Способности
Способностями называют индивидуально-психологические особенности, которыми обладает личность. Эти особенности способны обеспечить успех в деятельности, быстроту и легкость овладения деятельностью.
Способности не являются одним и тем же, что и умения, знания или навыки, которые свойственны человеку, при этом способности способствуют их быстрому приобретению, фиксации и эффективному практическому применению.
Природные и специфические виды способностей
Способности могут быть природными и специфическими.
Природные способности или естественные, обусловлены биологически. Они связаны с врожденными задатками, их формирование происходит на базе этих задатков, при этом важным условием служит наличие жизненного опыта через механизмы научения типа условно-рефлекторных связей.
Специфические способности человека имеют общественно-историческое происхождение и обеспечивают жизнь и социальное развитие.
Особенности специфических способностей человека
В свою очередь специфические способности человека подразделяются на общие и специальные.
Общие специфические способности могут быть определены успехами человека в самых различных видах деятельности и общения (уровень интеллекта, уровень развития памяти и речи, умственные способности).
Специальные специфические способности характеризуют успехи в конкретных видах деятельности и общения. В таких видах деятельности необходимы особого рода задатки и их развитие (способности литературно-лингвистические, математические, художественно-творческие, технические, спортивные).
Другие виды способностей
Можно выделить дополнительно другие виды человеческих способностей.
Общие способности подразумевают наличие способностей к распознаванию закономерностей, сюда входит и общий уровень интеллекта.
Модально общие способности включают вербальный интеллект, пространственный интеллект, технико-практический интеллект, нумерический интеллект, он позволяет производить вычисления.
Специальные способности. С их помощью осуществляется более легкое и успешное овладение определенными специальными видами деятельности: артистические, технические, музыкальные, педагогические.
Частные способности отвечают за уровень развития психических познавательных функций. К психическим познавательным функциям относятся память, внимание, восприятие, мышление, воображение.
Теоретические способности определяют склонность человека к абстрактно-логическому мышлению.
Практические способности лежат в основе склонности к конкретно-практическим действиям.
Сочетанием теоретических и практических способностей обладают одаренные и разносторонние люди.
Учебные способности оказывают влияние на успешную реализацию педагогического воздействия, уровень усвоения человеком знаний, умений, навыков. Учебные способности также способствуют формированию качеств личности.
Творческие способности связаны с успешностью создания произведений материальной и духовной культуры, новых идей, открытий, изобретений.
Проявление творческих способностей в высшей степени называется гениальностью. Проявление творческих способностей в конкретной деятельности называется талантом.
Коммуникативные способности и предметно-деятельностные способности обладают связью с взаимодействием людей и природы, техники, знаковой информации, художественными образами.
Нужна помощь преподавателя?
Опиши задание — и наши эксперты тебе помогут!
Описать заданиеСпособности и задатки
Общая одаренность человека выражается в способности к различным видам деятельности и общения. Это значит, что человек имеет единство общих способностей. Оно обуславливает диапазон интеллектуальных возможностей, уровень и своеобразие деятельности и общения личности.
Таким образом можно заключить, что способностями являются индивидуально психологические особенности человека. Они проявляются в деятельности, и считаются критерием успешности выполнения деятельности.
Хотя способности не сводятся к умениям или знаниям, от них зависят многие критерии успешного овладения умениями, знаниями и навыками.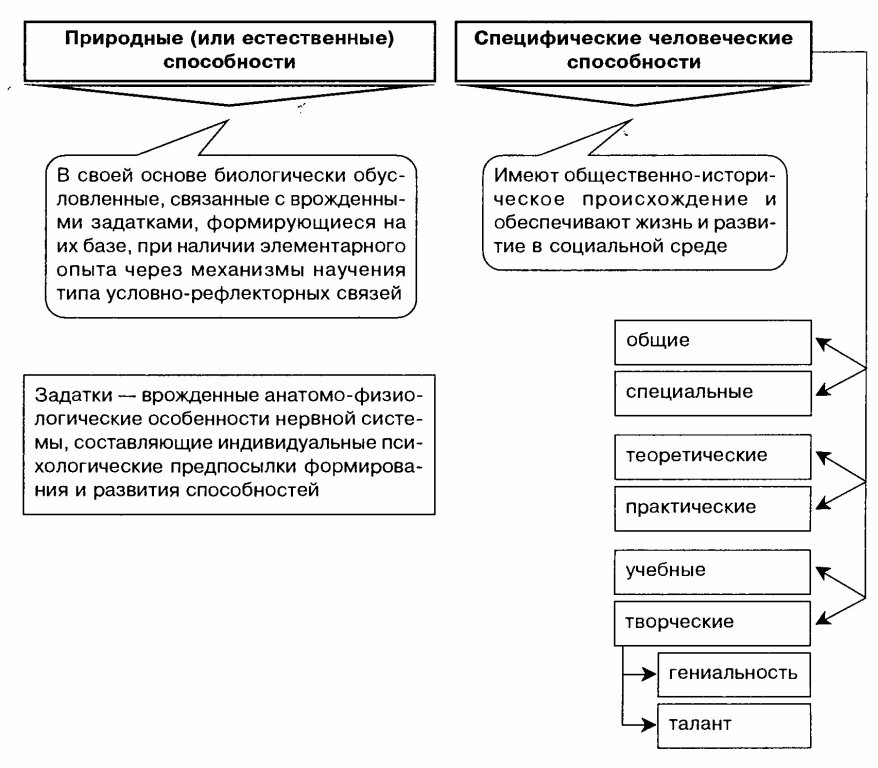
Согласно исследованиям, способности являются пожизненными образованиями. Процесс индивидуальной жизни обуславливает развитие пожизненных образований, а среда и воспитание формируют их.
Советский психолог и основатель школы дифференциальной психологии Б. М. Теплов произвел глубокий анализ проблемы способностей. Он развивал концепцию, в которой говорилось о анатомо-физиологических и функциональных особенностях человека.
Они могут считаться врожденными в том случае, если создают определенные предпосылки для развития способностей. Эти способности называются задатками.
Задатками называют некоторые врожденные генетически анатомо-физиологические особенности нервной системы. Задатки образуют индивидуально-природные предпосылки к формированию и развитию способностей.
Способности отличаются динамичностью образования. В процессе они могут формироваться и развиваться определенным образом организованной деятельности и общения. Способности развиваются постепенно.
Задатки характеризуются многозначностью. Они являются всего лишь предпосылками развития способностей.
Те способности, которые развиваются на основе задатков, могут быть обусловлены ими, но никак не предопределяться.
Задатки сами по себе не направлены. Хотя они способны влиять на процесс формирования и развития способностей, но не решающим образом.
Сформированные способности в свою очередь остаются навсегда, их образование происходит в процессе деятельности и воспитания. Разные пути формирования способностей обуславливаются задатками.
Некоторые задатки человек так и не реализовывает, они не превращаются в реальные способности и достижения человека.
Реализация перехода задатков в способности происходит при усилии человека или же благодаря каким-либо обстоятельствам. Человек может даже и не подозревать о наличии в себе каких-либо задатков.
Каждая способность обладает структурой. В структуре есть ведущие и вспомогательные свойства.
Например, ведущие свойства, которые характерны для людей с литературными способностями, включают особенности творческого воображения и мышления, яркие образы памяти, развитие эстетических чувств, чувство языка.
Для ведущих математических способностей характерно умение обобщать. Люди с математическими способностями имеют гибкие мыслительные процессы, они могут легко переходить от прямого к обратному ходу мысли.
Уровни способностей
Способности делятся на несколько уровней: репродуктивный и творческий.
На репродуктивном уровне реализуется высокое умение усваивать готовое знание, человек на этом этапе овладевает сложившимися образцами деятельности и общения.
На творческом уровне создается что-то новое, оригинальное. При этом важно отметить, что оба уровня обладают элементами друг друга.
Человек может обладать разными способностями, при этом одна из них проявляется наиболее значительно. С другой стороны, разные люди имеют одни и те же способности, но они различаются между собой по уровню развития.
Некоторые зарубежные психологи как Р. Кет-тел, Г. Айзенк, А. Бине, Ч. Спирмен предприняли попытки в начале XX в. измерить способности.
Для этого они использовали тесты.
Успешность выполненной деятельности может быть определена не конкретной и отдельной способностью, а только сочетанием способностей, своеобразным у каждого человека.
Успешное овладение той или иной деятельности может быть достигнуто различными путями. Например, при недостаточном развитии одной из способностей может компенсироваться усилением развития другой способности.
Рис. 1. Границы наших задатков (по Лангмейеру)
Благодаря особой генетической предрасположенности и специфике строения мозга человек может обладать высокой одаренностью и гениальностью.
Советский генетик В. П. Эфроимсон утверждал, что данная генетическая предрасположенность встречается у одного человека из тысячи, при этом она достигает необходимого уровня лишь у одного человека из миллиона, а действительно гением может стать лишь один из десяти миллионов.
Также отмечено, что люди с высокой одаренностью имеют также и высокую вероятность развития различных болезней.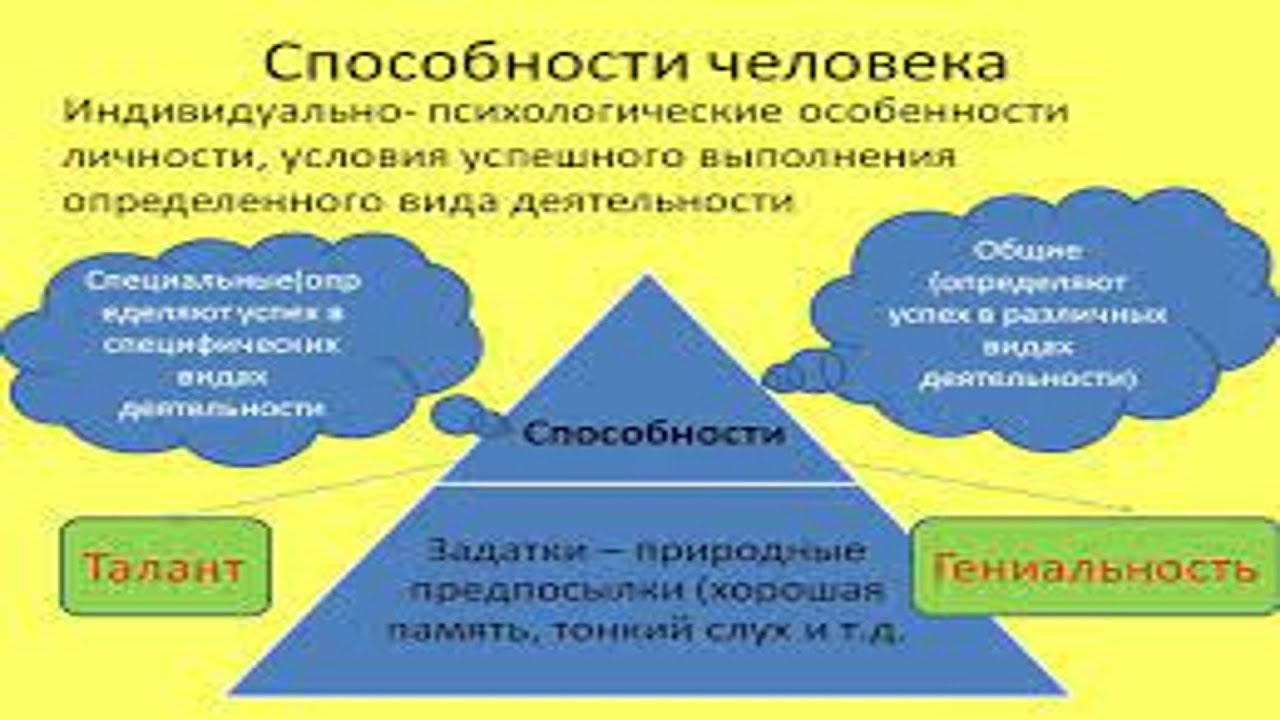
Продолжительность жизни таких людей значительно короче, чем у людей с обычными способностями, одаренные люди чаще подвержены депрессии и имеют более высокий риск душевных заболеваний (в семь раз выше нормы).
Дети «индиго»
80-е года XX в. ознаменовались рождением необычных детей. Необыкновенно одаренные дети получили название дети «индиго». Это название имело связь с цветом их ауры. Цвет индиго это темно-синий цвет с фиолетовым оттенком. Такие дети обладали необычными задатками, способностями к телепатии, ясновидению.
Такие дети отличаются тем, что работа их мозга разделена неравномерно. В нем заметна высокая активность работы правого полушария, в сравнении с большинством взрослых людей, у которых доминирует работа левого полушария мозга. Левое полушарие мозга отвечает за логику, речь, абстрактное мышление.
Из-за такой специфики работы мозга дети «индиго» отличаются высокой интуицией, творческими талантами, способностью получать информацию из всемирного информационного поля.
Ребенок-индиго с детства чувствует свое предназначение, не сомневается в своей значимости, ведет себя очень независимо, проявляет разнообразные таланты.
Способности. Виды способностей — презентация онлайн
1. Способности
Выполнила:Студентка
Группы ДО-4-15
Иванова Анастасия
Преподаватель: Михайлова Ольга Клементьева
2. Понятие
Способности– такие индивидуально психилогические особенности человека,
которые отвечают требованиям данной
деятельности и являются условием
успешного выполнения.
3. Признаки
Способности1. Под способностями понимают
индивидуальные психологические
способности отличающие одного человека от
другого.
2. Способностями называют лишь такие
индивидуальные способности, которые имеют
успешность выполнения деятельностей.
3.
 Под способностями подразумевают
Под способностями подразумеваютиндивидуальные особенности которые не
сводятся к наличным знаниям, умениям и
навыкам, но которые могут объяснить
легкость и быстроту приобретения этих
знаний, умений и навыков.
4. Виды способностей
1. Учебные – связаны сусвоением уже известных
способов выполнения
деятельности приобретением
знаний, умений и навыков.
2. Творческие – связаны с
созданием нового
оригинального продукта с
нахождением новых способов
выполнения деятельности.
3. Общие – проявляются во
многих областях знаний и
деятельности.
4. Специальные – связаны с
определенной
деятельностью, которые
помогают человеку достигать
высоких результатов.
5. Виды специальных способностей
Математические– большую роль играет
математическая память, легкое и свободное
переключение от одной операции к другой.
Конструктивно – технические – точность и
живость пространственных представлений.
Музыкальные – ладовое чувство,
проявляющееся в эмоциональном восприятии
и легком узнавании мелодий.
Литературные – творческая активность и
эстетическая позиция.
Художественно – изобразительные –
способность правильной оценки пропорций,
чувствовать выразительную функцию цвета.
6. Природные предпосылки
Формированиеспособностей происходит на
основе врожденных задатков.
Задатки – анатомо – физиологические
особенности мозга, нервной системы,
составляющие природную основу развития
способностей.
В способностях неразрывно переплетаются
природное (задатки) и приобретенное человеком
в процессе деятельности (социальное). Ведущую
роль играет социальный фактор.
7. Уровни развития способностей
Выделяютдва уровня:
1. Репродуктивный – высокая способность
овладевать умениями, усваивать знания,
овладевать деятельностью и осуществлять
ее по предложенному образцу в
соответствии с предложеной идеей.

2. Творческий – создание нового
оригинального продукта.
8. Уровни развития творческих способностей
Одарённость – своеобразноесочетание способностей,
которое обеспечивает
человеку возможность
успешного выполнения какой –
либо деятельности.
Талант – высокая ступень
развития способностей.
Талантливыми называют
людей способных быстро
усваивать знания и правильно
применять их в жизни и в
своей деятельности.
Гениальность – наивысшая
ступень проявления
творческих сил человека.
Главные способности человека
Люди отличаются не только внешним видом, темпераментом, характером, поведением, но и способностями.
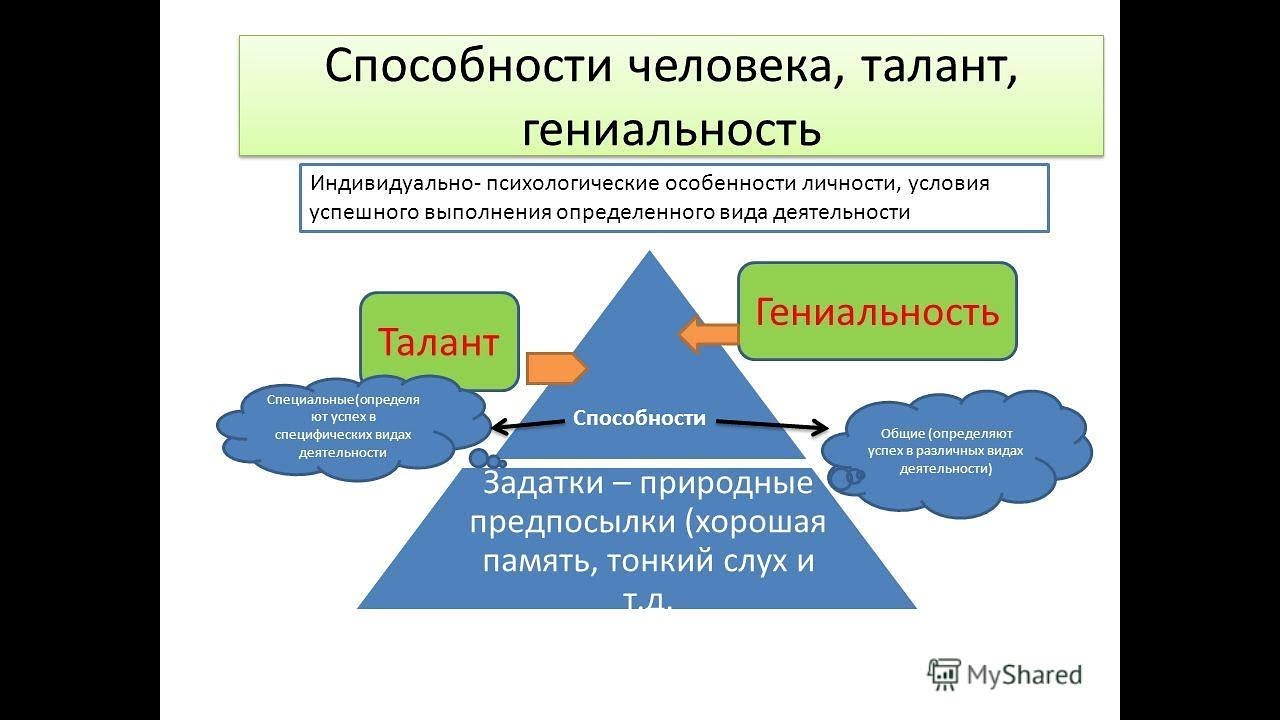
Способности — это стойкие индивидуальные психологические свойства человека, его интеллекта, от которых зависят достижения учебной, трудовой, научной, творческой деятельности, что являются основой жизненного успеха.
Если человек от природы талантлив или даже гениален, но ничему не научился и ничего не создал, его способности останутся нереализованными. Составляющими способностей является природная одарённость и психологические особенности человека, от которых зависит скорость овладения знаниями, умениями и навыками.
Составляющими способностей является природная одарённость и психологические особенности человека, от которых зависит скорость овладения знаниями, умениями и навыками.
Каждый человек имеет способности к определённому виду деятельности. Способности человека проявляются только в результатах учёбы и труда.
Кроме врождённой одарённости, к структуре способностей принадлежат познавательные процессы, знания, умения и навыки, которые человек приобретает в процессе воспитания, учёбы и труда, а также эмоциональные и волевые качества. Зерно станет колосом только после того, как его посеют в землю, будут поливать, удобрять и пропалывать, так и врождённая одарённость станет способностью человека лишь под воздействием воспитания, учёбы и труда. Кроме того, воспитание должно быть не только содержательным и тщательным, но и индивидуальным.
Именно индивидуальный подход в воспитании и учёбе обнаруживает и развивает естественную одарённость и способности человека. Николая Гоголя, например, педагоги оценивали как посредственного ученика, не разглядев его талантливости. Когда о человеке говорят, что он способный, имеют в виду не только его знания, умения и навыки, но и способность пользоваться ими, применять в своей деятельности. Способности имеют не только познавательные, но и эмоциональные свойства. Способные люди всегда переполнены эмоциями, которые ведут их к поиску новых знаний, решений и открытий. Трудно представить себе способного и в то же время безразличного, неэмоционального человека. Существует очень тесная связь между способностями и волевыми качествами человека — инициативой, решительностью, настойчивостью, умением владеть собой, преодолевать трудности.
Когда о человеке говорят, что он способный, имеют в виду не только его знания, умения и навыки, но и способность пользоваться ими, применять в своей деятельности. Способности имеют не только познавательные, но и эмоциональные свойства. Способные люди всегда переполнены эмоциями, которые ведут их к поиску новых знаний, решений и открытий. Трудно представить себе способного и в то же время безразличного, неэмоционального человека. Существует очень тесная связь между способностями и волевыми качествами человека — инициативой, решительностью, настойчивостью, умением владеть собой, преодолевать трудности.
Ответы на вопрос «20.Способности: понятие, виды, структура.»
Понятие способности используется в быту для объяснения случаев, когда разные люди в одинаковых условиях добиваются разных успехов (особенно если эти успехи очень сильно разнятся между собой). В этой связи можно сразу указать на тот феномен, что люди вообще-то очень часто склонны выдавать своё «не хочу» за «не могу». Под этим «не хочу» может скрываться безволие, лень, низкая мотивация иные личностные особенности. И за этим «не могу» (низкими способностями) во многих случаях скрывается психологическая защита. Размытость бытового понимания феномена способностей оказало влияние и на теоретическую психологию.
Под этим «не хочу» может скрываться безволие, лень, низкая мотивация иные личностные особенности. И за этим «не могу» (низкими способностями) во многих случаях скрывается психологическая защита. Размытость бытового понимания феномена способностей оказало влияние и на теоретическую психологию.
Слово «способность» имеет очень широкое применение в самых различных областях практики. Обычно способности тесно привязаны к тому или иному виду выполняемой деятельности: высокие способности — качественная и эффективная деятельность, низкие способности — некачественная и неэффективная деятельность.
Виды способностей различают по их направленности, или специализации:
— общие способности — индивидуальные свойства личности, обеспечивающие относительную легкость и продуктивность в овладении знаниями и осуществлении различных видов деятельности;
— специальные способности — свойства личности, помогающие достичь высоких результатов в какой-либо области деятельности.
Как считал Б. М. Теплов, способности могут существовать только в постоянном процессе развития. Способности, которые не развиваются, со временем утрачиваются. Примеры областей человеческой деятельности, в которых развиваются способности:
— наука,
— музыка,
— техническое творчество,
— художественное творчество,
— литература,
— математика,
— спорт.
Тезис о необходимости развития способностей может иметь и биологический подтекст. Как показывают исследования, гены людей и животных могут находиться в активированном или не активированном состоянии. Условия среды обитания, образ жизни влияет на то, будут ли активизированы гены или не будут. Это ещё один механизм адаптации, придуманный природой для живых существ.
Классификация способностей
Успешность выполнения деятельности обычно зависит не от какой-либо одной, но от сочетания различных способностей. Что характерно, разные сочетания способностей могут дать близкий результат. В случае отсутствия необходимых задатков их дефицит может быть восполнен за счёт более высокого развития других задатков и способностей.
В случае отсутствия необходимых задатков их дефицит может быть восполнен за счёт более высокого развития других задатков и способностей.
