding, A. Метаболический синдром резко повышает риск повторной преэклампсии.Новости Medscape от WebMD. 21 марта 2013 г. Доступно по адресу http://www.medscape.com/viewarticle/781185. Доступ: 26 марта 2013 г.
Stekkinger E, Scholten R, van der Vlugt MJ, van Dijk AP, Janssen MC, Spaanderman ME. Метаболический синдром и риск рецидива преэклампсии: ретроспективное когортное исследование. BJOG . 2013 июл.120 (8): 979-86. [Медлайн].
Тейлор В.Х., Маккуин GM. Когнитивная дисфункция, связанная с метаболическим синдромом. Obes Rev. . 2007 Сентябрь 8 (5): 409-18. [Медлайн].
Яффе К. Метаболический синдром и снижение когнитивных функций. Curr Alzheimer Res . 2007 апр. 4 (2): 123-6. [Медлайн].
Новичок JW. Метаболический синдром и психические заболевания. Am J Manag Care . 2007 13 ноября (7 приложение): S170-7. [Медлайн].
Fiedorowicz JG, Palagummi NM, Forman-Hoffman VL, et al. Повышенная распространенность ожирения, метаболического синдрома и факторов риска сердечно-сосудистых заболеваний при биполярном расстройстве. Психиатрия Энн Клин . 2008 июль-сен. 20 (3): 131-7. [Медлайн].
Эспозито К., Чиодини П., Капуано А., Колао А., Джульяно Д. Риск перелома и минеральная плотность костной ткани при метаболическом синдроме: метаанализ. J Clin Endocrinol Metab . 2013 августа 98 (8): 3306-14. [Медлайн].
Джой Т., Лахири П., Поллекс Р.Л. и др. Генетика метаболического синдрома. Curr Diab Rep . 2008 Апрель 8 (2): 141-8. [Медлайн].
Heima NE, Eekhoff EM, Oosterwerff MM, Lips PT, van Schoor NM, Simsek S.Функция щитовидной железы и метаболический синдром у пожилых людей: популяционное исследование. Eur J Endocrinol . 2013 Январь 168 (1): 59-65. [Медлайн].
Пуиг Дж. Г., Мартинес Массачусетс. Гиперурикемия, подагра и метаболический синдром. Curr Opin Rheumatol . 2008 20 марта (2): 187-91. [Медлайн].
Лам JC, ИП MS. Обновленная информация об обструктивном апноэ во сне и метаболическом синдроме. Curr Opin Pulm Med . 2007 13 ноября (6): 484-9.[Медлайн].
[Рекомендации] Гофф Д.К. младший, Ллойд-Джонс Д.М., Беннетт Г. и др. Рекомендации ACC / AHA по оценке риска сердечно-сосудистых заболеваний, 2013 г .: отчет Целевой группы Американского колледжа кардиологов / Американской кардиологической ассоциации по практическим рекомендациям. Тираж . 2014, 24 июня, 129 (25, приложение 2): S49-73. [Медлайн]. [Полный текст].
Welty FK, Alfaddagh A, Elajami TK. Устранение воспаления при метаболическом синдроме. Перевод статьи .2016 Январь 167 (1): 257-80. [Медлайн].
Таппер Т., Гопалакришнан Г. Профилактика развития диабета у лиц с метаболическим синдромом. Med Clin North Am . 2007 ноябрь 91 (6): 1091-105, viii-ix. [Медлайн].
Джеймс П.А., Опарил С., Картер Б.Л. и др. Основанное на фактах руководство 2014 года по лечению высокого кровяного давления у взрослых: отчет членов комиссии, назначенных в Восьмой объединенный национальный комитет (JNC 8). JAMA .2014 5 февраля. 311 (5): 507-20. [Медлайн].
Eckel RH, Jakicic JM, Ard JD, et al. Руководство AHA / ACC 2013 г. по управлению образом жизни для снижения риска сердечно-сосудистых заболеваний: отчет Целевой группы Американского колледжа кардиологов / Американской кардиологической ассоциации по практическим рекомендациям. Джам Колл Кардиол . 2014 июл 1. 63 (25, пт B): 2960-84. [Медлайн].
Jensen MD, Ryan DH, Apovian CM, et al. Руководство AHA / ACC / TOS, 2013 г., по лечению избыточной массы тела и ожирения у взрослых: доклад Целевой группы Американского колледжа кардиологов / Американской кардиологической ассоциации по практическим рекомендациям и Общества ожирения. Джам Колл Кардиол . 2014 июл 1. 63 (25, часть B): 2985-3023. [Медлайн].