Б. М. Теплов утверждал, что «Одной из важнейших особенностей психики человека является возможность чрезвычайно широкой компенсации одних свойств другими, вследствие чего относительная слабость какой-нибудь одной способности вовсе не исключает возможности успешного выполнения даже такой деятельности, которая наиболее тесно связана с этой способностью. Недостающая способность может быть в очень широких пределах компенсирована другими, высокоразвитыми у данного человека».
Близость способностей друг к другу, способность их взаимозаменяться, позволяет классифицировать способности. Однако неоднородность проблемы способностей привела к тому, что классификации значительно отличаются друг от друга.
Первое основание классификации
Одно из оснований классификации — степень естественности способностей:
— природные (естественные) способности (то есть биологически обусловленные),
— специфические человеческие способности (имеющие общественно-историческое происхождение.
Естественными элементарными способностями являются:
— восприятие,
— память,
— основы коммуникации.
Задатки человека и задатки животного — не одно и то же. У человека на базе задатков формируются способности. Формирование способности происходит при наличии элементарного жизненного опыта, через механизмы научения и т.д.
Специфически человеческие способности:
— общие,
— специальные способности,
— высшие интеллектуальные способности.
Общие способности свойственны большинству людей и определяют успех человека в различных видах деятельности:
— мыслительные способности,
— тонкость и точность ручных движений,
— память,
— речь и др.
Специальные способности определяют успех человека в специфических видах деятельности, для осуществления которых необходимы задатки особого рода и их развитие:
— музыкальные способности,
— математические способности,
— лингвистические способности,
— технические способности,
— литературные способности,
— художественно-творческие способности,
— спортивные способности и др.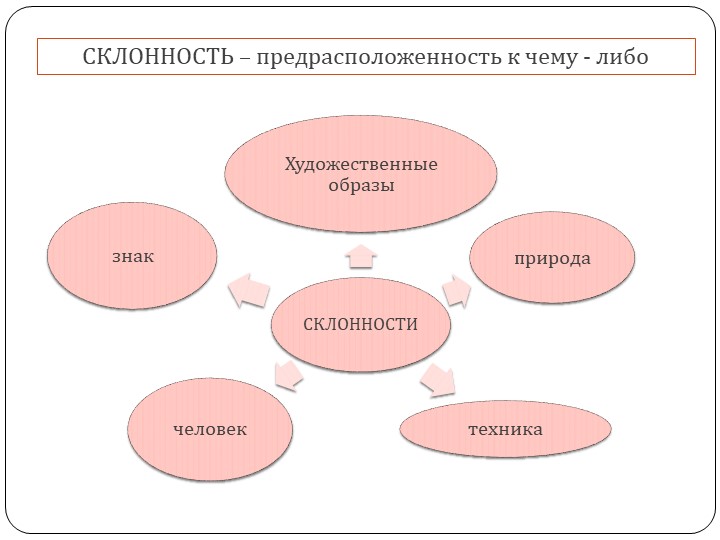
Интеллектуальные способности могут подразделяться на:
— теоретические способности,
— практические способности,
— учебные способности,
— творческие способности,
— предметные способности,
— межличностные способности.
Данные виды способностей тесно связаны друг с другом, переплетаются. Наличие у человека, скажем, общих способностей не исключает развития специальных способностей, как и наоборот. Общие, специальные и высшие интеллектуальные способности не конфликтуют, а сосуществуют, друг друга дополняют и обогащают. В отдельных случаях высокий уровень развития общих способностей может выступать в качестве специальных способностей по отношению к определённым видам деятельности.
Практическая ориентированность
Другим основанием классификации способностей является степень их практической ориентированности:
— теоретические способности,
— практические способности.
Теоретические способности обеспечивают качество и эффективность абстрактно-теоретических размышлений, практические — конкретные предметные действия. Развитие того или иного вида способностей здесь тесно связано со склонностями человека: что он любит, теоретизировать или действовать. Поэтому часто можно наблюдать, что у одних людей хорошо развиты только теоретические способности (разные), у других — только практические.
Развитие того или иного вида способностей здесь тесно связано со склонностями человека: что он любит, теоретизировать или действовать. Поэтому часто можно наблюдать, что у одних людей хорошо развиты только теоретические способности (разные), у других — только практические.
Учебные — творческие
Другое основание деления способностей — по характеру работы с информацией:
— учебные способности,
— творческие способности.
Учебные способности — это академические способности, то есть то, насколько человек качественно и эффективно усваивает учебный материал. Творческие способности — способности что-то сотворить, свершить, придумать, реализовать. Творческие способности это не только способности в области «изящных искусств», но изобретательство, рационализаторство и т.п.
Глава 5 Способности. Общая психология
Глава 5 Способности
5.1. Способности и их виды
Способности – психологические особенности человека, являющиеся условиями успешного выполнения деятельности, приобретения знаний, умений и навыков.
Выделяют естественные и специфические способности человека. Естественные (природные) способности связаны с врожденными задатками человека. В данном случае можно говорить о способностях как о рефлексах, возможности реагирования на внешние раздражители. Специфические способности проявляются в социальной среде и являются индивидуальными для каждого человека. Можно выделить несколько видов специфических способностей.
– Общие способности. Определяют успешность человека в различных областях и служат основой для развития специальных способностей.
Общими способностями можно считать интеллект, развитые познавательные процессы. Зачастую общие и специальные способности делятся на элементарные и сложные.
Общие элементарные способности присущи всем людям. К ним относят основные психические процессы человека – ощущение, восприятие, память, воображение.
Общие сложные способности являются способностями к различным видам деятельности – труду, игре, общению.
– Специальные способности. Определяют успешность в определенных видах деятельности. Выделяют математические, музыкальные, литературные и другие способности.
Специальные элементарные способности являются заметной выраженностью общих способностей, психических процессов. Примерами специальных элементарных способностей можно считать музыкальный слух и глазомер.
Музыкальный слух – качество слухового восприятия, проявляющееся в способности к различению музыкальных звуков и точному воспроизведению их.
Глазомер – качество зрительного восприятия, способность фиксировать, оценивать и сравнивать величины зрительно воспринимаемых объектов, расстояний между ними и до них.
– Теоретические способности. Предопределяют успешность в мыслительной, аналитической деятельности.
– Практические способности. Влияют на успешность в практических действиях.
– Учебные способности. Влияют на успешность усвоения человеком знаний, умений и навыков, предопределяют скорость обучения.
– Творческие способности. Связаны с успешностью создания новых идей, предметов или открытий.
Специальные сложные способности присущи не всем людям. Они являются способностями к определенным видам профессиональной деятельности, возникшим в процессе развития человеческой культуры. Эти способности обычно называют профессиональными.
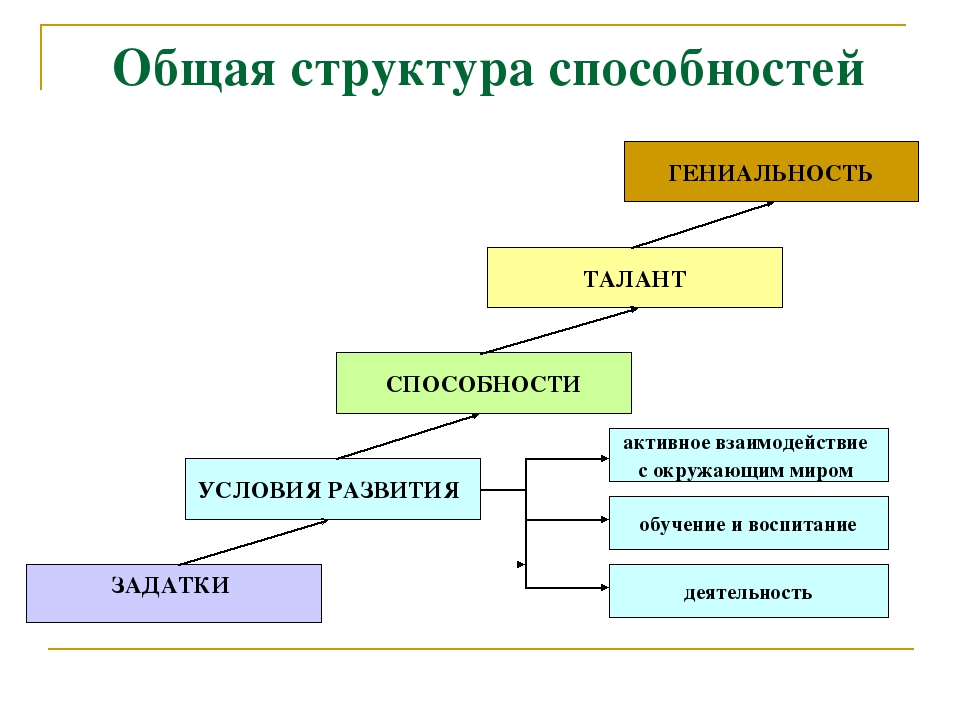
В основе способностей лежат задатки. Задатки – некоторые врожденные особенности телосложения и нервной системы, создающие основу для формирования и развития способностей. На основе одних и тех же задатков могут формироваться различные способности: например, можно назвать массу видов деятельности, в которых может проявить себя человек с развитым музыкальным слухом – музыкант, учитель в музыкальной школе, звукорежиссер, дирижер и т.д.
Задатки предоставляют возможность развития способностей в процессе обучения, воспитания или трудовой деятельности. Определив у ребенка те или иные задатки, можно целенаправленно формировать какую-либо способность.
Задатки зачастую проявляются в склонностях, тяготению к определенному виду деятельности. Выделяют истинные склонности и ложные. Первые характеризуются не просто интересом к какой-либо деятельности, а реальными достижениями в ней. Ложные склонности проявляются в длительном интересе без достижения сколь-нибудь значимых результатов. Зачастую такого рода склонности являются результатом внушения со стороны взрослых.
5.2. Развитие способностей
На развитие способностей могут влиять следующие факторы.
– Наличие задатков, что приводит к проявлению какой-либо способности в большей степени, чем при их отсутствии.
– Компенсация при отсутствии должных задатков – например, люди с расстройствами зрения нередко обладают куда более развитым слухом, чем люди с абсолютным зрением.
– Необходимость решения сложных задач. Люди, живущие в городах, естественно, не в состоянии выполнять целый ряд заданий по сравнению с людьми, живущими в деревнях и селах.
– Особенности передачи знаний, навыков и умений. Так, при равных условиях у двух в целом одинаковых детей способности могут развиться лучше при наличии лучшего учителя. В истории философии и психологии существует множество примеров того, как вокруг одного учителя, видного представителя науки, возникало целое сообщество талантливых учеников. Поэтому вполне естественно, что при должном подходе и в благоприятных условиях даже у «средних» людей могут проявляться различные способности, тогда как при неблагоприятных условиях или в руках плохих педагогов может не найти подходящей почвы для развития даже способный ученик.
Так, при равных условиях у двух в целом одинаковых детей способности могут развиться лучше при наличии лучшего учителя. В истории философии и психологии существует множество примеров того, как вокруг одного учителя, видного представителя науки, возникало целое сообщество талантливых учеников. Поэтому вполне естественно, что при должном подходе и в благоприятных условиях даже у «средних» людей могут проявляться различные способности, тогда как при неблагоприятных условиях или в руках плохих педагогов может не найти подходящей почвы для развития даже способный ученик.
– Условия внешней среды. Не секрет, что способности развиваются в определенной среде, которая оказывает зачастую определяющее значение на их проявление. На процесс развития способностей могут влиять сотни факторов внешней среды К примеру, достижений в спорте труднее добиться жителям мегаполисов, не имеющим условий для тренировок. В то же время высокий уровень развития технологий, как, например, в Японии, оказывает непосредственное влияние на степень развитости интеллекта людей – средний показатель интеллекта превосходит в Японии показатели стран третьего мира.
– Инициатива также влияет на развитие способностей. При определенной свободе, не боясь за результат, человек лучше проявит свои способности, чем если он будет скован в их проявлении.
В процессе развития способностей можно выделить несколько этапов.
– В период с рождения до 5 лет происходит совершенствование работы анализаторов, развитие отдельных участков коры головного мозга, работы органов движения. На данном этапе происходит развитие общих способностей, что создает предпосылку для развития специальных способностей на следующих этапах.
– Младший и средний школьный возраст – время ускоренного развития специальных способностей. Это происходит за счет игровой деятельности, в которой ребенок развивает свое творчество, волевые процессы, двигательные и художественные способности. Помимо игровой деятельности в школьном возрасте влиять на развитие способностей начинают учебная и трудовая деятельность. Большое значение для раскрытия способностей ребенка играет такая деятельность, которая находится в «зоне ближайшего развития». Различные способности ребенка должны развиваться в комплексе, не быть оторванными друг от друга. Отдавая ребенка в секции или кружки, родители делают акцент на одних способностях в ущерб другим. Это может привести к тому, что и в выбранной области ребенок не добьется должного процесса, и остальные способности у него будут развиты слабее, чем у сверстников.
Различные способности ребенка должны развиваться в комплексе, не быть оторванными друг от друга. Отдавая ребенка в секции или кружки, родители делают акцент на одних способностях в ущерб другим. Это может привести к тому, что и в выбранной области ребенок не добьется должного процесса, и остальные способности у него будут развиты слабее, чем у сверстников.Более высокая ступень развития способностей называется талантом. Талант – присущие от рождения определенные способности и умения, которые раскрываются с приобретением навыка и опыта.
Талант может проявляться в любой человеческой деятельности, а не только в области науки или искусства.
Талантливыми также называют людей, способных быстро усваивать знания и правильно применять их в жизни и в своей деятельности. Это талантливые ученики и талантливые студенты, талантливые скрипачи и пианисты, талантливые инженеры и строители.
Талант человека, направляясь выраженной потребностью в творчестве, всегда отражает определенные общечеловеческие запросы. Развитие талантов решающим образом зависит от общественно-исторических условий.
Развитие талантов решающим образом зависит от общественно-исторических условий.
Гениальность – это наивысшая степень проявления творческих сил человека. Это создание качественно иных творений, открывающих новую эру в развитии культуры, науки, практики. Гениальность – это высшая степень развития способностей, делающая человека незаурядной и выдающейся личностью во многих областях.
Литература1. Венгер Л.А. Педагогика способностей. – М.: Педагогика, 1973.
2. Гиппенрейтер Ю.Б. Введение в общую психологию: Курс лекций. – М.: Изд-во МГУ, 1988.
3. Гуревич К.М. Индивидуально-психологические особенности школьников. – М.: Знание, 1988.
4. Кузьмина Н.В. Способности, одаренность, талант учителя. – Л.: Изд-во ЛГУ, 1985.
5. Левитов Н.Д. Психология характера. – М.: Просвещение, 1969.
6. Лейтес Н.С. Способности и одаренность в детские годы. – М.: Знание, 1984.
7. Лейтес Н.С. Умственные способности и возраст. – М.: Педагогика, 1971.
– М.: Педагогика, 1971.
8. Мелхорн Г., Мелхорн Х.-Г. Гениями не рождаются. Общество и способности человека. – М.: Просвещение, 1989.
9. Мерлин B.C. Структура личности. Характер, способности, самосознание: Учеб. пособие к спецкурсу. – Пермь, 1990.
10. Пономарев Я.А. Психология творчества. – М.: Наука, 1976.
11. Рубинштейн С.Л. Основы общей психологии: В 2 т. – М.: Педагогика, 1989.
12. Теплов Б.М. Избранные труды: В 2 т. – М.: Наука, 1985.
13. Чудновский В.Э. Воспитание способностей и формирование личности. – М.: Знание, 1986.
14. Якиманская И.С. Развивающее обучение. – М.: Педагогика, 1979.Данный текст является ознакомительным фрагментом.
Продолжение на ЛитРесХарактеристика способностей.
 Общие и специальные способности
Общие и специальные способностиОтечественная теория способностей создавалась трудами многих выдающихся психологов – Выготским, Леонтьевым, Рубинштейном, Тепловым, Ананьевым, Крутецким, Голубевой.
Теплов, определяя содержание понятия способность, сформулировал 3 ее признака, которые лежат в основе многих работ:
- под способностями подразумеваются индивидуально-психологические особенности, отличающие одного человека от другого;
- они имеют отношение к успешности выполнения какой-либо Деятельности или многим Деятельностям;
- способности не сводятся к наличным навыкам, умениям и знаниям, но могут объяснить легкость и быстроту приобретения этих знаний.
Способность – это психологическая особенность человека и не является врожденным качеством, а представляет собой продукт развития и формирования в процессе какой-либо Деятельности. Но в их основе лежат врожденные анатомо-физиологические особенности – задатки. Хотя способности развиваются на основе задатков, они все же не являются их функцией, задатки – это предпосылки развития способностей. Задатки рассматриваются как неспецифические особенности нервной системы и организма в целом, следовательно отрицается существование для каждой способности своего предуготовленного задатка. На базе разных задатков развиваются разные способности, одинаково проявляющиеся в результатах Деятельности.
Но в их основе лежат врожденные анатомо-физиологические особенности – задатки. Хотя способности развиваются на основе задатков, они все же не являются их функцией, задатки – это предпосылки развития способностей. Задатки рассматриваются как неспецифические особенности нервной системы и организма в целом, следовательно отрицается существование для каждой способности своего предуготовленного задатка. На базе разных задатков развиваются разные способности, одинаково проявляющиеся в результатах Деятельности.
На основе одинаковых задатков у разных людей могут сформироваться разные способности. Отечественные психологи говорят о неразрывной связи способностей с Деятельностью. Способности всегда развиваются в Деятельности и представляют собой активный процесс со стороны человека. Виды Деятельности, в которых формируются способности всегда конкретны и историчны.
Одним из основных принципов отечественной психологии является личностный подход к пониманию способностей. Главный тезис: нельзя сужать содержание понятия «способность» до характеристик отдельных психических процессов.
Главный тезис: нельзя сужать содержание понятия «способность» до характеристик отдельных психических процессов.
I. Проблема способностей возникает при рассмотрении Личности как субъекта Деятельности. Большой вклад в понимание единства способностей и качеств Личности сделан Ананьевым, который рассматривал способность как интеграцию свойств субъективного уровня (свойств человека как субъекта Деятельности). В его теории структура свойств человека имеет 3 уровня:
- Индивидуальный (природный). Это половые, конституциональные и нейродинамические особенности, их высшими проявлениями являются задатки.
- Субъективные свойства характеризуют человека как субъекта труда, общения и познаний и включают особенности внимания, памяти, восприятия и прочее. Интеграцией этих свойств выступают способности.
- Личностные свойства характеризуют человека как социальное существо и связаны прежде всего с социальными ролями, социальным статусом и структурой ценностей.
 Высший уровень в иерархии личностных свойств представлен характером и склонностями человека.
Высший уровень в иерархии личностных свойств представлен характером и склонностями человека.
 Высший уровень в иерархии личностных свойств представлен характером и склонностями человека.
Высший уровень в иерархии личностных свойств представлен характером и склонностями человека.Уникальное сочетание всех свойств человека образует индивидуальность, в которой центральную роль играют личностные свойства, преобразующие и организующие индивидуальные и субъектные свойства.
II. Чаще всего рассматривается взаимоотношение направленности Личности и ее способностей. Интересы, склонности, потребности человека побуждают его к активной Деятельности, в которой формируются и развиваются способности. Успешное выполнение Деятельности, связанное с развитыми способностями благоприятно отражается на формировании положительной мотивации к Деятельности.
III. Велико влияние характерологических черт Личности на формирование способностей. Целеустремление, трудолюбие, упорство необходимы для того, чтобы добиться успехов в решении поставленных задач, а следовательно и развитии способностей. Недостаток волевых черт характера может помешать развитию и проявлению намеченных способностей. Исследователи отмечают характерологические черты, свойственные одаренным людям – инициативность, креативность, высокая самооценка.
Исследователи отмечают характерологические черты, свойственные одаренным людям – инициативность, креативность, высокая самооценка.
Сходные представления о способностях высказывают и зарубежные психологи. Они связывают их с достижениями в различных видах Деятельности, рассматривают как основу достижений, но не связывают способности и достижения как тождественные характеристики.
Способности – это понятие, которое служит для описания, упорядочивания возможностей, определяющих достижения человека. Способностям предшествуют навыки, являющиеся их условием для приобретения в процессе обучения, частых упражнений и тренировок. Достижения в Деятельности зависят не только от способностей, но и от мотивации, психического состояния.
Общие способности – интеллектуальные и творческие способности, которые находят свое проявление во многих разных видах Деятельности.
Специальные способности – определяются по отношению к отдельным специальным областям Деятельности.
Чаще всего соотношение общих и специальных способностей анализируется как соотношение общего и особенного в условиях и результатах Деятельности.
Теплов связывал общие способности с общими моментами в разных видах Деятельности, а специальные – с особыми специфическими моментами.
Потенциал человека и его скрытые способности
«Если вы можете это вообразить, вы можете это и сделать» .
Уолт Дисней
Определение внутреннего потенциала человека – это не самая простая задача. Первое, что необходимо сделать, так это понять, что потенциал – это способность человека реализовать собственные скрытые возможности. У каждого человека внутренний потенциал разный, как и каждый человек индивидуален. Потенциал представляет собой здоровую основу для самореализации в мире и обществе, и является внутренним резервом. В мире, где главенствует левое полушарие, успеха добиваются те, кто умеет использовать как левое(логическое) , так и правое (творческое) полушарие мозга.
Потенциал личности складывается в целостную картину из множества других потенциалов. Именно их уровень развития определяет, насколько человек будет здоров психологически и успешен, счастлив. В целом, можно выделить следующие виды потенциалов человека:
- Потенциал разума. Его можно охарактеризовать, как способность человека совершенствовать собственное мышление. Когда приходится решать различного рода жизненные задачи, ум становится подвижным, задействуется реалистичное, творческое, системное, дивергентное мышление. Однако человек с неразвитым потенциалом ума при возникновении проблем часто впадает в депрессию и у него могут возникать ощущения усталости от жизни.
- Потенциал воли. Он отражает то, насколько человек способен к самореализации, ставить перед собой цели и какие адекватные пути их достижения он выберет. Как утверждают ученые, в случаях, когда потенциал воли развит слабо, у человека возникают проблемы с психикой (чаще всего неврозы). Однако когда удается взрастить в себе волю, то людям становится проще жить, им удается определить свое место в жизни, создать личную систему ценностей и выстраивать стратегию, которая помогает добиваться поставленных целей.
- Потенциал чувств. Он характеризует то, насколько эмоциональная сфера человека богата и насколько адекватно он умеет выражать чувства, а также понимать эмоции других людей. От реализации этого потенциала зависит, насколько человек ощущает себя счастливым. В случае если потенциал чувств не был реализован, возникает ряд психосоматических расстройств. Профилактикой от различных психических болезней и психологических расстройств является умение позитивно смотреть на жизнь.
- Потенциал тела. Каждый человек должен понимать, что его тело – это часть его личности. Поэтому умение совершенствовать его и прислушиваться к нему является крайне важной частью в развитии потенциала в целом.
- Общественный потенциал. От уровня его развития зависит то, насколько человеку легко дается налаживание контактов с окружающими его людьми, развитие коммуникативной культуры и адаптация в социальных условиях.
- Креативный (творческий) потенциал. Он определяет, насколько человек способен к активности, самовыражению, преобразованию мира. Созидательная деятельность и реализация этого потенциала – основа психологической зрелости.
- Духовный потенциал. Как правило, он определяет стиль жизнедеятельности человека и то, как он развивает в себе духовную природу.

 Созидательная деятельность и реализация этого потенциала – основа психологической зрелости.
Созидательная деятельность и реализация этого потенциала – основа психологической зрелости.Все, перечисленное выше, является основой психологической устойчивости человека. Если какой-либо из потенциалов развит слишком слабо, возможен дисбаланс, который приводит к внутренним конфликтам.
Оценка внутренних резервов
Предполагается , что навыки родителей передаются ребенку в момент зачатия. Таким образом, можно утверждать, что многое в человеке зависит от его генетической предрасположенности. Поэтому, определяя потенциал кого-либо, следует изучить качества и способности его родителей. Именно таким образом и формируется природный потенциал – та основа, которая лежит в психологической составляющей здоровья человека.
Второй аспект оценки внутренних резервов человека заключается в определении его способностей к концентрации. Стоит отметить, что эта способность играет большую роль, так как именно она позволяет человеку решать сложные жизненные задачи. Умение сконцентрироваться, позволяет развивать и такие качества, как устойчивость в стрессовых ситуациях и способность выживать в сложных условиях.
При оценке потенциала личности, необходимо учитывать и то, насколько человек коммуникабелен, а также как легко ему удается располагать людей к себе и влиять на них. При этом следует понимать, что оценивать нужно только то общение, которое имеет позитивный характер (то есть нормальное, непринужденное и бесконфликтное). Обращают внимание и на то, умеет ли находить человек компромиссы и «сглаживать углы». Этот момент важен при оценке потенциала тем, что он дает возможность понять, насколько гармонична жизнь человека и определить, насколько легко ему будет добиться поставленных целей. Помимо перечисленного выше, при оценке потенциала необходимо учитывать и удачливость человека, его отношение к неудачам, привычки, увлечении и хобби.
Развитие потенциала
Следует учитывать, что те внутренние резервы, которые были заложены природой, можно не только укреплять, но и развивать. Иногда этот процесс происходит сам собой, под влиянием сложных жизненных обстоятельств. Раскрыть потенциал – это задача, которая состоит из двух составляющих – понять, что было упущено, и выстроить план дальнейших действий. К примеру, девушка в детстве имела способность к рисованию, но жизненные обстоятельства заставили ее забыть об этом увлечении. Она поступила в ВУЗ, получила образование по финансовой специальности и устроилась бухгалтером. Со временем ей стало тяжело, и она осознала, что та работа, которой она занимается, не приносит ей удовольствия. Она собрала последние силы, и начала по вечерам рисовать картины. Удовольствие от этого процесса начали перекрывать все негативное, что происходит вокруг. Это произошло под влиянием того, что девушка дала волю потенциалу, находящемуся «под замком» отсутствия времени и других дел. Развивайте в себе способности смотреть на мир по новому, возможно, не каждый угол обзора будет полезен, но этот навык в итоге окрасит вашу жизнь новыми красками. Мы уже много знаем про работу сознания, и точно можно сказать: сначала появляется идея в творческом полушарии, а потом логическое левое воплащает ее в жизнь.
Когда нужно заняться раскрытием внутренних резервов
Существует несколько признаков того, что человеку крайне необходимо заняться раскрытием собственного потенциала. К ним относятся следующие:
- нехватка энергии на то, чтобы что-то предпринять для изменения своей жизни в лучшую сторону;
- неполадки дома, на работе или в отношениях с близкими;
- возникновение мыслей, что жизнь проходит мимо.
Это все признаки того, что тот внутренний резерв, который был в человеке, исчерпал себя. В такие моменты следует найти источники и начать развивать свой потенциал, для того чтобы исправить ситуацию. Следует начать читать различную духовную литературу (это могут быть книги по психологии или эзотерике, учения философов.) Нужно находить время на медитацию. Необходимо начать вести личный дневник , следует вспомнить свои прошлые успехи.
Внешние источники
Источники извне – это обстоятельства, люди и ситуации, которые заставят понять, чего не хватает для раскрытия собственного потенциала. Это может быть общение с психологом, тренером или духовным наставников, посещение различных тренингов. Стоит отметить, что хорошо помогает общение с детьми и животными. Помимо этого, личностный потенциал хорошо развивается через помощь нуждающимся.
Если человеку удалось предпринять хотя бы первые шаги для развития личного потенциала, ему становится намного проще и легче жить дальше. Всегда стоит помнить, что никогда не поздно приступить к поискам внутренних резервов. Этот момент очень важен для того, чтобы постоянно улучшать качество своей жизни.
УЗ «4-я городская поликлиника» валеолог Наталья Дударчик
Вихревой тур по когнитивной биологии — Ассоциация психологических наук — APS
В течение очень долгого времени существовало два основных лагеря по поведению животных и познанию животных: эксклюзивисты, которые сосредотачиваются на различиях между животными и людьми, и инклюзивисты, которые концентрируют внимание на сходстве между людьми и остальными животными. Королевство. Эта давняя дискуссия началась тысячелетия, когда философы, такие как Аристотель и Декарт, утверждали, что люди являются единственными животными, способными к познанию более высокого порядка, таким как рациональное мышление и язык, и не менее выдающиеся мыслители, такие как Вольтер, Чарльз Дарвин и Дэвид Хьюм, утверждали что само собой разумеется, «что звери наделены мыслью и разумом так же, как человек.”
Перемещая мост между эволюционной биологией и когнитивной наукой, когнитивный биолог из Венского университета В. Текумсе Фитч продемонстрировал, что изучение наших более далеких родственников животных имеет жизненно важное значение для понимания человеческого познания.
«Основная идея, которую я хочу донести до вас сегодня, заключается в том, что в определенном смысле обе эти стороны правы», — подчеркнуло Fitch в своем программном выступлении на Международной конвенции психологических наук в Вене в 2017 году. «И с современной биологической точки зрения нам действительно нужно перевернуть эти идеи с ног на голову и признать очень простой биологический факт: это трюизм, но люди тоже животные.”
Биология общих основ
В основе биологии человека лежит огромное количество общих основ: все живые существа, от бактерий до нарциссов, имеют общий генетический код, а структура нашей нервной системы разделяется с животными низшего порядка, такими как мухи и черви, а также с более близкими родственниками. например бонобо. Но, конечно, каждый вид уникален.
В области когнитивной биологии Fitch исследователи пытаются установить связи между фундаментальной эволюционной биологией (например,г., Дарвин) и когнитивные науки (например, Ноам Хомский и Б. Ф. Скиннер). Но когнитивная биология — это не то же самое, что эволюционная психология, поясняет Fitch. В то время как эволюционная психология фокусируется на человеческом разуме в течение относительно короткого периода эволюции последних 6 миллионов лет, когнитивная биология использует более широкий подход, который восходит к гораздо более ранней эволюции человека.
Наряду с этим в высшей степени сравнительным подходом когнитивные биологи разбивают сложные черты, такие как язык или музыка, на несколько основных компонентов, некоторые из которых могут быть общими для людей и других животных, а некоторые могут быть уникальными для определенного вида.По словам Fitch, когнитивные биологи называют это «разделяй и властвуй» или многокомпонентным подходом. Основываясь на наличии или отсутствии этих компонентов, мы можем составить карту филогенетического дерева, которое позволяет исследователям восстановить эволюционное прошлое определенных когнитивных способностей.
Гомологи и аналоги
Люди имеют много общих черт с нашими ближайшими родственниками, человекообразными обезьянами. У нас большой мозг, большой размер тела, долгая жизнь и продолжительное детство, потому что наш общий предок, который не был шимпанзе, гориллой или человеком, также обладал этими характеристиками.Этот эволюционный процесс называется гомологией: разные виды обладают набором общих черт, потому что они унаследованы от общего предка. По словам Fitch, прелесть гомологии в том, что мы можем использовать ее для восстановления прошлого, глядя на живые виды.
Напротив, идет процесс конвергентной эволюции, в котором разные виды независимо адаптируют сходные черты. Например, люди и птицы двуногие, но не потому, что у нас был общий двуногий предок. Люди и птицы приспособились ходить на двух ногах по разным причинам в разные моменты времени.
Fitch также отметило, что эволюция часто является скорее окольным, чем линейным, с адаптациями, возникающими и исчезающими несколько раз у одного данного вида. Например, большинство людей и некоторые другие приматы являются трихроматами — мы обладаем цветовым зрением благодаря трем различным типам колбочек в наших глазах. С другой стороны, большинство других млекопитающих — дихроматы, лишенные цветового зрения. Если бы мы исследовали только млекопитающих, оказалось бы, что трихроматия — это высокоразвитая адаптация, которую люди разделяют лишь с несколькими другими высокоразвитыми видами.Но расширение сравнительной сети за пределы млекопитающих показывает, что птицы не только обладают трихроматией, но и на самом деле обладают четырьмя разными колбочками — тетрахроматией, объяснил он.
Рыбы, рептилии и земноводные также обладают тетрахроматией, что позволяет предположить, что общий предок всех живых позвоночных на самом деле был тетрахроматическим, и что со временем млекопитающие утратили адаптацию цветового зрения. По словам Fitch, где-то в процессе у приматов — по крайней мере, у некоторых из нас — восстановилось своего рода частичное цветовое зрение.
Использование орудий — еще одна адаптация, которая многократно развивалась у разных видов животных. Шимпанзе, наши ближайшие живые родственники, ловят термитов с помощью инструментов и колют орехи. Шесть миллионов лет назад наш общий предок с шимпанзе, вероятно, также использовал простые инструменты для выполнения аналогичных задач. Посредством гомологии мы можем представить когнитивные способности наших вымерших предков.
Но приматы — не единственные животные, способные использовать инструменты. Новокаледонские вороны используют острые прямые предметы в своей среде, чтобы выкапывать труднодоступные личинки из стволов деревьев.Исследователи из Оксфордского университета обнаружили, что в лаборатории эти вороны будут делать свои собственные инструменты, сгибая кусочки проволоки в крючкообразные формы, чтобы черпать пищу из контейнеров.
«Это очень умные животные, и они действительно способны решать задачи и выходить за рамки своих биологических предрасположенностей так же, как мы, люди. Вот как мы можем водить автомобили и делать электродрели », — сказал Fitch.
Конечно, добавило Fitch, наш общий предок с воронами вряд ли был пользователем инструментов, но эта аналогичная адаптация позволяет нам начать задавать важные вопросы: почему использование инструментов развивается? И почему он развивался все это время и разными способами среди таких разных организмов?
Сигналы, синтаксис и семантика
Наряду с использованием инструментов люди разделяют многие когнитивные способности с другими видами, включая формирование воспоминаний, категорий, основных эмоций, таких как гнев, планирование и постановку целей, а также обучение правилам.Эти виды основных невербальных концепций, вероятно, предопределили язык за многие миллионы лет эволюции. В отличие от использования орудий, язык, по-видимому, является чертой, присущей только людям. Однако, по словам Fitch, большинство составных частей языка являются общими с другими видами.
«Основное отличие нас от других видов заключается не в том, что нам есть о чем подумать, а в том, что мы можем передавать то, о чем думаем», — сказал он.
Хотя некоторые шимпанзе и бонобо научились подписывать или общаться с помощью клавиатуры, никто так и не научился здороваться или петь «С Днем Рождения.«Это не потому, что шимпанзе не умны или не могут имитировать, а потому, что у них очень ограниченная способность контролировать свою вокализацию и имитировать звуки из окружающей среды.
Одна из давних гипотез неспособности приматов говорить заключается в том, что у них (и других животных) отсутствует опущенная гортань, которой обладают люди. Однако большая часть информации о гортани животных получена при вскрытии мертвых животных. В качестве постдока Fitch заинтересовалось тем, как общаются живые животные.Пока что все исследованные им млекопитающие опускают гортань до положения человека, издавая при этом громкие звуки; когда собака лает, гортань втягивается вниз только на мгновение лая, а затем снова поднимается.
«Что необычно в нас, так это не то, что у нас опущена гортань, а просто то, что она все время опускается, — поясняет Fitch.
Это исследование показывает, что для речи решающее значение имеет не анатомия голоса, а что-то в мозгу. Одна давняя гипотеза состоит в том, что у большинства млекопитающих есть только косвенные связи от их моторной коры с нейронами, которые контролируют голосовой тракт, гортань и язык.Люди тоже имеют эти нейронные связи, но также имеют прямые связи между моторной корой и моторными нейронами, которые контролируют гортань. Это ключ, который дает людям контроль над голосовыми путями, которых не хватает шимпанзе.
Однако люди — не единственные животные, способные изучать сложные вокализации; Обучение вокалу независимо развилось у летучих мышей, слонов, тюленей, китообразных и нескольких различных видов птиц. Изучая нейронные корреляты вокального обучения у самых разных видов, исследователи могут проверить эту гипотезу о прямых нейронных связях.На данный момент в ходе исследований были изучены две стаи птиц — певчие птицы и попугаи — и в обоих случаях гипотеза подтвердилась. Птицы, обладающие способностями к обучению вокалу, имеют эти прямые связи, в то время как птицы, которые не умеют учиться вокалу, такие как голуби или куры, нет.
«Мое общее заключение состоит в том, что использование очень широкой сравнительной точки зрения на широкий спектр различных видов животных действительно дает нам мощный инструмент как для разработки гипотез, так и для проверки этих гипотез», — поясняет Fitch.«Мы можем проверить как механистические, так и функциональные эволюционные гипотезы».
Синтаксис: сердце языка
Углубляясь в предмет общения, Fitch заявило, что синтаксис, набор правил, определяющих значение предложения, действительно лежит в основе языка. Помимо устного слова, люди могут использовать язык во многих формах: язык жестов и письмо, например, возможны благодаря нашей способности использовать расширенный синтаксис. Обезьяны могут не говорить, но они могут выучить и выразить сотни слов с помощью знаков или клавиатуры.Однако, несмотря на владение большим словарным запасом, уровень синтаксиса, который они получают, примерно такой же, как у двухлетнего ребенка — в основном, они могут соединять два слова. Хотя это очень ограниченный уровень синтаксиса, это все же синтаксис, так что есть что-то общее с человеческим языком.
Люди не интерпретируют язык как просто последовательность слов; что очень важно, мы можем интерпретировать эти последовательности как имеющие иерархическую структуру более высокого порядка. Fitch и его коллеги пытаются определить, какими языковыми компонентами обладают разные организмы, исследуя их способность изучать простые грамматические структуры по сравнению с более сложными.
Пока что сравнительные эксперименты показали, что эта способность использовать иерархический синтаксис может быть уникальной для людей. В одной серии экспериментов исследователи попытались научить иерархической грамматике двух разных видов птиц: голубей и кей. Кеас — это попугай, обитающий в Новой Зеландии, и они известны своей невероятной умностью. Вместо того, чтобы использовать записи речи, исследователи научили птиц распознавать различные визуальные паттерны абстрактных форм. Даже голуби — не самые умные птицы — смогли освоить простые последовательные шаблоны, но, хотя они прошли несколько недель интенсивного обучения, обе группы птиц не смогли выучить более сложную грамматику.
«Итак, где мы сейчас остаемся, так это: было показано, что множество различных видов животных выполняют грамматики с очень конечным числом состояний [и] более простые последовательные грамматики, но прямо сейчас единственное хорошее свидетельство выхода за рамки иерархической грамматики — это для люди », — сказал Fitch.
Что именно позволяет людям совершить этот языковой скачок? Fitch подозревает, что люди развили когнитивную склонность к выводам древовидных структур из последовательностей, которые трудны или невозможны для других животных.Согласно его гипотезе дендрофилии, уникальные способности человека к синтаксису проистекают из автоматической интерпретации последовательностей в ветвящиеся иерархические блоки. По мнению Fitch, чтобы перейти на следующий уровень грамматики, люди могли развить дополнительную форму абстрактной памяти, которая позволяет нам отслеживать фразы даже после того, как они закончились. Чтобы сделать возможным эту новую адаптацию, человеческий мозг, возможно, укрепил необходимые нейтральные структуры для обработки языка. Fitch отметило, что площадь области Брока у человека в семь раз больше, чем у шимпанзе, что делает ее наиболее расширенной областью человеческого мозга по сравнению с известными нам шимпанзе.Кроме того, у человека он гораздо более взаимосвязан с другими структурами мозга, чем у других приматов.
«Для меня наиболее интересная возможность снова связана с синтаксисом», — заключило Fitch. «У нас много общего, но относительно небольшая разница в архитектуре мозга сильно повлияла на когнитивные способности».
человек могут быть наиболее адаптивными видами
За 5 миллионов лет, прошедшие с тех пор, как первые гоминиды впервые появились из Рифт-Валли в Восточной Африке, климат Земли стал все более неустойчивым.В течение циклов, продолжавшихся сотни тысяч лет, засушливые районы центральной Африки были захвачены лесами, леса уступили место пастбищам, а прилегающие ландшафты были изрезаны глубокими озерами.
«Именно в контексте этого быстро меняющегося ландшафта люди развили свой значительный мозг и способность к адаптивному поведению», — сказал Рик Поттс, директор Программы происхождения человека в Национальном музее естественной истории Смитсоновского института. По его словам, в таком мире способность мыслить творчески, придумывать новые решения угроз выживанию оказывается важным преимуществом.
«Эволюция мозга — наиболее очевидный пример того, как мы эволюционируем, чтобы адаптироваться», — объяснил он. «Но в современную эпоху мы знаем, что в геноме человека есть все виды взаимодействий, которые позволяют человеческим организмам иметь пластичность — способность приспосабливаться сама по себе является эволюционной характеристикой».
Он сказал, что у человека есть два ключевых преимущества: наш мозг и наша способность к культуре.
«Наш мозг — это, по сути, социальный мозг», — добавил он. «Мы делимся информацией, создаем и передаем знания.Это средство, с помощью которого люди могут приспосабливаться к новым ситуациям, и это то, что отличает людей от наших более ранних предков, а наших более ранних предков от приматов ».
Эта адаптивная способность не только позволила нашим предкам преодолевать огромные колебания климата, но и впоследствии помогла им колонизировать новые среды обитания. Более ранний вид гоминидов Homo erectus обитал на большей части Африки и Азии. Между тем, Homo neanderthalensis — неандертальцы — заселили большую часть Европы.Наш собственный вид, Homo sapiens , расселился по еще более отдаленным уголкам земного шара, используя лодки, чтобы добраться до Австралии более 50 000 лет назад.
Виды, ушедшие в холода
«У вас было Homo sapiens , которые попадали в более холодную среду, чем могли вынести даже неандертальцы, в то же время, когда они мигрировали в пустыни, тропические леса, степи и ледниковую среду», — сказал Поттс. «Как этот тонкий, длинноногий гоминид смог выжить во всех этих различных средах, для меня это история о том, как вы становитесь приспосабливаемыми.«
Теория «отбора по изменчивости», которую Поттс впервые описал в 1996 году, относится не только к людям и их мозгу, но может быть применена к любым видам, переживающим периоды нестабильности окружающей среды. По словам Поттса, универсальные черты характера, такие как широкая диета, будут преимуществом в такие времена как для пастбищных животных, так и для их хищников.
«Все организмы должны быть в состоянии поддерживать гомеостаз в некотором диапазоне условий, которые не являются полностью стабильными», — объяснил Поттс.«Сам геном представляет собой развитую структуру, а это означает, что все формы жизни обладают некоторой степенью приспособляемости к ним».
Идея о том, что адаптивность сама по себе может быть эволюционной характеристикой, является относительно новой концепцией. Когда Поттс впервые описал свою теорию почти два десятилетия назад, генетики-эволюционисты встретили здоровую дозу скептицизма, понимая, что эволюция — это процесс приведения животных в соответствие с окружающей их средой.
Палеоантропологи, в том числе и Поттс, в значительной степени понимали, что люди эволюционировали в период постепенного перехода от более холодного и влажного климата к более засушливой среде.
Идея о том, что основные события в эволюции человека происходили не постепенно, а скачками в периоды повышенной климатической изменчивости, казалось, бросила вызов научному консенсусу. Но у теории выбора по изменчивости было одно большое преимущество: ее можно было проверить.
«У нас есть маркеры для различных важных событий в истории гоминидов — происхождения новых видов, разработки новых инструментов», — сказал Мэтт Гроув, профессор археологии, классики и египтологии Ливерпульского университета, который работал с Поттсом над моделированием. выбор вариабельности.«Если эти события совпадают с тем, что согласно климатическим данным, это были периоды нестабильности, это, казалось бы, поддерживает теорию Рика. И, в общем, так оно и есть».
Новое и зловещее испытание на горизонте
По его словам, происхождение каждого рода гоминидов, включая наш, находится в пределах того или иного окна климатической изменчивости.
«Мы видим, что в эти периоды появляются не только новые виды [гоминидов], но и новый образ жизни, жизни и взаимодействия с окружающей средой», — сказал Поттс.
Гроув объяснил, что большая ирония заключается в том, что способность взаимодействовать с окружающей средой вернула нас на траекторию климатической нестабильности. И на этот раз, вызванное искусственными парниковыми газами, глобальное потепление происходит намного быстрее, чем предыдущие сдвиги.
На заре сельского хозяйства 10 000 лет назад люди приступили к новому эксперименту — вместо того, чтобы адаптироваться к окружающей среде, мы начали адаптировать ее к нашим потребностям, вырубая и сжигая леса, чтобы освободить место для сельского хозяйства.Это, в свою очередь, дало больше свободного времени, больше общества и более свободный обмен информацией. По мере совершенствования культурных и технологических знаний мы смогли использовать энергию других животных, а со временем использовать огромную силу ископаемого топлива.
Несколько раз на протяжении истории человечества периоды климатической нестабильности вызывали ударные волны через устоявшиеся империи, такие как Аккадская империя в Месопотамии или средиземноморские империи бронзового века ( ClimateWire , август.16). Однако каждый раз вид приходил в норму, более успешный и адаптивный, чем когда-либо.
Сейчас, когда глобальное потепление происходит с головокружительной скоростью, человеческая адаптивность, вероятно, столкнется с самым большим испытанием, считает Гроув.
«Мы занимаемся проблемой изменения климата с тех пор, как оказались на Земле», — пояснил он. «Проблема, однако, в том, что это происходит сейчас за такой короткий промежуток времени. И поэтому очень трудно предсказать, сможем ли мы отреагировать и какой ценой.«
Перепечатано из Climatewire с разрешения Environment & Energy Publishing, LLC. www.eenews.net, 202-628-6500
Обычные виды рыб обладают «человеческой» способностью к обучению — ScienceDaily
Несмотря на разные миры, способ обучения рыб может быть ближе к человеческому мышлению, чем считалось ранее. предлагает новое исследование.
Обычный вид рыб, который встречается по всей Европе, включая Великобританию, называется девятииглая колюшка, может быть первым животным, демонстрирующим важную стратегию социального обучения человека.Согласно исследованию университетов Сент-Эндрюса и Дарема, колюшки могут сравнивать поведение других колюшек со своим собственным опытом и делать выбор, ведущий к лучшим запасам еды.
Исследователи предполагают, что у этих рыб может быть необычно сложная способность к социальному обучению, которая еще не обнаружена у других животных, что называется стратегией «восхождения на холм».