Kini S, Herron DM, Yanagisawa RT. Бариатрическая хирургия при патологическом ожирении — лекарство от метаболического синдрома ?. Med Clin North Am . 2007 ноябрь 91 (6): 1255-71, xi. [Медлайн].
Багри Х.С., Рагхавендран С., Карли Ф. Метаболический синдром и инсулинорезистентность: периоперационные соображения. Анестезиология . 2008 Март 108 (3): 506-23. [Медлайн].
Драгер Л.Ф., Тогейро С.М., Полоцкий В.Ю., Лоренци-Филхо Г. Обструктивное апноэ во сне: кардиометаболический риск при ожирении и метаболическом синдроме. Джам Колл Кардиол . 2013 13 августа. 62 (7): 569-76. [Медлайн].
Шарма С.К., Агравал С., Дамодаран Д. и др. СИПАП при метаболическом синдроме у пациентов с синдромом обструктивного апноэ сна. N Engl J Med . 2011 декабрь 15. 365 (24): 2277-86. [Медлайн].
Suzuki T, Homma S.Лечение артериальной гипертензии и других факторов риска сердечно-сосудистых заболеваний у пациентов с метаболическим синдромом. Med Clin North Am . 2007 ноябрь 91 (6): 1211-23, x. [Медлайн].
Таун С.П., Тара Э. Уменьшают ли статины события у пациентов с метаболическим синдромом ?. Curr Atheroscler Rep . 2008 10 февраля (1): 39-44. [Медлайн].
Nissen SE, Tardif JC, Nicholls SJ, et al. Влияние торцетрапиба на прогрессирование коронарного атеросклероза. N Engl J Med . 2007 29 марта. 356 (13): 1304-16. [Медлайн].
Cannon CP, Дански Х.М., Дэвидсон М. и др. Дизайн исследования DEFINE: определение эффективности и переносимости ингибирования CETP с помощью AnacEtrapib. Am Heart J . 2009 Октябрь 158 (4): 513-519.e3. [Медлайн].
Barter PJ, Rye KA. Есть ли роль фибратов в лечении дислипидемии при метаболическом синдроме? Артериосклер тромб Vasc Biol .2008, 28 января (1): 39-46. [Медлайн].
Ито МК. Метаболический синдром: патофизиология, клиническая значимость и использование ниацина. Энн Фармакотер . 2004 Февраль 38 (2): 277-85. [Медлайн].
Stone NJ, Робинсон JG, Lichtenstein AH, et al. Руководство ACC / AHA 2013 г. по лечению холестерина в крови для снижения риска атеросклеротических сердечно-сосудистых заболеваний у взрослых: отчет Целевой группы Американского колледжа кардиологов / Американской кардиологической ассоциации по практическим рекомендациям. Джам Колл Кардиол . 2014 июл 1. 63 (25, часть B): 2889-934. [Медлайн].
Chan DC, Watts GF, Barrett PH и др. Регулирующие эффекты ингибитора HMG CoA редуктазы и рыбьего жира на кинетику аполипопротеина B-100 у инсулинорезистентных мужчин с ожирением и дислипидемией. Диабет . 2002 августа 51 (8): 2377-86. [Медлайн].
Орчард Т.Дж., Темпроса М., Голдберг Р. и др. Влияние метформина и интенсивного изменения образа жизни на метаболический синдром: рандомизированное исследование Программы профилактики диабета. Энн Интерн Мед. . 2005, 19 апреля. 142 (8): 611-9. [Медлайн].
Nieuwdorp M, Stroes ES, Kastelein JJ. Нормализация метаболического синдрома с помощью фенофибрата, метформина или их комбинации. Диабет, ожирение, метаболизм . 2007 9 ноября (6): 869-78. [Медлайн].
Derosa G, D’Angelo A, Ragonesi PD, et al. Метаболические эффекты пиоглитазона и розиглитазона у пациентов с диабетом и метаболическим синдромом, получавших метформин. Intern Med J . 2007 Февраль 37 (2): 79-86. [Медлайн].
Di Pino A, DeFronzo RA. Инсулинорезистентность и атеросклероз: последствия для инсулино-сенсибилизирующих агентов. Endocr Ред. . 2019 декабрь 1. 40 (6): 1447-67. [Медлайн].