Эта способность выбирать участок корма лучшего качества путем сравнения того, насколько успешно другие получают еду из него, с их личным опытом ранее не проявлялась на животных, говорят ученые.
Команда исследователей предполагает, что в случае с девятииглой колюшей это, вероятно, будет «необходимостью», поскольку анатомия этого конкретного вида рыб не обеспечивает существенной защиты от хищников для безопасного кормления. Возможно, они были «вынуждены» узнавать от других, где кормиться, прячась от хищников, поскольку сами не могут рисковать искать места для еды на открытом воздухе.
Ученые говорят, что результаты, опубликованные в академическом журнале Behavioral Ecology , показывают, что когнитивные механизмы, лежащие в основе кумулятивной культурной эволюции, могут быть более распространены у нечеловеческих животных, чем это считается в настоящее время.Полученные данные показывают, что большой мозг, как у людей, не обязательно является предпосылкой кумулятивной культуры.
Исследователи говорят, что полученные данные способствуют пониманию эволюции мозга и типов мозга, необходимых для определенных когнитивных функций, как у людей, так и у животных.
Ведущий автор доктор Джереми Кендал из факультета антропологии Даремского университета и член Исследовательского совета Великобритании сказал: «У маленьких рыбок может быть маленький мозг, но они все же обладают некоторыми удивительными когнитивными способностями.
«Стратегии« восхождения на холмы »широко распространены в человеческом обществе, согласно которым достижения в области технологий сводятся к тому, что люди выбирают лучшую технику посредством социального обучения и совершенствуют его, что приводит к накоплению культуры.
«Но наши результаты показывают, что размер мозга — это еще не все, когда дело доходит до способности к социальному обучению».
Около 270 рыб были пойманы с помощью рыболовных сетей в Мелтон-Брук в Лестере и помещены в аквариумы в лаборатории. Рыбы были разделены на три экспериментальные группы и одну контрольную группу.Рыбам в экспериментальных группах было предложено два разных опыта обучения и два теста предпочтения в аквариуме с кормушкой на каждом конце.
Во-первых, они могли свободно исследовать кормушку на каждом конце во время ряда тренировочных испытаний, когда одна кормушка давала больше червей, чем другая, называемая богатой кормушкой. Затем их протестировали, чтобы определить, какую кормушку они предпочитают. Во втором тренировочном испытании те рыбы, которые научились отдавать предпочтение богатой кормушке, наблюдали за кормлением других рыб, но на этот раз богатые и бедные кормушки были поменяны местами с богатой кормушкой, что давало даже больше червей, чем та, которую рыба ранее получала. примерно столько же или меньше.Во втором тесте рыбы снова могли свободно плавать и выбирать кормушку.
Около 75 процентов рыб были «достаточно умны», чтобы понять, наблюдая за другими рыбами, что богатый кормушечный, который раньше на собственном опыте ощущал себя плохим кормушкой, принес им большую прибыль. Для сравнения, значительно меньшее количество рыб предпочли кормушку, которая казалась богатой, наблюдая за другими, если они сами имели опыт того, что альтернативная кормушка будет давать примерно такое же или большее количество корма.
Дальнейшие исследования команды показали, что вероятность копирования поведения других возрастает с увеличением скорости, с которой другие питались.
Доктор Кендал сказал: «Многие животные наблюдают за более опытными сверстниками и таким образом приобретают навыки поиска пищи, развивают пищевые предпочтения и учатся избегать хищников. Но просто копировать кого-то — не всегда рецепт успеха. Животные часто живут лучше. избирательно выбирают, когда и кого копируют.
«Эти рыбы, очевидно, не имеют никакого отношения к людям, но у них есть человеческая способность копировать только тогда, когда отдача лучше, чем их собственная.Вы могли ожидать эту способность у животных, которые тесно связаны с людьми. Что касается девятииглой колюшки, то они, скорее всего, адаптировались к местной экологии ».
Соавтор, профессор Кевин Лаланд из Школы биологии Университета Сент-Эндрюс добавил: «Девятииглая колюшка может быть гениями мира рыб. Примечательно, что форма обучения, признанная оптимальной у людей, — это именно то, что эти рыбы делать.»
Исследование проводилось в Университете Сент-Эндрюс, где доктор Кендал работал до прихода в Даремский университет в 2007 году.
Он финансировался Исследовательским советом по биотехнологии и биологическим наукам.
Факты о девятииглой колюше
вида колюшки обитают в пресных и морских водоемах Европы, Азии и Северной Америки, в том числе в заросших водорослями канавах и реках.
Питаются мелкими ракообразными и личинками рыб.
В сезон размножения самец строит гнездо, подвешенное на кусочке водорослей. Самка привлекается самцом, и она откладывает яйца внутри гнезда.Самец охраняет эти яйца и молодь, когда они вылупляются. Когда у молодых появляются шипы, он отгоняет их, чтобы они заботились о себе.
Могут ли люди обладать способностями животных?
Этот вопрос поднимает всевозможные интересные вопросы о том, кто мы как люди и как мы сравниваемся с другими животными.
Например, мы делимся с животными способностью использовать зрение, чтобы понимать окружающий мир, но то, как мы это делаем, сильно отличается. Благодаря нашему цветовому зрению мы можем видеть цвета в широком спектре.Но есть цвета, существующие в природе, которые мы не видим, в отличие от других животных.
Пчелы, например, могут видеть ультрафиолетовый свет, который находится за пределами видимого нами спектра. Вы думаете, что цветок выглядит определенным образом, но бабочка видит что-то другое, а пчела видит совсем другое.
Обоняние — это то, что мы разделяем с другими животными, но, опять же, то, как мы обоняем и насколько хорошо мы это делаем, сильно различается. Многие из нас знают, что собаки действительно хороши в носах.Хотя мы умеем пользоваться носом лучше, чем обычно думаем, собаки чуют то, чего мы никогда не слышим.
Еще одна способность, которую мы разделяем с животными, — это получение феромонов. Феромоны — это химические вещества, которые мы выделяем через жидкости организма, такие как пот, и, хотя их запах не осознан, они попадают в рецепторы в вашем мозгу и могут оказывать сильное влияние на ваше поведение. Это действительно важно, например, для насекомых и того, как они находят себе пару, а также для человека.
Есть много умных животных, таких как попугаи, дельфины и вороны, и мы так много узнаем об их интеллекте. Но факт в том, что люди действительно хороши в мышлении, планировании и разработке стратегии, и благодаря этому мы смогли создать машины, которые позволяют нам расширять наши возможности.
Мы знаем, что пчелы могут видеть цвета, которых нет у нас, потому что мы достаточно умны, чтобы создавать машины, которые позволяют нам видеть то, что видят они. Мы можем создавать технологии, которые позволяют нам слышать то, что делают другие животные, то, что мы не смогли бы услышать иначе.
Мы все представляем мир как единую реальность, то, что мы видим, слышим и обоняем. Но факт в том, что есть много опыта, в зависимости от того, к какому виду вы принадлежите.
Раскрытие эволюции уникального человеческого познания
Abstract
Удовлетворительное описание когнитивной эволюции человека объяснит не только психологические механизмы, делающие наш вид уникальным, но также то, как, когда и почему эти черты эволюционировали. На сегодняшний день исследователи достигли значительного прогресса в определении уникальных человеческих аспектов познания, но значительно меньше усилий было потрачено на вопросы об эволюционных процессах, в результате которых возникли эти черты.В этой статье я стремлюсь связать эти взаимодополняющие цели, синтезируя последние достижения в нашем понимании того, что делает человеческое познание уникальным, с теорией и данными, касающимися процессов когнитивной эволюции. Я рассматриваю доказательства того, что исключительно человеческое познание зависит от синергизма между репрезентативными и мотивационными факторами и вряд ли может быть объяснено изменениями какой-либо единственной когнитивной системы. Я утверждаю, что, хотя ни одно животное, не являющееся человеком, не обладает полным набором черт, определяющих человеческий разум, гомологии и аналогии критических аспектов человеческой психологии можно найти в различных нечеловеческих таксонах.Я предполагаю, что филогенетические подходы к изучению познания животных, которые могут ответить на вопросы о влиянии отбора и непосредственных механизмах, управляющих когнитивными изменениями, могут дать важную информацию о процессах, посредством которых эволюционировал когнитивный фенотип человека.
Человеческий разум кажется непохожим на разум любого другого вида. Мы участвуем в крупных организациях, ведем войны из-за убеждений, представляем себе далекое будущее и сообщаем об этих процессах, используя синтаксис и символы.Какие аспекты человеческого познания позволяют нам совершать эти, казалось бы, уникальные подвиги, и качественно ли эти процессы отличаются от процессов у других животных? Не менее важно, как и почему возникла такая своеобразная психология? Что такого особенного в образе жизни ранних людей способствовало этим гибким формам познания, и как естественный отбор создал эти черты из нечеловеческой обезьяноподобной основы? Приведенные выше вопросы касаются различных уровней объяснения (1, 2) когнитивной уникальности человека, но в конечном итоге удовлетворительное описание когнитивной эволюции человека объяснит не только механизмы, которые делают наш вид уникальным, но также как, когда и почему эти черты эволюционировали. .На сегодняшний день ученые добились значительного прогресса в определении уникальных человеческих аспектов познания, но значительно меньше усилий было потрачено на вопросы об эволюционных процессах, в результате которых возникли эти черты. В этой статье я стремлюсь связать эти уникальные, но взаимодополняющие цели, сначала выделив недавние достижения в нашем понимании того, чем человеческая психология отличается от психологии других существующих таксонов. Затем я перехожу к менее понятным вопросам о том, как, когда и почему эти черты эволюционировали, и подчеркиваю важность понимания эволюционных процессов, а не только их продуктов, для всестороннего понимания когнитивной эволюции человека.
Что делает человеческое познание уникальным?
На первый взгляд когнитивные различия между людьми и всеми другими животными кажутся огромными. Только люди выполняют вычисления, путешествуют на машинах с глобальными системами позиционирования, ищут жизнь за пределами нашей планеты и хранят информацию о том, как это сделать, в цифровых репозиториях, доступных по всему миру. Но ни один из этих умений не запрограммирован в человеческом мозгу, и ни один из них не был изобретен de novo одним предприимчивым человеком.Напротив, все эти достижения зависели от наращивания тысячелетий постепенного прогресса и когнитивной и культурной системы, которая позволяла (и мотивировала) людей приобретать и передавать накопленные знания и навыки (3). Томаселло и Ракоци (4) подчеркивают этот момент, отмечая, что «если мы представим человеческое дитя, рожденное на необитаемом острове, каким-то волшебным образом сохранившее жизнь само по себе до взрослого возраста, возможно, что когнитивные навыки этого взрослого не будут сильно отличаться — возможно, немного, но не очень — по сравнению с другими человекообразными обезьянами (исх.4, стр. 121) ». Следовательно, существует важное различие между попыткой понять, как совокупная культура повлияла на коллективное познание нашего вида, и попыткой понять, как люди в первую очередь стали такими плодовитыми культурными существами.
Чтобы понять происхождение человеческого культурного познания, ученые обратили внимание на человеческое развитие и фундаментальные аспекты человеческого познания, которые позволяют нам общаться, делиться и получать информацию от других (5).Эти процессы возникают на ранних этапах онтогенеза человека и поддерживаются зарождающимся пониманием других как преднамеренных агентов. В течение первого года жизни человеческие дети начинают относиться к другим по-новому, настраиваясь на внимание других посредством таких процессов, как отслеживание взгляда и обмен информацией с другими посредством простых актов референциального жеста (6). Эти базовые навыки общения и совместного внимания обеспечивают социальную основу для различных форм культурного обучения, включая начальные этапы овладения языком (7, 8).Например, к 2 годам эти навыки перспективного восприятия позволяют человеческим детям делать прагматические выводы, связывающие новые слова с (предполагаемой) целью чужого внимания (9). Таким образом, уже в первые годы жизни человеческие дети начинают познавать мир не только своими глазами, но и вместе с другими, и эти способности рассуждать о чужом сознании — все вместе именуемые «теорией разума» — обеспечивают детям с мощными механизмами для получения и обмена культурной информацией, включая язык, социальные нормы и социальные убеждения.Примерно к четырем годам человеческие дети достигают еще одной вехи в понимании других как намеренных агентов, явно интерпретируя поведение других как результат психологии убеждений и желаний, а также рассуждая о целях и убеждениях не только других людей, но и своей культурной группы в более широком смысле (4). Недавние кросс-культурные исследования показывают, что эти рано появляющиеся навыки рассуждений о чужом сознании развиваются примерно в одном возрасте в разных культурах (10, 11) и представляют собой важные вехи на пути к уникальному человеческому культурному познанию.Конечно, овладение символическим языком еще больше продвигает человеческое познание, возможно, предоставляя новую репрезентативную среду, которая допускает новые формы абстрактного и реляционного мышления, а также беспрецедентные формы коммуникативной гибкости (12–15). Однако критическим моментом является то, что овладение и использование человекоподобного языка просто невозможно и не полезно для вида, не имеющего необходимых навыков для рассуждений о других разумах. Являются ли эти фундаментальные социокогнитивные навыки уникальными для людей?
До недавнего времени существовало общее мнение, что люди действительно уникальны в своем понимании других как преднамеренных агентов (16, 17).Однако исследования за последние 15 лет поставили эту черно-белую интерпретацию под сомнение (но см. Ссылку 18), обнаружив, что некоторые нечеловеческие виды действительно используют информацию о восприятии, знаниях и намерениях других (19). Пересмотренное представление о нечеловеческой теории разума развивалось параллельно с изменениями в экспериментальных методах, которые теперь делают упор на исследованиях животных в более экологически значимых условиях. Например, первые положительные доказательства того, что у ближайших живых родственников людей есть перспектива, появились, когда шимпанзе тестировались в конкурентных ситуациях с сородичами, а не в контексте сотрудничества с людьми (20, 21).Со времени этих первоначальных исследований несколько видов, помимо обезьян, продемонстрировали аналогичные навыки в контексте социальной конкуренции (22⇓ – 24). Эта социально-когнитивная гибкость в конкурентных, но не кооперативных контекстах хорошо согласуется с гипотезой «Макиавеллистского интеллекта», которая предполагает, что разум приматов сформировался в результате эволюционной гонки вооружений, в которой навыки манипулирования и обмана других имели первостепенное значение (25, 26).
Если способность рассуждать о том, что думают другие люди, не уникальна только для людей, то чем же объясняются уникальные человеческие формы культуры и общения, которые начинают проявляться в первые дни рождения детей? Одно из объяснений делает особый акцент на сочетании навыков взгляда на перспективу и совместной мотивации для обмена психологическими состояниями с другими, включая совместные цели и намерения (27).В отличие от нечеловеческих обезьян, которые используют точки зрения других в первую очередь для своих собственных целей (28), человеческие младенцы применяют свои навыки восприятия точки зрения в контексте разделения внимания с другими и совместного общения друг с другом. Важно отметить, что человеческие дети также ожидают, что их социальные партнеры будут иметь такую же мотивацию, создавая основу для взаимного сотрудничества для коммуникативных и совместных усилий. Например, в первые дни своего рождения человеческие дети начинают показывать указывающие жесты, просто чтобы привлечь внимание других к интересующим объектам, и, когда другие указывают на них, дети предполагают, что мотив сотрудничества имеет отношение к общей почве между двумя коммуникаторами (29 ).Напротив, в то время как человекообразные обезьяны могут научиться императивно указывать, например, при запросе еды (30), они не производят указывающих жестов просто для того, чтобы поделиться информацией с другими, и, когда другие указывают на них сообща (например, чтобы указать местоположение скрытая еда), обезьяны, как правило, плохо работают, скорее всего, из-за того, что они не понимают намерения своего партнера к сотрудничеству. Вскоре после 1 года человеческие просоциальные и кооперативные мотивы начинают проявлять себя более явно через акты (незапрошенной) инструментальной помощи, которые снова критически поддерживаются способностью делать выводы о намерениях, знаниях и желаниях других (31).Следовательно, в отличие от нечеловеческих обезьян, человеческое познание, по-видимому, больше приспособлено для совместных и просоциальных целей, чем для целей Макиавелли (32).
Важно отметить, что ни понимание других как намеренных агентов, ни просоциальные и кооперативные установки сами по себе не могут поддерживать уникальное человеческое культурное познание. Скорее, это синергия между мотивациями к участию в совместной деятельности с общими целями и психологическими процессами для представления лежащей в основе интенциональности «мы» (33), которая позволяет людям создавать культурные продукты, которые так существенно отличаются от продуктов других видов (34). .Как и почему тогда возникло это необычное созвездие черт? Были ли эти мотивационные и репрезентативные изменения эволюционно связаны или они имеют независимое эволюционное происхождение?
Стать человеком
Попытки реконструировать когнитивную эволюцию человека за последние 5–7 миллионов лет назад требуют выводов о характеристиках нашего последнего общего предка (LCA) с нашими ближайшими живыми родственниками — бонобо и шимпанзе. Поскольку бонобо и шимпанзе в равной степени связаны с людьми, но отличаются друг от друга морфологически, поведенчески и когнитивно, велись активные дебаты относительно того, какой (если какой-либо из двух видов) из двух видов Pan является лучшей живой моделью нашего LCA ( 35⇓⇓ – 38).Хотя нет четкого консенсуса по этому вопросу, исследования, сравнивающие развитие черепа у человекообразных обезьян, показывают, что бонобо отклоняются от консервативного паттерна, обнаруженного у горилл и шимпанзе, и указывают на то, что бонобо могут быть относительно производными из-за неотенического развития (39). Считается, что эти изменения в сроках развития оказали каскадное воздействие на различные аспекты биологии бонобо, что привело к производным аспектам социального поведения и познания, которые отличаются от шимпанзе и, предположительно, также от LCA бонобо, шимпанзе и человека (37). , 40).
Если предположить, что последний общий предок был более похож на шимпанзе, эволюционные изменения темперамента и, в частности, сдвиг в сторону более терпимого и менее агрессивного социального поведения, возможно, были критическими предпосылками для эволюции человеческих форм культурного поведения и познания. У диких шимпанзе внутригрупповая агрессия как среди самцов, так и среди самок может быть экстремальной, а межгрупповая агрессия часто приводит к летальному исходу (41, 42). Хотя шимпанзе сотрудничают в таких контекстах, как групповая охота, успешный похититель обычно сохраняет большую часть добычи и предоставляет обрывки другим преимущественно в ответ на преследование (43).Следовательно, хотя в некоторых контекстах они являются умелыми сотрудниками, кажется, что шимпанзе мотивированы в первую очередь индивидуальными целями, являются относительно нетерпимыми партнерами по совместному использованию и мало заботятся о справедливом распределении ресурсов, возникающих в результате совместных усилий (44). Сдерживающая роль социальной толерантности в сотрудничестве и познании шимпанзе также хорошо подтверждена экспериментальными исследованиями. Например, шимпанзе более успешны в задачах инструментального сотрудничества, когда вознаграждение физически рассредоточено, чем когда оно сгруппировано и индивидуально монополизируется (45), а нетерпимость между людьми может препятствовать социальному обучению в парадигмах модель – наблюдатель (46).Следовательно, социальная нетерпимость между людьми может представлять собой эмоциональный барьер, который значительно снижает потенциал сотрудничества и социального обучения.
По сравнению с шимпанзе бонобо, которые в равной степени родственны людям, характеризуются менее интенсивными формами агрессии как внутри социальных групп, так и между ними (47). Бонобо питаются друг с другом более терпимо, чем шимпанзе (ссылка 48; но см. Ссылки 49 и 50), и добровольно делятся пищей с другими особями (51) (рис. 1), включая незнакомцев (52).Эта терпимость и готовность делиться позволяет бонобо превосходить шимпанзе в некоторых совместных задачах, особенно когда вознаграждение потенциально монополизировано (48). Предполагается, что относительно более низкий уровень агрессии и повышенная социальная терпимость у бонобо по сравнению с шимпанзе являются результатом процесса «самодомашнивания» (40). Эта гипотеза подтверждается данными, раскрывающими синдром поведенческих, морфологических и психологических черт бонобо, сходных с таковыми у искусственно одомашненных видов.Считается, что совпадение этих черт у бонобо и одомашненных видов является результатом отбора (естественного или искусственного) против агрессии, который привел к изменениям в сроках развития и нейрофизиологии, включая изменения оси гипоталамус-гипофиз-надпочечники, уровней андрогенов и серотонинергическая система (40, 53).
Рис. 1.Два бонобо делят фрукт. Бонобо более терпимо делятся едой, чем шимпанзе, что позволяет им успешно сотрудничать, даже когда вознаграждение потенциально монополизировано.Отбор, ориентированный на повышенную социальную терпимость и заботу о справедливом распределении ресурсов, вероятно, был важным предшественником крупномасштабного сотрудничества среди людей. Изображение предоставлено Цзинчжи Таном (фотографом).
Почему естественный отбор мог благоприятствовать этим поведенческим изменениям у бонобо, но не у шимпанзе? Одно правдоподобное объяснение подчеркивает различия в экологии кормления между регионами к северу и югу от реки Конго (54). В то время как шимпанзе и гориллы разделяют среду обитания и соревнуются за растительность к северу от реки, бонобо ограничены регионами к югу от реки, которые не пересекаются с таковыми шимпанзе или горилл.Из-за высокого уровня конкуренции самки шимпанзе прибегают к поиску пищи в одиночку, не образуют прочных союзов с другими самками и уязвимы для сексуального принуждения со стороны самцов (55–57). Напротив, самки бонобо сталкиваются с более низким уровнем конкуренции в схватке и образуют союзы с другими самками, которые могут эффективно сдерживать принуждение самцов, а самцы бонобо получают выгоду за счет аффилированного, а не агрессивного поведения по отношению к самкам (47, 58). В этих условиях отбор, вероятно, отдавал предпочтение самцам со сниженной склонностью к агрессии, что в конечном итоге привело к самодомашниванию фенотипа (40).
Данные по искусственно прирученным видам предоставляют дополнительные доказательства того, как снижение агрессии может потенциально трансформировать не только темперамент, но и аспекты социального познания, включая некоторые коммуникативные навыки сотрудничества, которые считаются критически важными для когнитивного развития человека. Например, сравнение одомашненных видов и их диких предков, включая собак и волков, экспериментально одомашненных серебристых лисиц и представителей контрольной линии, а также домашних и диких хорьков, показывает, что одомашненные формы проявляют повышенную чувствительность к совместному общению, а также изменения в других социальное поведение, такое как готовность поддерживать зрительный контакт (59–62).Важно отметить, что эти изменения в кооперативном поведении, как считается, возникли как побочный продукт изменений темперамента (63), демонстрируя, как эмоциональная эволюция может снять ограничения социальной терпимости и эффективно разрешить новые формы социального взаимодействия (64). Был ли этот тип трансформации темперамента также ключевым шагом в когнитивной эволюции человека? Прав ли был Дарвин, предположив, что «[m] an во многих отношениях может быть сравнен с теми животными, которые уже давно одомашнены» (ссылка 65, с.172)?
Одной из основных морфологических тенденций в эволюции человека было уменьшение полового диморфизма как массы тела, так и размера клыков. Сравнительный анализ с современными приматами связывает эти морфологические черты с различиями в социальных системах и предполагает, что высокие уровни диморфизма связаны с интенсивной конкуренцией самцов и самцов и полигинными системами спаривания (66). Таким образом, снижение полового диморфизма в человеческой линии может отражать переход от более похожей на шимпанзе системы спаривания с высоким уровнем насилия между самцами и сексуальным принуждением самок к моногамии и кооперативному размножению.Этот сдвиг в социальных системах, который, возможно, начался еще 3 миллиона лет назад (67). –, вероятно, способствовал первоначальному снижению агрессии и повышению терпимости между людьми (68). В течение последних 200000 лет дополнительные изменения в морфологии черепно-лицевой морфологии человека поднимают интригующую возможность второй волны отбора против агрессии, которая совпала с появлением модерна поведения (69). В частности, Cieri et al. задокументировано усиление феминизации черепов человека от среднего плейстоцена до наших дней, о чем свидетельствует уменьшение выступа надбровных дуг и укорочение верхней части лица (70).Предполагается, что эти анатомические изменения являются результатом снижения активности андрогенов и согласуются с хорошо задокументированными эффектами тестостерона на черепно-лицевую маскулинизацию. Учитывая, что тестостерон связан с доминированием и агрессивным поведением у мужчин (71), эти изменения могут отражать недавнюю волну отбора в пользу повышенной социальной терпимости, которая позволила людям продуктивно работать с сородичами новыми способами (72), как предполагает распространение культурных артефактов 20–70 тыс. лет назад (тыс. лет назад) (69, 73, 74).
Помимо воздействия на темперамент, снижение уровня тестостерона могло также напрямую повлиять на аспекты человеческого познания, включая процессы, связанные с общением и теорией разума. Доказательства этой возможности вытекают из теории систематизации и сочувствия, разработанной Бароном-Коэном и др. (75) и предложили в качестве объяснения недостатков теории разума, наблюдаемых при расстройстве аутистического спектра (76). Короче говоря, теория предполагает, что когнитивные различия между мужчинами и женщинами частично возникают из-за различного пренатального воздействия андрогенов.Барон-Коэн рассматривает доказательства того, что на уровне популяции мужчины превосходят женщин в задачах, связанных с зрительно-пространственными и пространственно-временными способностями (77), предположительно отражающими «систематизирующие» навыки или тенденцию анализировать мир с точки зрения законных и детерминированных правил (78). ). Напротив, женщины демонстрируют преимущество в речи (77) при интерпретации выражений лица и участии в интерактивном социальном обмене, включая обмен мнениями и смену очереди, а девочки достигают определенных этапов развития теории разума раньше, чем мальчики (79).Хотя некоторые из этих эффектов могут быть обусловлены культурой, аналогичные различия были зарегистрированы у нечеловеческих животных, некоторые из которых уменьшаются при кастрации самцов или искусственной андрогенизации самок (78). Гипотеза систематизации и сочувствия предполагает, что высокие уровни воздействия андрогенов во время пренатального развития могут вызвать крайнюю маскулинизацию мозга, что приводит к (патологическому) уклону к систематизации и приводит к дефициту эмпатических процессов, таких как теория разума.Барон-Коэн и его коллеги предполагают, что расстройства аутистического спектра, которые характеризуются дефицитом в теории психики, вызываются «исключительно мужским мозгом», но также приводят данные, что даже у типично развивающихся людей пренатальное воздействие андрогенов является прогностическим признаком аутистических черт. измеряется позже в детстве (80).
Хотя гипотеза систематизации и сочувствия была предложена для объяснения внутривидовых вариаций у современных людей, ее предсказания могут также частично объяснять видовые различия в социальном познании человекообразных обезьян и ключевые изменения в когнитивной эволюции человека.Например, бонобо, которые демонстрируют более низкую пренатальную экспозицию андрогенов, чем шимпанзе (81), превосходят шимпанзе по некоторым показателям теории разума и сотрудничества (48, 82), обращают внимание на лицо и глаза чаще, чем шимпанзе при просмотре социальных изображений. (83) и чаще делятся едой и играют в обществе, будучи взрослыми, чем шимпанзе (ссылки 48 и 84; но см. Ссылки 49 и 50) — все эти черты связаны с сочувствием. Напротив, шимпанзе превосходят бонобо в тестах на использование инструментов, причинное мышление и пространственную память (82, 85) — когнитивные черты, связанные с систематизацией — а шимпанзе демонстрируют более серьезную агрессию, чем бонобо (47), что согласуется с предсказаниями систематизации — сопереживающая гипотеза (рис.2). Сравнительные исследования изображений мозга шимпанзе и бонобо также показывают, что у бонобо больше серого вещества в областях, связанных с эмпатией, и более устойчивые нервные пути, связанные с подавлением агрессии (86), причем у бонобо примерно в два раза больше серотонинергической иннервации миндалины по сравнению с шимпанзе (53).
Рис. 2.Когнитивные и поведенческие различия между шимпанзе и бонобо, которые совпадают с предсказаниями гипотезы систематизации и сочувствия.B, бонобо; C, шимпанзе.
Если предположить, что предки человека были наделены базовыми навыками взгляда на перспективу, присущими другим человекообразным обезьянам, снижение активности андрогенов в недавней эволюции человека могло быть катализатором как для развития этих способностей, так и для их применения в новых типах сотрудничества. социальные взаимодействия. Следовательно, в этом сценарии критические аспекты типичной для человека социальной мотивации (например, толерантность и общительность) и социального познания (например, аспекты теории разума) могли развиваться параллельно, отчасти из-за схожих биологических механизмов, регулирующих оба набора. черт.
Проверка гипотез о когнитивной эволюции
Описанные выше эволюционные сценарии являются спекулятивными и основаны на данных относительно небольшого (но филогенетически информативного) таксонов, чтобы сделать выводы о ключевых процессах когнитивной эволюции человека. Хотя человеческое познание развивалось только один раз и представляет собой конечный продукт бесчисленных постепенных эволюционных изменений, сравнительные исследования с животными, не относящимися к человеку, дают возможность явно проверить гипотезы о том, как и почему некоторые из этих изменений могли произойти (2).На сегодняшний день немногие исследования использовали этот подход — в основном из-за проблем, связанных с составлением больших и высококачественных межвидовых наборов данных о познании животных, — но несколько недавних исследований иллюстрируют ценность явно сравнительных подходов для исследований когнитивной эволюции. Например, MacLean et al. (87) провели тесты самоконтроля у 36 видов позвоночных и обнаружили прочную связь между абсолютным (но не относительным) размером мозга и навыками самоконтроля. Эти данные повышают вероятность того, что увеличение абсолютного размера мозга, определяющее свойство эволюции человека, могло привести к улучшенным способностям к саморегуляции, возможно, поддерживая возросшую социальную терпимость (например,g., через подавление агрессии), что имеет решающее значение для человеческого сотрудничества. Интересно, что в этом исследовании не было выявлено связи между типичным для вида размером социальной группы и самоконтролем, что позволяет предположить, что простого проживания в более крупных социальных группах недостаточно для развития этих способностей. Однако в другом эксперименте с лемурами MacLean et al. (88) обнаружили, что более крупные типичные для вида социальные группы были связаны с повышенными навыками, необходимыми для восприятия зрительной перспективы (ключевой компонент теории разума), подтверждая гипотезу о том, что жизнь в сложных социальных группах является движущей силой когнитивных навыков, которые позволяют люди, чтобы превзойти других в доступе к пище и партнерам.Следовательно, жизнь в более крупных и сложных социальных сетях, возможно, способствовала первоначальной эволюции некоторых компонентов теории разума, которые у людей были перепрофилированы в новых контекстах сотрудничества. Что касается эволюции человеческих кооперативных и просоциальных мотивов, Burkart et al. (89) протестировали 15 видов приматов в серии проактивных задач просоциальности и исследовали ряд социоэкологических предикторов видовых различий. В этом случае степень материнской заботы (забота о детенышах, оказываемая не матерями) была лучшим предиктором видовых различий в просоциальном поведении.Эти результаты согласуются с идеей о том, что переход от полигамии к кооперативному разведению мог иметь решающее значение для эволюции уникальных человеческих форм кооперативной психологии (90). Важно отметить, что поскольку ранние люди, скорее всего, уже были наделены многими компонентами теории разума, мотивационные изменения, сопровождающие кооперативное размножение, могли послужить катализатором для применения этих навыков в условиях сотрудничества, в которых общая интенциональность стала адаптивной (68).Напротив, у других совместно размножающихся приматов (например, каллитрихидов) могут отсутствовать общечеловеческие общие намерения, потому что они обладают мотивационной, но не репрезентативной основой для этих процессов, и наоборот для современных человекообразных обезьян, обладающих некоторыми необходимыми репрезентативными способностями. но может не обладать уровнем просоциальной мотивации, присущей кооперативным заводчикам (68).
Приведенные выше исследования подчеркивают полезность филогенетических сравнительных подходов для вывода о том, как и почему развиваются определенные аспекты психологии, включая психологические черты, которые считаются критическими для когнитивного фенотипа человека.Тем не менее, таких исследований было проведено мало, и многие из гипотез, изложенных в этой статье, все еще созрели для сравнительного исследования. Например, зависит ли социальная терпимость и навыки сотрудничества или социального обучения от нечеловеческих видов? Близкородственные таксоны, которые существенно различаются по социальной толерантности, например, радиация макак (91), предоставляют мощные возможности для оценки того, могут ли эти черты быть функционально связаны. Точно так же, если влияние пренатального воздействия андрогенов приводит к систематическим различиям в познании, связанным с систематизацией и сочувствием, можно предсказать, что эти эффекты должны быть очевидны в ряде таксонов за пределами человекообразных обезьян.Что касается вопросов такого типа, то сравнительные исследования будут иметь решающее значение для оценки того, совпадают ли гипотезы о когнитивной эволюции человека с закономерностями, наблюдаемыми в других таксонах. Нет сомнений в том, что человеческое познание уникально и состоит из совокупности черт, которые в совокупности могут не совпадать ни с одним другим видом. Однако многие важные аспекты человеческого познания имеют гомологии — и часто, что более интересно, аналогии (в результате конвергентной эволюции) — в других таксонах, что создает широкие возможности для выводов о том, когда, как и почему эти черты развиваются.
Наконец, в тех случаях, когда аспекты познания человека кажутся радикально отличными от познания других видов, можно использовать филогенетические подходы для оценки того, следует ли рассматривать человека в качестве эволюционного выброса (92, 93). Например, недавний анализ показал, что, несмотря на наличие гораздо большего числа нейронов, чем у любого другого мозга приматов, количество нейронов в мозге человека не является значительным, учитывая объем человеческого мозга и данные об общих правилах клеточного масштабирования мозга приматов (94 , 95).Аналогичный анализ может быть проведен с когнитивными характеристиками, чтобы оценить, представляют ли явно необычные наблюдения у людей крайнее, но предсказуемое явление, принимая во внимание филогению приматов и набор переменных-предикторов, которые зависят от интересующего признака в разных таксонах. Когда человеческие черты можно частично объяснить более широкими эволюционными паттернами, сравнительные подходы будут особенно полезны для ответа на вопросы о том, как, когда и почему эти черты эволюционировали у людей.В случаях, когда люди существенно отклоняются от широкомасштабных эволюционных паттернов, эти находки предполагают, что люди могут считаться эволюционными выбросами (96) и требуют большой осторожности при реконструкции того, как и почему эти аспекты человеческого познания могли развиться.
Выводы
Люди — необычные животные во многих отношениях, но именно в нашем типичном для вида познании человеческая уникальность проявляется наиболее ярко. Точные способы, которыми человеческое познание отличается от познания других видов, остается предметом интенсивных дискуссий (14), но многие данные в настоящее время подтверждают гипотезу о том, что это ранний набор социальных навыков для рассуждений о сородичах как о намеренных агентах в сочетании с отчетливо кооперативная и просоциальная мотивация, которая питает многие из наших самых замечательных когнитивных достижений (97).Хотя научная литература изобилует попытками определить отдельные способности, которые делают человеческое познание уникальным, вполне вероятно, что человеческое познание — это больше, чем сумма его частей, и зависит от синергии между уникальной комбинацией репрезентативных и мотивационных черт. Следовательно, хотя человеческое познание в целом не имеет себе равных в животном царстве, ключевые компоненты нашего когнитивного фенотипа можно найти в других таксонах, включая не только человекообразных обезьян, но и более отдаленные виды, имеющие когнитивное сходство с людьми в результате конвергентная эволюция.Соответственно, новые направления исследований, объединяющие филогенетические сравнительные методы с изучением разума животных, будут играть важную роль в нашем стремлении определить не только то, что делает человеческое познание уникальным, но также то, как и почему эти черты эволюционировали.
Благодарности
Я благодарю Б. Хэра, Дж. Тана, К. Крупенье и двух анонимных рецензентов за полезные комментарии к более раннему черновику этой рукописи, Дж. Тана за предоставление фотографии на рис. 1 и Фонд Стэнтона. за финансовую поддержку во время написания рукописи.
Сноски
Автор: E.L.M. написал газету.
Автор заявляет об отсутствии конфликта интересов.
Эта статья представляет собой прямое представление PNAS.
Границы | Есть ли различия в «интеллекте» нечеловеческих видов? Роль контекстных переменных
Претензия Макфейла
Таким образом, в данном случае мы должны сделать вывод об отсутствии качественных или количественных различий среди позвоночных (за исключением человека; Macphail, 1985, p.46).
В 1985 году Макфейл отстаивал нулевую гипотезу для интеллекта животных, в которой говорилось, что нет никаких качественных или количественных различий в интеллекте между нечеловеческими видами. Позже Макфейл опубликовал свою Нулевая гипотеза в качестве целевой статьи в Behavioral and Brain Sciences (Macphail, 1987). Справедливо сказать, что комментарии коллег были в целом негативными. Например, Штернберг (1987, стр. 680) заявил, что «Макфейл предпринял смелые, но не полностью успешные усилия», в то время как Элепфандт (1987, стр.662) прокомментировал недавно появившееся исследование интеллекта позвоночных, что «этот новый рост не должен останавливаться узкими взглядами или поспешными выводами». Возможно, самый резкий комментарий был сделан Голдман-Ракичем и Прейссом (1987, с. 667), которые заявили, что «нулевая гипотеза Макфейла — это просто эпитафия на краеугольном камне сравнительного познания».
Вместо того, чтобы остановить рост сравнительного познания или стать эпитафией на его надгробии, за более чем три десятилетия после публикации Разум позвоночных: нулевая гипотеза (Macphail, 1985) произошел взрыв исследований в области когнитивных способностей. вместимости животных.Такие темы, как эпизодическая память , теория разума и планирование будущего , мало исследовались в 1985 году, тогда как сейчас они составляют основу исследований когнитивных функций животных. И другие темы, такие как представление отношений эквивалентности (рефлексивность, симметрия и транзитивность), имеют долгую историю исследований и продолжают способствовать глубокому пониманию умственных способностей нечеловеческих животных. В свете большого количества данных, накопленных с тех пор, как Макфейл (1985) опубликовал свою нулевую гипотезу , цель этой статьи — выяснить, выдержала ли она проверку временем: действительно ли нет качественных или количественных различий в познавательные способности позвоночных животных?
Некоторые общие проблемы
В данной статье мы рассматриваем текущее состояние утверждения Макфейла об отсутствии качественных или количественных различий в интеллекте между нечеловеческими видами позвоночных.Многие критические замечания в адрес Макфейла (1987) касались использования им термина «интеллект». Например, Барлоу (Barlow, 1987, p. 657) прекрасно выразил это, когда заявил, что «поскольку еще не существует общепринятого определения интеллекта, позволяющего определить для него количественную шкалу… нельзя с полным основанием сказать, что количественные различия либо существуют или не существуют ». Мы считаем, что это справедливая критика, и, кроме того, согласны с Ходосом (1987, с. 668), когда он заявил, что «мы не должны увязать в общей концепции интеллекта животных, потому что ее измерение находится далеко за пределами нашей досягаемости.”
Помимо ограничений в определении «интеллекта», область сравнительного познания заключается в сравнении способностей разных животных, чтобы понять не только их способности, но и эволюцию умственных способностей людей. Однако оценка того, как животные различаются по «интеллекту», может быть не лучшим подходом. Скорее, мы думаем, что лучший подход — сосредоточиться на конкретных, определяемых и измеримых возможностях, которые позволяют проводить прямые сравнения между видами.Д’Амато и Салмон (1984, стр. 149) выдвинули такую точку зрения относительно сравнения когнитивных способностей разных видов, когда они сказали, «насколько проще была бы задача, если бы мы могли идентифицировать относительно небольшое количество ядерных когнитивных способностей». это позволило бы нам с помощью их измерений делать разумные утверждения о когнитивных потенциалах и способностях различных видов ». В этой степени мы сосредоточимся на наборе таких ядерных когнитивных способностей, которые были предметом обширных исследований у разных видов: рефлексивность, симметрия, транзитивность и поведение последовательного порядка, а также коснемся некоторых более современных ядерных когнитивных способностей, таких как эпизодическая память и ToM.
Помимо проблем, связанных с использованием термина «интеллект», есть еще одно предостережение, касающееся использования нами термина «познание». Мы используем этот термин больше для простоты изложения, чем обязательно для обозначения того, что наши животные решают задачи, используя процессы, которые выходят за рамки поведенческих принципов, охватываемых оперантными и классическими обусловливающими или ассоциативными процессами, или не объясняются ими. Фактически, использование нами термина «познание» является синонимом определения Шеттлворта (1998, стр. 5), согласно которому он охватывает «механизмы, с помощью которых животные получают, обрабатывают, хранят и действуют на основе информации из окружающей среды.”
Третье предостережение заключается в том, что мы не собираемся сравнивать когнитивные способности нечеловеческих животных с человеческими. Такие сравнения подробно рассматривались в недавнем обзоре (Penn et al., 2008). Скорее, наша цель — сравнить когнитивные способности нечеловеческих животных и, в частности, рассмотреть значение нулевой гипотезы . Точно так же, поскольку большинство исследований проводилось на обезьянах, обезьянах, крысах или птицах, наши сравнения ограничиваются этими видами.Тем не менее, эти виды обладают достаточным диапазоном эволюционной независимости, а также различиями в нейроанатомии и нишах, чтобы предотвратить любую критику, которую мы не смогли широко проанализировать.
Роль контекстных переменных
В 1965 году Биттерман выдвинул идею контекстной переменной, некогнитивного фактора, который объясняет различия в наблюдаемом поведении между видами. Говоря о неспособности рыб продемонстрировать улучшения в обратном обучении как при решении пространственных, так и визуальных задач, Биттерман (1965, стр.95) заявил, что:
«Другая возможность состоит в том, что условия, в которых тестировалась рыба, виноваты в ее плохой демонстрации, что разница в производительности должна быть связана не только с разницей в возможностях, но и с неравенством в некоторых контекстных переменных, таких как как сенсорная потребность, двигательная потребность, степень голода или привлекательность вознаграждения ».
Биттерман (1965, стр. 95) также предвидел проблему с понятием контекстных переменных, когда заявил: «Можем ли мы когда-нибудь исключить возможность того, что разница в производительности двух разных животных в таком эксперименте проистекает из разницы в какая-то запутанная контекстная переменная? » Макфейл (1985, стр.39) пересмотрел понятие контекстных переменных в своей статье и повторил ту же озабоченность, когда заявил, что «не существует конечного каталога потенциально релевантных контекстных переменных: как, следовательно, можно окончательно исключить их влияние?»
Хотя озабоченность по поводу контекстных переменных разумна, мы полагаем, что контекстные переменные действительно находятся в континууме важности и релевантности. Хотя это может быть оправдано, мало кто будет склонен утверждать, что разница в способностях между видами A и B объясняется тем, что стимулы, использованные в эксперименте с видом A, отличались по размеру от тех, которые использовались в эксперименте с видом B.С другой стороны, устройство, которое не позволяет животному должным образом обрабатывать стимул, действительно может быть обоснованным обращением к контекстной переменной. Действительно, Макфейл (1985) пришел к выводу, что нельзя упускать из виду важность контекстных переменных, и мы полностью разделяем эту точку зрения. Как мы покажем в текущем обзоре, контекстные переменные часто играют роль в том, может ли животное проявлять определенную способность.
Сначала мы сосредоточимся на наборе когнитивных способностей, называемых отношениями эквивалентности (рефлексивность, симметрия и транзитивность).Хотя идея отношений эквивалентности может и не вызывать представления о когнитивном мастерстве, отношения эквивалентности лежат в основе ряда сложных форм поведения. Согласно Сидману (2018, стр. 33), например, отношения эквивалентности играют центральную роль, «в том, чтобы сделать язык таким мощным фактором в нашем повседневном социальном общении друг с другом».
Рефлексивность
Первое отношение эквивалентности, которое мы исследуем, — это рефлексивность, более известная в литературе по познанию животных как концепция «одинаковое-различное» или «соответствие».Задача, наиболее часто используемая для изучения того, могут ли животные сформировать концепцию сопоставления, — это задача одновременного сопоставления с образцом (SMS). Хотя вариантов много, основная процедура очень проста. Животному показывают образец стимула, например, геометрическую форму круга или вертикальной линии. После ответа на образец стимула два стимула сравнения появляются по обе стороны от образца стимула, один такой же, как образец, а другой другой. Животное должно реагировать на стимул сравнения, который совпадает с стимулом образца.В этом примере от испытания к испытанию в образце чередуются стимулы в виде круга и вертикальной линии.
Животное может решить SMS-задачу одним из трех основных способов (Скиннер, 1950; Фартинг и Опуда, 1974; Картер и Вернер, 1978). Один из способов — изучить каждую из возможных конфигураций стимулов выборки и сравнения. С двумя стимулами (A и B) и стимулами, расположенными так, что образец стимула появляется в центре, а стимулы сравнения появляются по обе стороны от образца стимула, существует четыре возможных конфигурации сравнения образцов: AAB, BAA, BBA, и ABB.В соответствии с представлением конфигурации животное узнает, что конфигурации AAB и BBA означают клевание левого стимула для получения вознаграждения, а конфигурации ABB и BAA означают клевание правого стимула для получения вознаграждения. Второй способ решить задачу SMS — это изучить серию ассоциаций стимул-ответ , таких как «если круг был образцом, то нажмите на стимул сравнения круга» и «если вертикальная линия была образцом, то нажмите на стимул сравнения вертикальной линии. .Наконец, третий способ решить задачу SMS — это изучить обобщенную концепцию соответствия , такую как «клевать стимул сравнения, который соответствует образцу стимула». Решение задачи путем реализации концепции обобщенного соответствия является «необходимым следствием рефлексивности, которая, следовательно, передает понятие тождества» (Сидман и др., 1982, стр. 24).
Чтобы выяснить, какой из трех возможных способов решения задачи SMS животным, проводится тест передачи, в котором испытуемым предъявляются новые стимулы, такие как красный и зеленый.Существует множество вопросов, касающихся условий, которые должны преобладать во время теста передачи, чтобы вывести решение с помощью концепции сопоставления. Во-первых, «новые» стимулы должны быть действительно новыми в том смысле, что нельзя использовать понятие обобщения стимулов для объяснения хороших характеристик передачи. Другими словами, если мы дрессируем животное с кругом и вертикальной линией в качестве стимулов и тестируем их с помощью овальной и наклонной линии, хорошие результаты теста передачи, скорее всего, связаны с обобщением стимула, чем с применением концепции сопоставления.Чтобы избежать ловушки генерализации стимула, стимулы в тесте передачи должны быть полностью отличными (т.е. ортогональными) обучающим стимулам. В нашем примере стимулы передачи красного и зеленого цвета ортогональны обучающим стимулам круга и вертикальной линии.