Bragt MC, Popeijus HE. Рецепторы, активируемые пролифератором пероксисом, и метаболический синдром. Physiol Behav . 2008 г. 23 мая. 94 (2): 187-97. [Медлайн].
[Рекомендации] Skyler JS, Bergenstal R, Bonow RO, et al.Интенсивный гликемический контроль и предотвращение сердечно-сосудистых событий: последствия испытаний диабета ACCORD, ADVANCE и VA: заявление о позиции Американской диабетической ассоциации и научное заявление Американского колледжа кардиологического фонда и Американской кардиологической ассоциации. Джам Колл Кардиол . 2009 20 января. 53 (3): 298-304. [Медлайн].
Shields TM, Hennekens CH. Лечение метаболического синдрома: аспирин. Endocrinol Metab Clin North Am .2004 сентябрь 33 (3): 577-93, vii. [Медлайн].
Блаха М.Дж., Бансал С., Руф Р. и др. Практический «ABCDE» подход к метаболическому синдрому. Mayo Clin Proc . 2008 августа 83 (8): 932-41. [Медлайн].
Инь Дж, Чжан Х., Йе Дж. Традиционная китайская медицина в лечении метаболического синдрома. Препараты для лечения иммунных расстройств Endocr Metab нацелены на . 2008 июн. 8 (2): 99-111. [Медлайн].
Холландер JM, Mechanick JI.Дополнительная и альтернативная медицина и лечение метаболического синдрома. J Am Diet Assoc . 2008 Март 108 (3): 495-509. [Медлайн].
Руидавец Дж. Б., Бонгард В., Даллонжвилл Дж. И др. Высокое потребление зерна, рыбы, молочных продуктов и их сочетаний связано с низкой распространенностью метаболического синдрома. J Epidemiol Community Health . 2007 Сентябрь 61 (9): 810-7. [Медлайн].
Mattei J, Bhupathiraju S, Tucker KL.Более высокая приверженность к диете, основанная на рекомендациях Американской кардиологической ассоциации, связана с более низкими шансами аллостатической нагрузки и метаболического синдрома у взрослых пуэрториканцев. J Nutr . 2013 ноябрь 143 (11): 1753-9. [Медлайн]. [Полный текст].
Эспозито К., Чиотола М., Джульяно Д. Средиземноморская диета и метаболический синдром. Mol Nutr Food Res . 2007 Октябрь, 51 (10): 1268-74. [Медлайн].
Буитраго-Лопес А., Сандерсон Дж., Джонсон Л. и др.Потребление шоколада и кардиометаболические расстройства: систематический обзор и метаанализ. BMJ . 2011 26 августа. 343: d4488. [Медлайн]. [Полный текст].
Лю Л., Ван И, Лам К.С. и др. Умеренное употребление вина для профилактики метаболического синдрома и связанных с ним медицинских осложнений. Препараты для лечения иммунных расстройств Endocr Metab нацелены на . 2008 июн.8 (2): 89-98. [Медлайн].
Чо С., Намкунг К., Шин М. и др. Сердечно-сосудистые защитные эффекты и клиническое применение ресвератрола. J Med Food . 27 марта 2017 г. [Medline].
Миллер А., Адели К. Диетическая фруктоза и метаболический синдром. Curr Opin Gastroenterol . 2008 24 марта (2): 204-9. [Медлайн].
Johnson RJ, Segal MS, Sautin Y, et al. Возможная роль сахара (фруктозы) в эпидемии гипертонии, ожирения и метаболического синдрома, диабета, заболеваний почек и сердечно-сосудистых заболеваний. Am J Clin Nutr . 2007 Октябрь 86 (4): 899-906.[Медлайн].
Vrolix R, van Meijl LE, Mensink RP. Метаболический синдром в зависимости от гликемического индекса и гликемической нагрузки. Physiol Behav . 2008 г. 23 мая. 94 (2): 293-9. [Медлайн].
Юберо-Серрано Е.М., Дельгадо-Листа Дж., Тирни А.С. и др. Резистентность к инсулину определяет дифференцированный ответ на изменение диетического жира на факторы риска метаболического синдрома: исследование LIPGENE. Am J Clin Nutr . 2015 декабрь102 (6): 1509-17. [Медлайн].
Робертс К.К., Хевенер А.Л., Барнард Р.Дж. Метаболический синдром и инсулинорезистентность: основные причины и модификация при тренировках. Compr Physiol . 2013 Январь 3 (1): 1-58. [Медлайн].