Другой важный вопрос — насколько хороша должна быть производительность передачи с новыми стимулами, чтобы сделать вывод о решении с помощью концепции сопоставления? Основная идея состоит в том, что если животные выучили исходную задачу, приняв концепцию соответствия, они должны быстро перейти на новые стимулы, потому что концепция соответствия имеет тенденцию быть независимой от стимулов.С другой стороны, если животные выучили исходную задачу с использованием правила конфигурации или правила ассоциации стимул-ответ , то выполнение новых стимулов должно быть плохим, потому что оба эти процесса зависят от исходных стимулов, и в самом деле, чтобы научиться заданию с новыми стимулами передачи, животному может потребоваться столько же попыток, сколько и разучить задание с помощью обучающих стимулов. Естественно, что какая-либо из этих экстремальных ситуаций встречается редко, и часто мы остаемся с мерами экономии, исходя из которых нужно принимать правильные решения относительно того, какой процесс использовало животное.Например, если животному потребовалось 500 попыток, чтобы выучить исходное задание с круговыми и вертикальными стимулами, и им потребовалось 50 попыток, чтобы изучить задание с красными и зелеными стимулами, это достаточно хорошее выполнение, из которого можно сделать вывод, что исходное задание были изучены с использованием подходящей концепции? Скорее всего, да. Но как насчет 100 испытаний?
Существует множество свидетельств того, что животные учатся решать задачи SMS, применяя концепцию сопоставления, по широкому кругу видов.Шимпанзе, как взрослые (Nissen et al., 1948; Robinson, 1955), так и младенцы (Oden et al., 1988), легко переходят на новые стимулы до такой степени, что можно почти говорить о почти идеальном уровне производительности при первом же подходе. несколько испытаний. Например, как группа, детеныши шимпанзе в Oden et al. (1988) в исследовании потребовалось 816 испытаний, чтобы изучить задачу сопоставления с обучающими стимулами до уровня примерно 85% правильных, и продолжали набирать баллы на этом уровне в первых 24 испытаниях с множеством различных новых стимулов.Хотя обезьяны не совсем соответствуют уровню компетентности шимпанзе, они также способны демонстрировать высокий уровень передачи новых стимулов (Mello, 1971; Milner, 1973; D’Amato et al., 1985a).
За исключением приматов, не относящихся к человеку, исследования в основном сосредоточены на способностях голубей. Ранние исследования либо не смогли найти доказательств соответствия концепции (Камминг и Берриман, 1961; Фартинг и Опуда, 1974; Холмс, 1979), но предоставили в лучшем случае слабые доказательства соответствия концепции (Wilson et al., 1985a, b), либо свидетельства соответствия концепции были открыты для альтернативных объяснений (Zentall and Hogan, 1974, 1978; Urcuioli and Nevin, 1975; Edwards et al., 1983). Одним из таких альтернативных объяснений было обычное кодирование стимулов. Например, Зенталл и Хоган (1974) тренировали голубей с помощью красных и зеленых стимулов, а затем тестировали с желтыми и синими стимулами, и птицы показали достаточно хорошие уровни перехода на «новые» стимулы. К сожалению, голуби склонны кодировать желтый и красный как одинаковые, а синий и зеленый как одинаковые (Wright and Cummings, 1971), поэтому наблюдаемая передача была не более чем примером обобщения стимулов, то есть нарушением принципа ортогональности. .Даже дальнейшее исследование (Zentall and Hogan, 1976), в котором голуби тренировались с кругом и поперечными геометрическими формами, а затем переходили на (явно новые) красные и зеленые стимулы, показало высокий уровень передачи, но не смогло распознать, что голуби изучают задание SMS. с красными и зелеными стимулами очень быстро (Zentall and Hogan, 1974), что ставит под сомнение то, что быстрый переход на красный и зеленый был вызван применением концепции соответствия.
Ранние исследования концепций сопоставления голубей имели тенденцию поддерживать точку зрения о том, что вместо изучения концепции сопоставления поведение голубей может быть лучше всего описано как обучение серии ассоциаций стимула-реакции или конфигураций.Однако доказательства того, что голуби формируют концепцию соответствия, сделали большой шаг вперед, когда Райт (1997) показал, что количество ответов, испускаемых на образец стимула, является решающим фактором, определяющим, сформируют ли голуби подходящую концепцию. Различные группы птиц были обучены испускать FR0, FR1, FR10 или FR20 на образец стимула, а затем тестировались с новыми стимулами в одинаковых условиях реакции. Райт (1997) обнаружил, что птицы, обученные с помощью FR0 или FR1, не смогли перейти на новые стимулы, тогда как птицы, обученные с помощью FR10 или FR20, показали уровни производительности с новыми стимулами, подобными (или эквивалентными в случае состояния FR20) их терминальное выступление с обучающими стимулами.Количество ответов, испускаемых на образец стимула, было контекстной переменной, которая не учитывалась во многих ранних исследованиях голубей, когда на образец стимула требовалось мало ответов.
Согласно Райту (1997), конфигурационное обучение является доминирующей и предпочтительной стратегией обучения для голубей, и для того, чтобы продемонстрировать свидетельство соответствия концепции, нужно сначала сломать предрасположенность к обработке выборки и стимулов сравнения как конфигурации. Фактически, чем больше требование FR, тем больше вероятность того, что животное отделяется от конфигурирующего обучения, и тем более вероятно, что тогда оно примет подходящую концепцию.Возьмем случай условия FR0. Образцы стимулов и стимулы сравнения предъявляются одновременно, поэтому маловероятно, что птицы даже осознают наличие «образца» стимула, который необходимо сопоставить с одним из стимулов «сравнения». И зачем им это? Фактически, единственное решение при условии FR0 — рассматривать все отображение стимулов «выборки» и «сравнения» как единое целое, то есть конфигурацию, и соответствующим образом направлять свои ответы. С другой стороны, в состоянии FR20 образец появляется, и только после 20 ответов появляются стимулы сравнения.Структура этой задачи побуждает животных воспринимать образец как нечто, что они должны сопоставить для сравнения, и в результате голуби более склонны принимать концепцию сопоставления и переходить на новые стимулы.
Последующее исследование Colombo et al. (2003) обнаружили еще одну контекстную переменную, которую необходимо скорректировать, прежде чем голуби отобразят подходящую концепцию. Эти авторы были удивлены, когда их голуби FR20 не смогли перейти на новые стимулы. Однако они отметили, что еще одно различие между исследованием Райта (1997) и их исследованием заключалось в том, что Райт (1997) первоначально тренировал своих птиц с помощью трех стимулов, тогда как Коломбо и др.(2003) тренировали их всего с двумя. Хотя обучение с двумя или тремя стимулами может показаться неважной контекстной переменной, два обучающих стимула дают четыре возможных конфигурации сравнения выборок, тогда как три обучающих стимула дают 12 возможных конфигураций сравнения выборок. Действительно, когда Коломбо и др. (2003) обучили другую группу птиц трем стимулам и требованию FR20, они перешли на новые стимулы на очень высоком уровне. Таким образом, обучающих стимулов также являются контекстной переменной.Они рассудили, что, хотя можно было бы изучить правые / левые ответы, связанные с четырьмя конфигурациями, изучение правых / левых ответов, связанных с 12 конфигурациями, может представлять трудности для животных и поощрять использование концепции соответствия для решения задачи.
Таким образом, если правильно спланировать эксперимент, можно показать уровни переноса у голубей, фактически идентичные уровням переноса у обезьян (Colombo et al., 2003). Верно, что в случае с голубем нужно наложить FR20 на образец стимула и обучить их с тремя стимулами, по сравнению с обезьянами, которые демонстрируют переход с FR1 на образец стимула и обучение всего с двумя стимулами.Однако, как только эти контекстные переменные учтены, поведение голубей становится неотличимым от обезьян. Это верно не только для условий, которые приводят к успешной передаче, но также и для условий, которые приводят к неудачной передаче (см. Рисунок 1). И D’Amato et al. (1985a) и Colombo et al. (2003) в исследованиях использовался тот же формат обучения и тестирования, в котором животных обучали с помощью ряда стимулов, а затем тестировали в течение четырех сеансов с новыми стимулами, а также с обучающими стимулами.Из рисунка 1 видно, что, когда контекстные переменные FR и количество обучающих стимулов регулируются, характеристики переноса птиц неотличимы от таковых у обезьян, как с точки зрения успешного перехода к новому цвету, так и по форме стимула ( левая панель), а также неудачный переход на два стимула новой формы (правая панель). Подробнее о разнице между переходом на стимулы цвета / формы и формы / формы позже.
Рисунок 1 . Передаточное представление обезьян и голубей.Данные по обезьянам основаны на D’Amato et al. (1985a), а данные о голубях основаны на Colombo et al. (2003). Животных тестировали в течение четырех 48-пробных сессий, причем половина испытаний была посвящена обучающим (старым) стимулам, а половина — новым (новым) стимулам. Левая панель показывает эффективность перехода к стимулу нового цвета и формы (для обучения обезьян тренировали с помощью стимулов двух форм, а голубей тренировали с помощью стимулов трех форм). Правая панель показывает эффективность переноса на два стимула новой формы (для обучения обезьян тренировали с помощью стимула цвета и формы, тогда как голубей обучали с помощью стимула двух форм и стимула цвета).Когда контекстные переменные установлены надлежащим образом для голубей (обучение с тремя стимулами и FR20 на образце), и обезьяны, и голуби легко переходят на новый цвет и формируют стимул, но не на два стимула новой формы.
Еще один последний момент в литературе по концептуальной совместимости заслуживает некоторого внимания. Премак (1983) утверждал, что животных можно различать на основе применяемой процедуры сопоставления. Согласно Premack (1983), процедуры, обсуждаемые во всех вышеупомянутых исследованиях, представляют собой то, что он называет «последовательными» задачами сопоставления, когда одинаковые или разные реакции направлены на сами физические стимулы (например,g., нажмите красный стимул сравнения, если образец был красным). Премак (1983) считает, что способность решать такие «последовательные» задачи сопоставления широко распространена среди животных. С другой стороны, задачу «одновременного» сопоставления может решить только не любой шимпанзе, а только обученный языку шимпанзе. В «одновременной» процедуре различение, на основании которого должны выноситься суждения об одном и том же и различном, отделено от фактических стимулов, которые оцениваются как одинаковые и разные. Например, если животному были предъявлены стимулы A и B, ему пришлось бы выбрать сигнал, означающий «разные», например красный прямоугольник, или, если животному были предъявлены стимулы A и A, ему пришлось бы выбрать сигнал, который означает «то же самое». , »Желтый прямоугольник (Premack et al., 1978). Кажется мало сомнений в том, что шимпанзе могут решать такие «одновременные» задачи (Premack et al., 1978), и, несмотря на заявление Премака (1983) о предварительных условиях языкового обучения, то же самое могут сделать и обезьяны без обучения языку (Sands and Wright, 1980; Д’Амато и Коломбо, 1989).
Могут ли голуби решать задачи одновременного сопоставления, как это часто бывает с голубями, требовалось больше времени, чтобы показать. Ранние положительные отчеты были омрачены альтернативными интерпретациями, такими как животные, потенциально обучающиеся фиксированному порядку левых / правых ответов, связанных с «одинаковыми» и «разными» результатами (Сантьяго и Райт, 1984), или неспособность полностью сбалансировать дизайн, тем самым позволяя птицам решать задачу, используя ассоциации для конкретных предметов (Эдвардс и др., 1983). Намного лучшая производительность передачи была получена при одновременных задачах сопоставления, когда дискриминанда состояла из массивов нескольких одинаковых и нескольких разных стимулов (Сантьяго и Райт, 1984; Вассерман и др., 1995; Кук и др., 1997). Если на самом деле птицы обрабатывают определенные элементы в массивах, то эти исследования предоставят доказательства того, что голуби могут одновременно решать задачи сопоставления. Однако критика этих исследований состоит в том, что новые «одинаковые» и «разные» массивы на самом деле не новы.Если вместо того, чтобы смотреть на отдельные элементы, составляющие массив, животные обрабатывают глобальный признак, возможно, меру «энтропии» массива стимулов, тогда «новые» массивы действительно не новы (Young et al. , 1997). Однако совсем недавно Блейсделл и Кук (2005) показали, что голуби могут выполнять задачу одновременного сопоставления, когда одновременно предъявляются только два стимула, и они переходят на новые стимулы на уровне, который предполагает свидетельство концепции сопоставления.
Когда Макфейл (1985) заявил об отсутствии качественных или количественных различий между видами, он имел в виду только виды позвоночных. Чтобы убедить нас в отсутствии различий между позвоночными в способности формировать подходящую концепцию, стоит завершить этот раздел исследованием концепции сопоставления с использованием беспозвоночных. Giurfa et al. (2001) показали, что пчелы также решают задачу SMS, используя концепцию сопоставления. Они использовали Y-лабиринт, в котором пчелы сталкивались с образцом стимула на ножке Y-лабиринта и со сравнительными стимулами на рукавах Y-лабиринта.Пчелы легко усвоили задание и отлично перенеслись на новые раздражители. Производительность пчел была настолько исключительной, что они не только передавали новые визуальные стимулы, но также передавали концепцию соответствия между модальностями, способность, которая никогда не проявлялась даже у нечеловеческих приматов (см. D’Amato et al., 1985а). Таким образом, когда контекстные переменные корректируются для каждого вида, ряд животных демонстрируют переход на новые стимулы на уровне, который предполагает использование соответствующей концепции.На самом деле, необходимо ли формулировать производительность в терминах когнитивной конструкции концепции соответствия , в отличие от работы ассоциативных процессов, — это вопрос, к которому мы вернемся в конце этого обзора.
Симметрия
Второе отношение эквивалентности, которое мы исследуем, известно как симметрия. Когда вы выучите название объекта, скажите «дверь», слово «дверь» вызывает в памяти образ двери. Точно так же изображение двери напоминает слово «дверь».Это пример симметрии , двунаправленной ассоциации между двумя стимулами. Симметрия в контексте литературы о животных обычно обучается с использованием версии задачи сопоставления с образцом, называемой символическим или условным сопоставлением с образцом, в которой разные образцы стимулов отображаются на разные стимулы сравнения. Таким образом, цель задачи состоит не в том, чтобы сопоставить с точки зрения сходства стимул сравнения с образцом стимула, а в том, чтобы выбрать стимул сравнения, который связан с образцом стимула.Например, если A1 и A2 являются выборочными стимулами, а B1 и B2 — стимулами для сравнения, тогда, когда A1 появляется в качестве образца, правильным выбором будет B1, тогда как когда A2 появляется в качестве образца, правильным выбором будет B2. Для проверки симметрии B1 и B2 теперь становятся выборочными стимулами, а A1 и A2 — стимулами сравнения. Если усвоенные отношения A1 → B1 и A2 → B2 симметричны, то при предъявлении B1 или B2 в качестве эталонных стимулов испытуемый должен выбрать A1 и A2 соответственно. Хотя животные легко обучаются символическим задачам сопоставления с образцами, демонстрация симметрии у ряда видов оказалась сложной задачей.
Здесь стоит упомянуть, что термин «симметрия» обычно подразумевает, что обратная ассоциация изучается в той же степени, что и прямая ассоциация. Судя по этому суровому определению, может показаться, что существует мало или совсем нет доказательств такой симметрии у нечеловеческих животных. Как и в большинстве случаев, в литературе, посвященной нечеловеческим животным, мы принимаем значительную обратную связь (хотя и менее выраженную, чем прямую) как доказательство симметрии. Имея это в виду, Tomonaga et al. (1991) обучили трех шимпанзе сопоставить один из двух образцов цвета с одной из двух форм сравнения с критерием не менее 80%, затем перетренировали животных для сотен испытаний, а затем проверили симметрию в 12 испытаниях.Имейте в виду, что проверка проявления способности всего за 12 попыток — сложная задача, поскольку животные часто страдают от любого изменения условий тестирования. Тем не менее, один из трех дрессированных шимпанзе показал превосходные результаты в тесте на симметрию, что свидетельствует о том, что шимпанзе способны формировать симметричные отношения. Однако доказательства симметрии у шимпанзе отнюдь не всегда положительны. Ямамото и Асано (1995), например, обнаружили, что у их одного шимпанзе не было доказательств симметрии после тренировки с одним набором стимулов, но после специальной тренировки и тестирования с шестью наборами стимулов, процедура, называемая образцовой тренировкой, все же появилась.
Демонстрация симметрии у обезьян также была встречена с большим трудом. Сидман и др. (1982) не смогли показать никаких доказательств симметрии у обезьян, обученных с помощью геометрических (вертикальная и горизонтальная линия) образцов и стимулов для сравнения цветов. Макинтайр и др. (1987) якобы показать доказательства симметрии у макак; однако их выводы были встречены серьезной критикой на том основании, что проверенные отношения уже были обучены (см. Hayes, 1989). Удивительно, но исследование D’Amato et al.(1985b), который обычно называют отрицательным открытием (Hayes, 1989; Sidman, 1994; Lionello-DeNolf and Urcuioli, 2002; Frank and Wasserman, 2005), дает некоторые благоприятные доказательства симметрии у обезьян. Д’Амато и др. (1985b) утверждали, что использование стимулов сравнения вертикальных и горизонтальных линий в Sidman et al. (1982) исследование могло быть контекстной переменной, которая поставила обезьян в невыгодное положение. Используя гораздо более различимые стимулы в качестве выборки и сравнения, а также оценивая эффективность первых 12 испытаний, D’Amato et al.(1985b) показали наличие значительных обратных ассоциаций у двух из шести протестированных обезьян.
Многочисленные исследования изучали степень симметрии голубей, и было трудно получить положительные результаты. Ранние исследования либо не смогли найти никаких доказательств даже обратных ассоциаций (Lipkens et al., 1988), были подвергнуты критике за альтернативные интерпретации, когда они это сделали (Vaughan, 1988; Hayes, 1989), либо во многом аналогично шимпанзе и обезьянам, найденным в лучшем случае только слабые доказательства обратных ассоциаций (Hogan and Zentall, 1977; Richards, 1988).Интересно, что в исследовании Hogan and Zentall (1977) некоторые положительные доказательства симметрии были замечены в начале теста на симметрию, но затем рассеялись, что также наблюдалось D’Amato et al. (1985b) с обезьянами. Учитывая контекст этой статьи, возможно, уместно включить одну возможность, поднятую Хоганом и Зенталлом (1977, стр. 14), относительно того, почему голуби плохо справляются с задачами симметрии: «также возможно, что развитие обратных ассоциаций зависит от видоспецифическая функциональная ценность таких ассоциаций (т.е., у людей может возникнуть необходимость в формировании обратных ассоциаций, а у голубей — нет).
Лионелло-ДеНольф и Уркуиоли (2002) также не смогли найти доказательств симметрии у голубей, но их исследование заслуживает упоминания, потому что оно представляет собой одну из самых ранних попыток обратиться к возможным контекстным переменным, которые могут мешать голубям (и, возможно, другим животным) демонстрировать симметрия. Основываясь на представлении McIlvane et al. (2000) о топографии стимулов-ответов о том, что голуби могут обрабатывать аспекты стимула, которые мешают интересующим аспектам тестов на симметрию, Лайонелло-ДеНольф и Уркуиоли (2002) пришли к выводу, что во время Проверка симметрии не только выполняет обмен ролями выборки и сравнения, но и меняет позиции.Таким образом, голуби, по-видимому, кодируют не только особенности стимула, но и положение стимулов как часть топографии реакции на стимул . Возьмем ситуацию в задачах сопоставления, когда образец стимула обычно появляется в центре, а стимулы сравнения появляются по обе стороны от положения образца. Для проверки симметрии стимулы сравнения теперь появляются в центральной позиции. Для человека может быть неуместно то, что стимул сравнения теперь появляется в положении, в котором он никогда не появлялся раньше, но для нечеловека позиция может быть частью топографии реакции на стимул , и, следовательно, нечеловеческие животные могут нарушить симметрию. тест, потому что неясно, как они должны себя вести, когда стимулы появляются в позициях, в которых они никогда не появлялись раньше.Таким образом, Лионелло-ДеНольф и Уркуиоли (2002) обучили своих животных так, чтобы стимулы выборки и сравнения могли появляться в любом из ряда положений, таким образом эффективно тренируя «положение» как компонент топографии реакции на стимул . Несмотря на это обучение, голуби по-прежнему не смогли показать никаких доказательств симметрии, открытие, которое, несмотря на то, что доказательства симметрии есть у нечеловеческих приматов, еще больше отдаляет голубей от нечеловеческих в их способности формировать симметричные отношения.
Однако Франк и Вассерман (2005) отметили, что помимо стимулов, связанных с их пространственным расположением, они также связаны с их временным расположением. Другими словами, если отношение A1 → B1 обучается, а затем проверяется отношение B1 → A1, элемент B никогда не появляется первым. Как и в случае с положением, упомянутым ранее, если элемент B теперь появляется первым, мы, люди, можем быстро предположить, что, поскольку он появляется первым, он должен выполнять роль образца стимула, но, опять же, нет причин, по которым другие животные должны производить это предположение.Чтобы учесть потенциально контролирующее влияние контекстной переменной временного местоположения, Фрэнк и Вассерман (2005) использовали последовательные задачи на сопоставление «годен / не годен», где стимулы выборки и сравнения появляются последовательно в одной и той же позиции, а субъект должен ответьте на второй стимул, если он сочетается с первым (например, A1 → B1), и отрицательный ответ (т.е. воздержитесь от ответа) на второй стимул, если он не сочетается с первым (например, A1 → B2). Чтобы контролировать потенциально разрушительные эффекты контекстной переменной временного порядка и тот факт, что, например, стимул B никогда не появлялся первым, голубей обучали не только символическим отношениям (A1 → B1, A2 → B2), но также и тем, что отношения идентичности (A → A и B → B), таким образом обучая животных тому, что оба стимула A и B могут возникать в любой временной позиции.Принимая во внимание эти контекстные переменные, голуби демонстрировали устойчивую симметрию. Франк и Вассерман (2005, стр. 157) пришли к выводу, что «симметрия может быть получена с нечеловеческими животными при надлежащих условиях обучения и тестирования». Интересно, что один успешный шимпанзе в книге Tomonaga et al. (1991) исследование также проходило как с символическими отношениями, так и с отношениями идентичности.
Таким образом, элегантное исследование Фрэнка и Вассермана (2005) показывает, что если принять во внимание контекстные переменные, голуби могут демонстрировать симметрию, причем на уровне, не слишком отличном от шимпанзе.Кроме того, мало, если вообще есть доказательства, которые могли бы различать поведение нечеловеческих приматов и птиц в отношении формирования симметрии. Как и в литературе, посвященной концептуальному соответствию, исследователи сейчас исследуют, можно ли продемонстрировать симметрию беспозвоночными. Учитывая недавнюю попытку Moreno et al. (2012) с пчелами, кажется, что появление симметрии у беспозвоночных — лишь вопрос времени.