Fappa E, Yannakoulia M, Pitsavos C, et al. Вмешательство в образ жизни при лечении метаболического синдрома: можем ли мы улучшить соблюдение режима лечения ?. Питание . 2008 24 марта (3): 286-91. [Медлайн].
Департамент здравоохранения и социальных служб, Центры по контролю и профилактике заболеваний.Физическая активность и здоровье: польза от физической активности. Доступно по адресу http://www.cdc.gov/physicalactivity/basics/pa-health/index.htm. Доступ: 26 декабря 2015 г.
Broekhuizen LN, Boekholdt SM, Arsenault BJ, et al. Физическая активность, метаболический синдром и коронарный риск: проспективное популяционное исследование EPIC-Norfolk. Eur J Cardiovasc Назад Rehabil . 2011 Апрель 18 (2): 209-17. [Медлайн].
Шуваль К., Финли К.Э., Шартье К.Г., Баласубраманян Б.А., Габриэль К.П., Барлоу К.Э.Кардиореспираторная подготовка, потребление алкоголя и частота метаболического синдрома у мужчин. Медико-спортивные упражнения . 2012 ноябрь 44 (11): 2125-31. [Медлайн].
Hamilton MT, Hamilton DG, Zderic TW. Роль низкого расхода энергии и сидения при ожирении, метаболическом синдроме, диабете 2 типа и сердечно-сосудистых заболеваниях. Диабет . 2007 ноябрь 56 (11): 2655-67. [Медлайн].
[Рекомендации] Goldstein LB, Bushnell CD, Adams RJ, et al.Рекомендации по первичной профилактике инсульта: руководство для медицинских работников Американской кардиологической ассоциации / Американской ассоциации инсульта. Инсульт . 2011 Февраль 42 (2): 517-84. [Медлайн].
Холл MH, Малдун MF, Дженнингс-младший, Buysse DJ, Флори JD, Manuck SB. Самостоятельно сообщаемая продолжительность сна связана с метаболическим синдромом у взрослых людей среднего возраста. Сон . 2008 май. 31 (5): 635-43. [Медлайн]. [Полный текст].
Кавада Т., Оцука Т.Влияние сменной работы на развитие метаболического синдрома через 3 года у японских рабочих-мужчин. Arch Environ Occup Health . 2014. 69 (1): 55-61. [Медлайн].
McCall B. Ресвератрол способствует формированию костной ткани у мужчин с ожирением. Медицинские новости Medscape от WebMD. Доступно на http://www.medscape.com/viewarticle/833807. 24 октября 2014 г .; Доступ: 25 октября 2014 г.
Орнструп MJ, Harslof T, Kjaer TN, Langdahl BL, Pedersen SB. Ресвератрол увеличивает минеральную плотность костей и щелочную фосфатазу костей у мужчин с ожирением: рандомизированное плацебо-контролируемое исследование. J Clin Endocrinol Metab . 2014 декабрь 99 (12): 4720-9. [Медлайн].
Марсо С.П., Дэниэлс Г.Х., Браун-Франдсен К. и др., Для Руководящего комитета ЛИДЕР, Исследователи судебных процессов ЛИДЕР. Лираглутид и сердечно-сосудистые исходы при диабете 2 типа. N Engl J Med . 2016 28 июля. 375 (4): 311-22. [Медлайн].
Марсо С.П., Бейн С.К., Консоли А. и др., Для исследователей SUSTAIN-6. Семаглутид и сердечно-сосудистые исходы у пациентов с диабетом 2 типа. N Engl J Med . 2016 10 ноября. 375 (19): 1834-44. [Медлайн]. [Полный текст].
Bateman LA, Slentz CA, Willis LH, et al. Сравнение эффектов аэробных тренировок и тренировок с отягощениями на метаболический синдром (из исследований целевого снижения риска посредством определенных упражнений — STRRIDE-AT / RT). Ам Дж. Кардиол . 2011 Сентябрь 15. 108 (6): 838-44. [Медлайн].
Браузер Д. Переедание с большей вероятностью приводит к метаболическому синдрому у мужчин.Медицинские новости Medscape от WebMD. Доступно на http://www.medscape.com/viewarticle/811732. 26 сентября 2013 г .; Доступ: 7 октября 2013 г.
Удо Т., Макки С.А., Уайт М.А., Машеб Р.М., Барнс Р.Д., Грило С.М. Половые различия в биопсихосоциальных коррелятах компульсивного переедания: исследование обращающихся за лечением взрослых с ожирением в учреждениях первичной медико-санитарной помощи.