Транзитивность
Третье отношение эквивалентности, которое мы исследуем, известно как транзитивность.Нет необходимости апеллировать к понятию контекстных переменных, потому что было показано, что большинство видов способны решать задачи транзитивности. Скорее, мы включаем краткое упоминание этой темы, чтобы завершить обсуждение отношений эквивалентности, и, что более важно, чтобы выделить другую проблему, которую мы хотим кратко затронуть в этом обзоре, а именно желание интерпретировать поведение нечеловеческих животных в откровенно когнитивно богатых терминах. .
Транзитивность — это операция, при которой, учитывая информацию о том, что A умнее B, а B умнее C, можно сделать логический вывод, что A умнее C, даже несмотря на то, что никакой прямой информации о взаимосвязи между A и C никогда не давалось. .Согласно Пиаже (1928), способность решать такую задачу трехчленного транзитивного вывода не развивается примерно до 7 лет, и этот вывод был оспорен Брайантом и Трабассо (1971), которые продемонстрировали надежные способности к транзитивному выводу в четырех случаях. -, 5- и 6-летние. Хотя основной целью этого обзора является сравнение нечеловеческих животных, стоит упомянуть процедуру, использованную Брайантом и Трабассо (1971), потому что очень похожие процедуры обучения использовались для изучения транзитивности у нечеловеческих животных.В своем исследовании детей учили различать цветные стержни разной длины. Стержни были представлены парами, и тренировка состояла из повторных воздействий на четыре тренировочные пары, A + B−, B + C−, C + D− и D + E−, с буквами, обозначающими разные длины стержней (например, , A был самым длинным, а E — самым коротким), а знаки «+» и «-» указывают на правильный и неправильный стимул, соответственно, для выбора. Например, при предъявлении парного компакт-диска и подсказке с вопросом «, какая штанга длиннее? ”, испытуемый должен выбрать пункт C.
Конечно, при изучении транзитивного ряда из пяти элементов A всегда правильно, E всегда неверно, а B, C и D правильны и неверны в зависимости от пары, в которой они появляются. В серии из пяти пунктов есть 10 возможных пар для тестирования предмета (AB, AC, AD, AE, BC, BD, BE, CD, CE и DE). Из них мы ожидаем, что испытуемый будет хорошо работать с любой парой, содержащей элемент A (AB, AC, AD и AE), потому что при обучении элемент A всегда был правильным. Мы также ожидаем, что испытуемые будут хорошо работать с любой парой, содержащей элемент E (AE, BE, CE и DE), потому что при обучении элемент E всегда был неправильным, и, следовательно, его всегда следует избегать в пользу другого стимула.Наконец, мы явно ожидаем, что они будут хорошо работать с парой, которая была одной из обучающих пар (AB, BC, CD и DE), оставив в качестве критического теста для пары транзитивность BD. Брайант и Трабассо (1971) обнаружили, что 4-, 5- и 6-летние дети хорошо справляются с парным BD.
Исследования с участием нечеловеческих животных имеют тенденцию следовать той же общей процедуре, принятой Брайантом и Трабассо (1971): сначала дрессируют животных на четырех парах предпосылок AB, BC, CD и DE, а затем тестируют их на критической паре BD.Используя эту процедуру, шимпанзе (Gillan, 1981), обезьяны (McGonigle and Chalmers, 1977), крысы (Davis, 1992) и голуби (von Fersen et al., 1991; Paz-y-Miño C et al., 2004) имеют все они показали высокие результаты в паре критических тестов BD и действительно достигли уровня производительности, не слишком отличающегося от того, о котором сообщили Брайант и Трабассо (1971) для детей младшего возраста. Конечно, у голубей не всегда проявлялась транзитивность (D’Amato et al., 1985b), но также были случаи, когда у приматов не отображалась транзитивность (Sidman et al., 1982). Несмотря на случайные неудачи, нет необходимости обращаться к контекстным переменным, потому что в целом голуби решали задачи транзитивности так же, как и другие животные.
Ключевой особенностью многих из этих исследований является степень, в которой высокий уровень производительности в тестовой паре BD отражает когнитивную / логическую операцию или поведенческую / ассоциативную операцию. В когнитивно-логическом лагере придерживаются мнения, что при обучении реагированию на каждую из пяти пар предпосылок (например,, A + B−, B + C−, C + D− и D + E−) животные образуют иерархическое линейное мысленное представление о том, как пять стимулов связаны друг с другом (например, A> B> C> D> E), и используйте это представление, чтобы направлять их относительно того, как реагировать на критическую тестовую пару BD. В поведенческом / ассоциативном лагере не формируется линейное мысленное представление пяти пунктов. Скорее решение критической пары BD основано на проведении ассоциативных вычислений на основе значений вознаграждения, присвоенных каждому из элементов (теория передачи ценности), или на использовании ранее изученных пар предпосылок для решения проблемы BD (модель двоичной выборки).Согласно теории передачи ценностей (von Fersen et al., 1990, 1991), каждому из пяти стимулов присваивается разная сила в зависимости от того, в каких парах они появились во время тренировки и были ли они связаны со стимулом, который всегда вознаграждается. A или стимул E, который никогда не вознаграждается. В результате таких ассоциаций элемент B получает более высокий рейтинг, чем элемент D, и поэтому животное выберет B, когда ему будет представлена пара BD. Действительно, полученные рейтинги можно использовать для очень точного предсказания, какой стимул выберет животное, когда любые два стимула будут парными.
Модель двоичной выборки (McGonigle and Chalmers, 1977) также является простым, но эффективным некогнитивным объяснением того, почему животное выбирает элемент B во время критического теста BD. Согласно модели (см. Рисунок 2), увидев пару BD, животное пытается решить задачу, как если бы это была пара BC, CD или BD. Сеанс тестирования обычно состоит из множества представлений пары BD, и, согласно модели, существует 1 из 3 шансов, что любая из трех пар будет выбрана в любом данном испытании.Учитывая, что каждая пара выбирается в 33% случаев, мы можем думать о каждой паре как о 0,33 единицы, которые вносят вклад в решение проблемы BD. Если животное пытается решить пару BD, как если бы это была пара BC, оно выберет B, потому что B + C− — одна из обучающих пар, где животное учат выбирать B. Таким образом, элемент B набирает 0,33 единицы. Если животное пытается решить пару BD, как если бы это была пара CD, оно выберет C, потому что C + D− является одной из обучающих пар, где животное учат выбирать C.Но имейте в виду, что нет пункта C для выбора, потому что, помните, животному представлена пара BD, а не CD. Однако, если животное пыталось решить пару BD, как если бы это была пара CD, животное также обучается этой паре, чтобы избегать элемента D, и поэтому животное избегает элемента D в паре BD и выбирает элемент B. 0,33 единицы, в результате чего текущий показатель составляет 0,66 единицы. Наконец, животное может попытаться решить пару BD, как если бы это была пара BD. К сожалению, пара BD не является обучающей парой, поэтому не было установлено никаких ассоциаций между элементами B и D, и животное случайным образом выберет B в половине случаев, D в половине случаев и 0.33 доступных юнита делятся между двумя предметами. Окончательный результат таков: элемент B получает 0,83 единицы, а элемент D — 0,16 единицы, что при выражении в процентах правильных ответов очень близко к показателям животных с тестовой парой BD в широком диапазоне исследований.
Рисунок 2 . Модель двоичной выборки. Согласно МакГониглу и Чалмерсу (1977), животное пытается решить пару BD, как если бы это была пара BC, CD или BD. B + C− — это обучающая пара, поэтому она выберет элемент B.C + D− также является обучающей парой, в которой животное учат выбирать C. К сожалению, поскольку животному была представлена пара BD, элемент C не отображается, и, следовательно, нет элемента C для выбора. Но если животное пыталось решить пару BD, как если бы это был пункт C + D−, оно также научилось избегать D, что и делает животное, и снова выбирает пункт B. Наконец, если животное пытается решить пару BD, как если бы это была BD, он не получил обучения с этими двумя стимулами, представленными вместе, и случайным образом выбирает между ними.В результате животное будет выбирать B в 83% случаев и D в 17% случаев, что очень близко к уровням производительности, достигнутым многими животными с помощью теста BD.
Теория передачи значений и модель двоичной выборки, а также другие некогнитивные объяснения транзитивности (см. Delius and Siemann, 1998) очень хорошо объясняют высокие уровни производительности тестовых пар BD без необходимости обращения к когнитивным учетным записям, таким как иерархические ментальные представления. Конечно, у этих более простых объяснений транзитивного вывода были проблемы (Steirn et al., 1995; Лазарева и Вассерман, 2012), но трудно не заметить силу ассоциативной силы через историю подкрепления (Siemann et al., 1996). В этом обзоре трудно отдать должное литературе по сложным транзитивным выводам, но независимо от того, верит ли кто-то в когнитивные / логические или поведенческие / ассоциативные объяснения, одно можно сказать наверняка: точка зрения, что обезьяны выполняют такие задачи по-разному, не поддерживается. голубям. Недавняя демонстрация этой способности у беспозвоночных означает, что этот вопрос теперь можно распространить за пределы позвоночных (Tibbetts et al., 2019). Возможно, что еще более важно, тот факт, что вид, имеющий всего 0,001% нейронов в человеческом мозгу (Azevedo et al., 2009; Menzel, 2012), может выполнить задачу, должен поставить под сомнение когнитивно-богатые термины, с помощью которых исследователи описывают переходные процессы. вывод.
Поведение последовательного заказа
Концептуально с исследованиями транзитивности связаны исследования, изучающие способности животных к последовательному порядку. Задача последовательного порядка, также известная как процедура одновременного связывания, предоставила большой объем информации о структуре представлений, которые, как считается, лежат в основе транзитивных суждений.Задача проста и, как и процедура транзитивности, часто использует пять стимулов. Однако вместо того, чтобы представлять пять стимулов в виде четырех пар обучающих предпосылок, в задании последовательного порядка животных обучают реагировать на пять одновременно предъявляемых стимулов в определенном порядке, а именно: A → B → C → D → E. И обезьяны, и голуби могут научиться выполнять задачи последовательного заказа из пяти предметов на одном и том же высоком уровне (D’Amato and Colombo, 1988; Terrace 1993; Scarf and Colombo, 2010). Чтобы определить, чему научились животные, как и в тесте на транзитивность, испытуемым предлагается попарный тест, состоящий из всех 10 возможных пар стимулов, которые могут быть сгенерированы из списка из пяти пунктов (AB, AC, AD, AE, BC, BD, BE, CD, CE и DE).Правильный ответ в парном тесте требует, чтобы животные реагировали на два отображаемых элемента в соответствии с их порядком в серии из пяти элементов. Например, при представлении пары BC для получения награды животное должно сначала ответить на вопрос B, а затем на элемент C.
Попарный тест предоставил значительное понимание процессов, которые разные животные используют при обучении исходной задаче последовательного заказа из пяти элементов. Фактически, до недавнего времени производительность парных тестов, а также меры задержки, которые могут быть сгенерированы на основе правильных ответов, казались одним из лучших доказательств того, что обезьяны и голуби обрабатывают информацию о последовательном порядке принципиально разными способами (Terrace, 1993; Шарф и Коломбо, 2008).Например, с точки зрения производительности по 10 парам, обезьяны показывают очень высокие показатели во всех тестовых парах, тогда как голуби показывают высокие показатели только в парах, содержащих либо элемент A, либо элемент E. внутренние пары BC, BD и CD (см. рисунок 3). Такой результат согласуется с точкой зрения, согласно которой в процессе обучения последовательной задаче обезьяны формируют мысленное представление списка и используют это представление для управления своим поведением (D’Amato and Colombo, 1988).С другой стороны, голуби, похоже, неспособны сформировать такое представление и скорее изучают простой набор поведенческих правил, таких как «(1) Сначала отвечайте на элемент А. (2) Последним отвечайте на элемент D. (3) Отвечайте на элемент». любой другой элемент по умолчанию »(Terrace, 1993, стр. 164).
Рисунок 3 . Показатели по 10 парам во время парного теста. Обезьяны хорошо справляются со всеми парами, в то время как голуби хорошо справляются только с парами, у которых есть элемент A или элемент E, и случайно работают с внутренними парами, в которых эти элементы отсутствуют.
Еще одно свидетельство того, что обезьяны формируют мысленное представление ряда, получено из двух типов анализа данных о задержке, чтобы ответить на первый и второй элементы отображаемой пары. В случае эффекта первого элемента задержка до первого элемента пары усредняется по всем парам, которые совместно используют один и тот же первый элемент. Другими словами, задержка до элемента A усредняется по парам AB, AC, AD и AE, задержка до элемента B усредняется по парам BC, BD и BE, задержка до элемента C усредняется по парам CD и CE, и задержка до элемента D основана на единственной паре, которая имеет элемент D в качестве первого элемента, пару DE (см. рисунок 4, левая панель).Обезьяны четко демонстрируют эффект первого элемента , в котором задержка ответа на первый элемент пары тем больше, чем дальше по списку находится первый элемент. Например, время ожидания ответа на элемент C в паре CD занимает больше времени, чем время ожидания ответа на элемент B в паре BD. Такая функция задержки предполагает, что обезьяны получают доступ к списку в элементе A и продвигаются по списку линейным образом, пытаясь сопоставить элемент в памяти с отображаемым элементом. В отличие от обезьян, голуби демонстрируют плоский эффект для первого пункта .
Рисунок 4 . Левая панель : эффект первого элемента . Задержка ответа на первый элемент пары для обезьян и голубей в зависимости от того, был ли первый элемент A (усреднен по парам AB, AC, AD и AE), B (усреднен по парам BC, BD и BE ), C (усредненное по парам CD и CE) или D (только по паре DE). Обезьяны показывают линейное увеличение по первому пункту, а голуби — нет. Данные основаны на правильных испытаниях. Правая панель : Эффект пропавшего элемента .Задержка ответа на второй элемент пары в зависимости от того, был ли второй элемент отделен от первого отсутствием 0 элементов (усредненным по парам AB, BC, CD и DE), 1 отсутствующим элементом (усредненным по парам AC , BD и CE), 2 отсутствующих элемента (усредненных по парам AD и BE) или 3 отсутствующих элемента (только на основе пары AE). Обезьяны показывают линейное увеличение количества недостающих предметов, а голуби — нет. Данные основаны на правильных испытаниях.
Обезьяны также демонстрируют так называемый эффект пропавшего предмета (рис. 4, правая панель).Эффект пропущенного элемента относится к задержке ответа на второй элемент пары как функции расстояния от первого элемента до второго элемента. В парах AB, BC, CD и DE нет недостающих элементов, поскольку второй стимул пары возникает сразу после первого стимула. В парах AC, BD и CE отсутствует один элемент, в парах AD и BE отсутствуют два элемента, а в паре AE отсутствуют три элемента. Обезьяны демонстрируют очень четкий эффект пропущенного элемента в том смысле, что задержка ответа на второй элемент пары является функцией количества пропущенных элементов между первым и вторым элементом.Например, обезьяны быстрее реагируют на элемент D в паре CD, чем на элемент D в паре BD. Причина в том, что в парном CD нет недостающих элементов для доступа, тогда как в паре BD обезьяна должна получить доступ к одному недостающему элементу, элементу C. В отличие от обезьян, у голубей не отображается эффект недостающего элемента .
Производительность по 10 парам, а также наличие эффекта первого элемента и эффекта пропущенного элемента подтверждают точку зрения, что в процессе обучения задачи последовательного порядка обезьяны образуют линейное ментальное представление элементов и использовать это представление для управления их поведением, например, во время парного теста.Напротив, отсутствие этих эффектов у голубей предполагает, что они решают задачу последовательного порядка принципиально иначе, чем обезьяны. Эти взгляды хорошо согласуются с представлением о том, что успех обезьян вполне может быть связан с их способностью надлежащим образом реагировать на иерархии доминирования (Cheney et al., 1986), что не является необходимым для голубя, чья социальная структура имеет определенную структуру. гораздо менее иерархическая организация (Masure and Allee, 1934).
Правда ли, что голуби не знают порядка следования стимулов в задаче с последовательным порядком, или проявление этой способности замаскировано какой-то контекстной переменной? Напомним, что попарный тест проводится после того, как животные достигли определенного уровня мастерства в задании последовательного порядка из пяти пунктов, и состоит из представления испытуемым всех 10 пар стимулов, которые могут быть сгенерированы из пяти пунктов (AB, AC , AD, AE, BC, BD, BE, CD, CE и DE).Кроме того, каждая из 10 пар отображается несколько раз в течение сеанса (обычно четыре раза в течение сеанса из 40 проб). Мы задавались вопросом, может ли структура попарного теста и удивление от перехода от задания с пятью пунктами к парному тесту со всеми 10 парами, смешанными в течение сеанса, вызывать затруднения у голубей. Было ли резкое изменение контекста контекстной переменной, которая объясняла плохие результаты голубей в парном тесте? Мы исследовали эту возможность в двух экспериментах (Scarf and Colombo, 2010).
В одном эксперименте мы обучили четырех голубей выполнению задачи серийного заказа из четырех предметов и еще четырех голубей — задаче серийного заказа из пяти предметов. Вместо того, чтобы затем провести попарный тест из шести пар (число, которое можно получить из списка из четырех пунктов) для птиц, обученных заданию из четырех пунктов, или 10 пар для птиц, обученных заданию из пяти пунктов, мы попытались смягчить эффекты изменения контекста, представив птиц, обученных четырем пунктам, только парой BC (критическая внутренняя пара после обучения в списке из четырех пунктов) или птиц, обученных пяти пунктам, только парой BD ( критическая внутренняя пара после тренировки по списку из пяти пунктов).Пары BC или BD были представлены 40 раз за сеанс. Мы рассудили, что если голуби ничего не узнали о порядке пунктов B и C или порядке пунктов B и D, то те, кто тестировались на положительном условии пары (BC + или BD +), получали вознаграждение за нажатие B → C или B → D. должны работать не лучше, чем те, кто был протестирован на отрицательном парном условии (BC- и BD-) и получать вознаграждение за ответ на вопросы в противоположном направлении, то есть C → B или D → B. Результаты показаны на рисунке 5. Очевидно, что животные, обученные на положительных парах, освоили задачу значительно быстрее, чем те, которые были обучены на отрицательных парах, что позволяет предположить, что при правильной настройке условий голуби демонстрируют свидетельство того, что они понимают порядок внутренних элементов. по четырёх- и пятипозиционным задачам серийного заказа.
Рисунок 5 . Выполнение пары BC после тренировки по списку из четырех пунктов ( левая панель, ) и BD после тренировки по списку из пяти пунктов ( правая панель, ). По два животных были обучены условиям BC +, BC-, BD + и BD-, где знак «+» означал, что вознаграждение можно получить, отвечая на элементы в том порядке, в котором они появлялись в исходной последовательности (B → C или B → D), а знак «-» указывает на то, что награду можно получить, отвечая на элементы в порядке, противоположном тому, в котором они появлялись в исходной последовательности (C → B или D → B).Если птицы ничего не узнали о порядке внутренних элементов, то при представлении этих пар им потребуется столько же времени, чтобы выучить положительное состояние пары, как и отрицательное. Скорее, очевидно, что птицы, испытанные с положительными парами, оказались намного лучше, чем птицы, испытанные с отрицательными парами.
При тестировании только одной пары птицы смогли указать, что они действительно поняли, что элемент B предшествует элементу C, или элемент B предшествует элементу D, и, таким образом, предоставили нам свидетельство того, что они действительно поняли, по крайней мере, на некоторых элементарных уровень, организация внутренних элементов в серию.Тем не менее, птицы с положительной парой действительно испытали гораздо больше испытаний (40–80 для птиц BC +, 120 для птиц BD +) на соответствующих парах, чем те, которые обычно испытывают обезьяны на тех же парах во время обычного парного теста (обычно около 8–12 испытаний). Мы задались вопросом, смогут ли голуби когда-либо продемонстрировать высокий уровень производительности на критически важной паре, как это сделали обезьяны, после проведения ограниченного числа испытаний. Чтобы проверить это понятие, мы снова изменили процедуру попарного тестирования.Для второго эксперимента голуби были обучены выполнению четырехэлементной задачи последовательного заказа и представлены с критической парой BC в качестве теста четырех испытаний, встроенных в базовый уровень из 36 испытаний, посвященных стандарту (A → B → C → D). четырехэлементная задача серийного заказа. Тест проводился в течение четырех сеансов, что дало в общей сложности 16 испытаний до н.э., что очень похоже на то, что было на обезьянах. Результаты показаны на Рисунке 6. Все четыре птицы показали очень высокие уровни в 16 испытаниях зонда BC. Для сравнения на рисунке также показана производительность голубей в паре BC, когда она была доставлена в стандартном формате попарного теста, в котором шесть пар, которые могут быть сгенерированы после обучения с помощью списка из четырех пунктов (AB, AC, AD , BC, BD и CD) представлены смешанными в рамках сеанса (Straub and Terrace, 1981) без исходных испытаний A → B → C → D.Ясно, что голуби могут хорошо работать с критической парой после ограниченного воздействия на эту пару, но только тогда, когда контекст общего теста не сильно изменится по сравнению с ситуацией обучения.
Рисунок 6 . Производительность пары BC при поставке в виде датчиков, встроенных в базовую задачу серийного заказа из четырех изделий. Все четыре голубя показали высокие показатели после того, как подверглись испытаниям только с 16 BC зондами. Также показаны показатели BC голубей Straub и Terrace (1981), где испытания BC проводились стандартным способом попарно вместе со всеми другими парами в сеансе.
Мы показали, что, смягчая последствия резкого изменения контекста, голуби могут хорошо работать на критической внутренней тестовой паре, тем самым подтверждая мнение, что они действительно понимают порядок внутренних элементов в списке. Казалось бы, для голубей, по какой-либо причине, отображение всех пар сразу, как в стандартном парном тесте, является контекстной переменной, которая не позволяет им отображать свое понимание организации элементов четырех- и пятипозиционного теста. списки.
Эпизодическая память и теория разума
Существует множество задач, которые использовались для исследования способностей нечеловеческих животных, для которых не только нет различий в характеристиках между видами, но и для которых некоторые из наиболее убедительных доказательств определенной способности на самом деле исходят от птиц, а скорее обезьяны или шимпанзе. Такие эксперименты очень ясно говорят о Нулевая гипотеза . Речь идет об эпизодической памяти. Эпизодическая память — это воспоминания о личном опыте жизни.Тулвинг (Tulving, 1972) первоначально предполагал, что эпизодическая память состоит из памяти для того, что было событием, , где произошло событие, и , когда произошло событий в жизни человека, в просторечии называемых WWW-памятью. Позже Талвинг (1985) уточнил свое определение, включив в него концепцию автономного сознания (автоноэзиса), феноменологический опыт, который извлекается из воспоминаний, и это действительно что-то, что происходило с вами в прошлом. Если эпизодическая память определяется как требующая автономии, доступ к которой возможен только с помощью устного сообщения, то маловероятно, что какое-либо животное, не являющееся человеком, может удовлетворить критерию обладания эпизодической памятью.Однако, если мы вернемся к первоначальному определению эпизодической памяти Тулвинга (1972) как памяти для , , , , и , где , то накапливаются доказательства того, что различные животные обладают эпизодической памятью или, по крайней мере, какой-то некоторой. осторожно назвали эпизодической памятью .
Исследование Клейтона и Дикинсона (1998) до сих пор считается наиболее убедительным доказательством того, что нечеловеческие существа, в их случае сойки-кустарники, могут использовать what, , , где , и , когда информацию, чтобы управлять своим поведением.После публикации исследования Клейтона и Дикинсона (1998) было предпринято множество других попыток продемонстрировать WWW-память у ряда видов, таких как крысы (Bird et al., 2003; Babb and Crystal, 2005; Ergorul and Eichenbaum, 2007), голуби (Skov-Rackette et al., 2006), обезьяны (Hampton et al., 2005) и обезьяны (Schwartz et al., 2002, 2004, 2005; Mulcahy and Call, 2006). Во многих случаях эти эксперименты имеют альтернативные объяснения, которые не требуют атрибуции эпизодической памяти (см. Colombo and Hayne, 2010).В других случаях доказательства могут быть заманчиво близкими к данным исследования Клейтона и Дикинсона (1998) с сойками (Mulcahy and Call, 2006), но всегда, кажется, не соответствуют показателям сойок, хотя некоторые из более поздних Работа Кристалла и его коллег поднимает способности крыс наравне с сойками (обзор см. в Crystal, 2011). То же самое верно и в отношении способности планировать будущие потребности, что очень элегантно было продемонстрировано у соек (Raby et al., 2007), затем у шимпанзе (Inoue and Matsuzawa, 2007), а в последнее время и у крыс (Crystal, 2013) .Также важно помнить, что даже исследование Клейтона и Дикинсона (1998) не обходится без критиков, которые выступают против точки зрения, согласно которой сойки проявляют эпизодическую память (Suddendorf and Busby, 2003; Suddendorf and Corballis, 2007). Тем не менее, при наличии надлежащего экспериментального дизайна, это просто вопрос времени, когда все животные покажут высокий уровень мастерства в задачах WWW.
Исследования, изучающие способность животных проявлять Теорию Разума (ToM), — еще один пример того, как птицы проявляют замечательные способности.Премак и Вудрафф (1978) задали вопрос: « Есть ли у шимпанзе теория разума? ». На основании способности шимпанзе выбирать правильное изображение, изображающее решение ранее просмотренного 30-секундного видеоклипа человека, стоящего перед дилеммой, авторы пришли к выводу, что у шимпанзе действительно есть ToM. Точно так же Повинелли и его коллеги сравнили две формы атрибуции психического состояния, смену ролей и концепцию знающего и предполагающего (Povinelli et al., 1990; Povinelli, 1993).В случае эксперимента с изменением ролей шимпанзе смогли оценить не только свою роль в обеспечении еды, но и роль человека, с которым они были в паре, так что даже если переключиться на роль другого, они все равно преуспели в добывании пищи. Аналогичным образом, в эксперименте «знающий-угадывающий» шимпанзе предоставили доказательства того, что они понимали, что человек, оставшийся в комнате (знающий), знал о местонахождении спрятанной пищи, в то время как человек, вышедший из комнаты (догадывающийся), знал. нет, поэтому, когда им предоставляется выбор, они выбирают место, указанное знающим, а не угадывающим.
Хотя ни видеоклипы, ни эксперименты по смене ролей, ни эксперименты по поиску догадок с птицами не проводились, Эмери и Клейтон (2001) не исследовали влияние опыта и социального контекста на способность сойки-кустарника кэшировать пищу. Сойкам была предоставлена возможность кэшировать еду либо в присутствии сойки-наблюдателя, либо наедине. Авторы обнаружили, что сойки с гораздо большей вероятностью откроют пищу, если они ранее были кэшированы во время наблюдения, предполагая, что они понимают намерения наблюдающей сойки.Действительно, только те сойки, которые сами испытали воровство тайников, проявили такую способность, тогда как наивные сойки в наблюдаемых условиях не возвращались больше, чем в частных. Эти результаты подтверждают старую пословицу о том, что «требуется вор, чтобы узнать вора», и подчеркивают замечательные способности этих птиц в отношении определения психического состояния.
Чтобы быть уверенным, что есть критики всех этих исследований, действительно, Повинелли (1994) с тех пор признал, что шимпанзе, возможно, научились реагировать на поведенческие сигналы, а не делать выводы о состоянии знаний каждого из экспериментаторов, что гораздо проще, чем мысленный подход. государственная атрибуция.Критикуя литературу по ToM, Heyes (1998, стр. 101) оценил эмпирические доказательства того, что шимпанзе обладают ToM, и пришел к выводу, что «во всех случаях, когда нечеловеческое поведение приматов интерпретировалось как признак ToM, оно могло вместо этого иметь произошло случайно или как продукт нементалистических процессов, таких как ассоциативное обучение или умозаключение, основанное на нементальных категориях ». Аналогичным образом, результаты Эмери и Клейтона (2001) также можно отнести к простым процессам обучения и ассоциациям.Хотя мы согласны с этими более простыми интерпретациями, главное, что мы хотим сделать сейчас, это то, что нет никаких доказательств того, что конкретная способность, такая как эпизодическая память или ToM (или любая из предыдущих способностей, которые мы обсуждали), является присутствует у одного вида, а не у другого.
Возвращение к Макфейлу
Наш обзор не является исчерпывающим в том смысле, что мы не рассмотрели все задачи, по которым сравнивались виды. Например, то, как различные виды выполняют задачи по привыканию, классической обусловленности и инструментальной обусловленности, которые Макфейл назвал «простыми» задачами, было подробно рассмотрено Макфейлом (1982, 1985, 1987), и мы не собирались останавливаться на достигнутом. это опять же, главным образом потому, что, вероятно, нет особых разногласий в том, что позвоночные выполняют аналогичным образом с такими «простыми» задачами.Скорее, нашей целью было оценить нулевую гипотезу Макфейла (1985) в свете недавнего всплеска интереса к умственным способностям нечеловеческих животных и задач, которые использовались для вывода этих способностей. Эти задачи называются Макфейлом «сложными» задачами, и Макфейл понимал, что разногласия по поводу его нулевой гипотезы будут сосредоточены на этих «сложных» задачах.
Мы рассмотрели большое количество таких «сложных» задач, как рефлексивность (концепция соответствия), симметрия и поведение последовательного порядка, и показали, что различия в производительности между видами могут быть связаны с контекстной переменной, будь то FR требования к образцу стимула или количеству обучающих стимулов в случае рефлексивности, аспекты топографии стимульного ответа в случае симметрии или тестовая ситуация в случае поведения последовательного порядка.Для других задач, таких как транзитивность, эпизодическая память и ToM, производительность птиц-соперников, а иногда и выше, чем у нечеловеческих приматов. Наш обзор литературы показывает, что существует очень небольшая разница в выполнении этих «сложных» задач у разных видов позвоночных. На основании вышеприведенного обзора и несмотря на потенциальные ловушки, присущие всем подобным сравнениям, мы согласны с Макфейлом (1985, стр. 39), когда он заявил, что «в настоящее время нет феномена обучения, продемонстрированного в одном (нечеловеческом) человеке. виды позвоночных, которые не были обнаружены у всех других позвоночных, у которых его систематически искали.”
Качественные различия и количественные различия
Под качественным различием между видами подразумевается обладание одним видом механизма, который отсутствует у другого…. Количественное различие между двумя видами означало бы, что один вид использовал механизм или механизмы, общие для обоих видов, более эффективно, чем другой »(Macphail, 1985, стр. 38).
Мы не подразумеваем, что не существует примеров конкретной задачи, в которой производительность одного вида превосходила бы производительность другого.Действительно, таких случаев много. Поэтому трудно не заметить тот факт, что виды действительно различаются количественно. Легкость, с которой шимпанзе и обезьяны могут изучать задачи, слишком очевидна, и хотя скорость обучения не является лучшим показателем когнитивных способностей, она говорит о некоторой разнице в способности обработки, даже если учтены такие проблемы, как контекстные переменные. И тот простой факт, что голубю нужна определенная ситуация тестирования, в то время как обезьяна не может, еще больше говорит о количественной разнице на феноменологическом уровне и, возможно, также на уровне процесса (см. Дальнейшее обсуждение ниже).Эти количественные различия также несомненно распространяются на диапазон ситуаций переноса с более ограниченным диапазоном у голубей, чем у обезьян, и действительно более ограниченным диапазоном у обезьян, чем у шимпанзе или людей (Weinstein, 1941). И в конце концов, это способность переноситься в новые ситуации, что на самом деле является отличительной чертой того, что мы называем интеллектом, и в этом отношении способности людей превосходят способности обезьян, точно так же, как способности обезьян более чем вероятно превышают способности голуби.В самом деле, мы утверждаем, что основное различие в «интеллекте» животных заключается в степени, в которой нужно учитывать контекстные переменные, что, в свою очередь, отражает уровень гибкости поведенческого репертуара животного. Несомненно, дополнительная корковая ткань мозга приматов, даже если принять во внимание размер тела, — это то, что позволяет ему выражать поведение менее ограниченными способами, и, несомненно, это то, что лежит в основе «интеллекта».
Ассоциативные процессы или когнитивные процессы?
Важный момент, который следует иметь в виду при сравнении производительности видов при выполнении конкретной задачи, заключается в том, что похожие графы не подразумевают схожие процессы, лежащие в основе.Тот факт, что голубь показывает уровни передачи в задаче сопоставления, аналогичной задаче обезьян, или выполняет аналогичные тесты на транзитивность, не означает, что он вызывает те же процессы для решения задачи, что и обезьяна. Аналогичный тезис был резко высказан Гэллапом в его ответе на демонстрацию Эпштейном и др. (1981) способности голубей к самопознанию, когда он заявил: «Просто потому, что вы можете имитировать поведение одного вида, усиливая серию следующих друг за другом». приближения к тому, что выглядит как один и тот же распорядок в другом, из этого не следует, что поведение первого вида обязательно возникло таким же образом »(Gallup, 1985, p.633). Несмотря на справедливую критику, простой факт состоит в том, что практически нет доказательств того, что голуби решают сложные задачи иначе, чем обезьяны, или обезьяны, иначе, чем шимпанзе, если, конечно, учитывать контекстные переменные. Тот факт, что голуби, обезьяны и шимпанзе решают задачи одинаково, подтверждается не только метрикой успешного тестирования, но и, что более важно, метрикой тестирования сигнатур, которая исследует различные сигнатуры выполнения задачи (Taylor, 2014; Шарф и Коломбо, 2020).
Важный вопрос для сравнительного познания не в том, может ли животное решать задачу или нет, а в том, как оно решает задачи ? Мы вызываем конструкции, такие как концепция соответствия , симметрия , транзитивность и орфографическая обработка , как если бы эти конструкции были объяснением поведения. Это не так, это просто ярлыки для поведения. Эпштейн и др. (1981, стр. 696) красиво сформулировали это, когда они сказали, что «такие конструкции препятствуют поиску управляющих переменных поведения, которое, как они говорят, выполняются.Соблазн подробно интерпретировать поведение животного широко распространен (Haith, 1998; Shettleworth, 2010). Говоря о нашем собственном исследовании, мы можем утверждать, что у голубей есть понятие соответствия (Colombo et al., 2003), абстрактные числовые способности (Scarf et al., 2011) и орфографическая обработка (Scarf et al., 2016), но мы не верьте, что голуби (или обезьяны) преуспевают в таких задачах, потому что у них развитые когнитивные навыки. Скорее, мы используем эти конструкции, во многом так же, как Скиннер, Эпштейн и их коллеги использовали их в исследованиях моделирования Columban (самосознание: Epstein et al., 1981; символическая коммуникация: Эпштейн и др., 1980; Insight: Epstein et al., 1984), чтобы имитировать конструкции, которые использовались с приматами, для которых нам гораздо удобнее использовать такие ярлыки.
Если не «когнитивные» процессы, то какие процессы лежат в основе этих впечатляющих способностей? Мы определенно недооцениваем силу более простых (но не простых) учетных записей, таких как процессы ассоциативного обучения или обучения с подкреплением (Dickinson, 2012; Hanus, 2016; Haselgrove, 2016). Мы сомневаемся, что наши голуби (или обезьяны) действительно участвуют в «орфографической обработке» и разбивают каждое четырехбуквенное слово, которое они видят, на составляющие его пары, и оценивают частоту, с которой каждая пара может встречаться в словах или не словах ( Grainger et al., 2012). Скорее, мы согласны с Vokey and Jamieson (2015, см. Также Linke et al., 2017) в том, что обезьяны и птицы, вероятно, отображают новые слова на прототипные «словесные» и «несловесные» шаблоны, что является впечатляющей и, конечно, непростой способностью. конечно, но тот, который отличается от «орфографического» описания. Точно так же мы могли бы использовать «ментальные представления» как процессы, управляющие поведением голубей и обезьян в задаче транзитивности, но более простые концепции, такие как теория передачи значений и модель двоичной выборки, имеют большое значение для объяснения поведения.Правда, эти более простые объяснения не могут объяснить все нюансы наблюдаемого поведения (а они должны), но какая часть из этого может отражать наше непонимание этих более простых отчетов, в отличие от недостатков этих более простых отчетов?
Проблема, которую мы затронули выше, является важной проблемой для сравнительного познания, и ее невозможно отдать должным в качестве примечания к нескольким абзацам. Мы согласны с Алленом (2014, с. 76) в том, что существует слишком много «трофейной охоты» и что те теории, которые доступны, не формализованы в достаточной степени, чтобы по-настоящему распутать разницу между ассоциативными и когнитивными моделями поведения.Но модели критически важны, если мы хотим продвинуться в этой области, особенно модели, основанные на процессах (Luce, 1995; Buckner, 2011). Тем не менее, сами модели не лишены ограничений. Например, Smith et al. (2016) отмечают, что ассоциативные модели, основанные в основном на принципах подкрепления, и когнитивные модели, основанные, главным образом, на реакциях неопределенности, математически одинаковы, и что, если кто-то не хочет ссылаться на канон Моргана, нет особых причин принимать одну за другой. Мы придерживаемся другого мнения, что, возможно, причина того, что эти модели математически идентичны, заключается в том, что лежащие в их основе процессы не так различны, как мы думаем; несомненно, мониторинг неопределенности тесно связан не только с нашей недавней историей, но и с историей удаленного подкрепления.Как утверждает Кристал (2011, стр. 417), «если реакция неопределенности никогда не подкреплялась, кажется маловероятным, что она была бы вызвана субъектом, и кажется практически невозможным, чтобы она использовалась функционально, чтобы выразить неопределенность или избежать трудного испытания. . »
Заключительные комментарии
Мы рассмотрели ряд исследований и, надеюсь, убедили читателя в том, что в ситуациях, когда один вид превосходит другой, причина часто может быть связана с контекстными переменными.Макфейл (1985) пришел к выводу, что он не переоценивал важность контекстных переменных, и более чем через три десятилетия мы согласимся с тем, что контекстные переменные действительно лежат в основе многих различий в производительности, наблюдаемых у разных видов. В сопроводительной статье (Scarf and Colombo, 2020) мы также показали, что сходство распространяется не только на производительность при выполнении задачи, но и на сигнатуры, лежащие в основе успешного выполнения задачи. Взятые вместе, мы полностью поддерживаем точку зрения Макфейла о том, что по крайней мере нет качественных различий между видами позвоночных и, конечно же, между птицами и обезьянами.С другой стороны, мы думаем, что существует достаточно оснований для мнения о количественных различиях между видами. Возможно, воспринимая мир через количественную призму разницы в степени, мы сможем лучше преодолеть разрыв между ассоциативными процессами и когнитивными процессами.
Авторские взносы
MC и DS разработали концепцию и написали рукопись. Все авторы внесли свой вклад в статью и одобрили представленную версию.
Финансирование
Исследование финансировалось MC Королевского общества Новой Зеландии, грант Марсдена 19-UOO-162.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Список литературы
Азеведо, Ф. А., Карвалью, Л. Р., Гринберг, Л. Т., Фарфель, Дж. М., Ферретти, Р. Е., Лейте, Р. Е. и др. (2009). Равное количество нейронных и ненейрональных клеток делает человеческий мозг изометрически увеличенным мозгом приматов. J. Comp.Neurol. 513, 532–541. DOI: 10.1002 / cne.21974
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бабб С. Дж. И Кристал Дж. Д. (2005). Дискриминация того, что, когда и где: последствия для эпизодической памяти у крыс. ЖЖ. Мотив. 36, 177–189. DOI: 10.1016 / j.lmot.2005.02.009
CrossRef Полный текст | Google Scholar
Барлоу, Х. Б. (1987). Эффективность, универсальность, когнитивные карты и язык. Behav. Brain Sci. 10, 657–658. DOI: 10.1017 / S0140525X0005500X
CrossRef Полный текст | Google Scholar
Берд, Л. Р., Робертс, В. А., Абромс, Б., Кит, К. К., и Крапи, К. (2003). Пространственная память о еде, спрятанной крысами ( Rattus norvegicus ) в радиальном лабиринте: исследования памяти о том, где, что и когда. J. Comp. Psychol. 117, 176–187. DOI: 10.1037 / 0735-7036.117.2.176
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бакнер, К.(2011). Два подхода к различию между познанием и «простой ассоциацией». Внутр. J. Comp. Psychol. 24, 314–348.
Google Scholar
Картер Д. Э. и Вернер Т. Дж. (1978). Комплексное обучение и обработка информации голубями: критический анализ. J. Exp. Анальный. Behav. 29, 565–601. DOI: 10.1901 / jeab.1978.29-565
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Коломбо М., Коттл А. и Фрост Н. (2003).Степень репрезентативности концепции соответствия у голубей ( Columba livia ). J. Comp. Psychol. 117, 246–256. DOI: 10.1037 / 0735-7036.117.3.246
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Коломбо, М., и Хейн, Х. (2010). «Эпизодическая память: вопросы сравнения и развития» в Оксфордском справочнике по развитию и сравнительной неврологии . ред. М. С. Блумберг, Дж. Х. Фриман и С. Р. Робинсон (Нью-Йорк, штат Нью-Йорк: издательство Оксфордского университета), 617–636.
Google Scholar
Кук, Р. Г., Кац, Дж. С., и Кавото, Б. Р. (1997). Обучение голубей одной и той же концепции с использованием нескольких классов стимулов. J. Exp. Psychol. Anim. Behav. Процесс. 23, 417–433. DOI: 10.1037 / 0097-7403.23.4.417
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кристалл, Дж. Д. (2011). Навигация по интерфейсу между обучением и познанием. Внутр. J. Comp. Psychol. 24, 412–436.
PubMed Аннотация | Google Scholar
Камминг, В.У. и Берриман Р. (1961). Некоторые данные о подходящем поведении голубя. J. Exp. Анальный. Behav. 4, 281–284. DOI: 10.1901 / jeab.1961.4-281
CrossRef Полный текст | Google Scholar
Д’Амато М. Р. и Коломбо М. (1988). Изображение серийного порядка у обезьян ( Cebus apella ). J. Exp. Psychol. Anim. Behav. Процесс. 14, 131–139. DOI: 10.1037 / 0097-7403.14.2.131
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Д’Амато, М.Р. и Салмон Д. П. (1984). «Познавательные процессы у обезьян Cebus» в Познание животных . ред. Х. Л. Ройтблат, Т. Г. Бевер и Х. С. Террас (Хиллсдейл, Нью-Джерси: Эрлбаум).
Google Scholar
Д’Амато М. Р., Салмон Д. П. и Коломбо М. (1985a). Степень и пределы концепции соответствия у обезьян ( Cebus apella ). J. Exp. Psychol. Anim. Behav. Процесс. 11, 35–51. DOI: 10.1037 // 0097-7403.11.1.35
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Д’Амато, М.Р., Салмон, Д. П., Лукас, Э., и Томи, А. (1985b). Симметрия и транзитивность условных отношений у обезьян ( Cebus apella ) и голубей ( Columba livia ). J. Exp. Анальный. Behav. 44, 35–47. DOI: 10.1901 / jeab.1985.44-35
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Делиус, Дж. Д., и Симанн, М. (1998). Транзитивная реакция у животных и людей: скорее экстаптация, чем адаптация. Behav. Процессы 42, 107–137.DOI: 10.1016 / S0376-6357 (97) 00072-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эдвардс, К. А., Джагиело, Дж. А., и Зенталл, Т. Р. (1983). Использование голубями символа «одинаковый / разный». Anim. Учиться. Behav. 11, 349–355. DOI: 10.3758 / BF03199787
CrossRef Полный текст | Google Scholar
Элепфандт А. (1987). Сравнительное познание: неадекватный подход, опрометчивые выводы. Behav. Brain Sci. 10, 661–662. DOI: 10.1017 / S0140525X00055059
CrossRef Полный текст | Google Scholar
Эпштейн, Р., Киршнит, К. Э., Ланза, Р. П., и Рубин, Л. С. (1984). «Проницательность» голубя: предшественники и факторы, определяющие умную производительность. Природа 308, 61–62. DOI: 10.1038 / 308061a0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эпштейн Р., Ланца Р. П. и Скиннер Б. Ф. (1980). Символическое общение двух голубей ( Columbia livia domestica ). Наука 207, 543–545. DOI: 10.1126 / science.207.4430.543
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Франк, А.Дж. И Вассерман Э. А. (2005). Ассоциативная симметрия у голубя после последовательного обучения сопоставлению с выборкой. J. Exp. Анальный. Behav. 84, 147–165. DOI: 10.1901 / jeab.2005.115-04
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гиллан Д. Дж. (1981). Рассуждения на шимпанзе: II. Переходный вывод. J. Exp. Psychol. Anim. Behav. Процесс. 7, 150–164. DOI: 10.1037 / 0097-7403.7.2.150
CrossRef Полный текст | Google Scholar
Джурфа, М., Zhang, S., Jenett, A., Menzel, R., and Srinivasan, M. V. (2001). Понятия «одинаковость» и «различие» у насекомых. Природа 410, 930–933. DOI: 10.1038 / 35073582
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Goldman-Rakic, P. S., and Preuss, T. M. (1987). Поднимется сравнительная психология? Behav. Brain Sci. 10, 666–667. DOI: 10.1017 / S0140525X00055102
CrossRef Полный текст | Google Scholar
Грейнджер, Дж., Дюфау, С., Монтант, М., Зиглер, Дж. К., и Фагот, Дж. (2012). Ортографическая обработка павианов ( Papio papio ). Наука 336, 245–248. DOI: 10.1126 / science.1218152
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хейт, М. М. (1998). Кто заложил винтик в познании младенцев? Слишком дорого обходится богатая интерпретация? Infant Behav. Dev. 21, 167–179. DOI: 10.1016 / S0163-6383 (98)-7
CrossRef Полный текст | Google Scholar
Хэмптон, Р.Р., Хэмпстед Б. М. и Мюррей Е. А. (2005). Обезьяны-резусы ( Macaca mulatta ) демонстрируют устойчивую память на то, что и где, но не когда, в тесте памяти в открытом поле. ЖЖ. Мотив. 36, 245–259. DOI: 10.1016 / j.lmot.2005.02.004
CrossRef Полный текст | Google Scholar
Ханус, Д. (2016). Случайные рассуждения против ассоциативного обучения: полезная дихотомия или соломенная битва в сравнительной психологии. J. Comp. Psychol. 130, 241–248.DOI: 10.1037 / a0040235
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Hodos, W. (1987). Общий интеллект животных: идея, опередившая свое время. Behav. Brain Sci. 10: 668. DOI: 10.1017 / S0140525X00055138
CrossRef Полный текст | Google Scholar
Хоган Д. Э. и Зенталл Т. Р. (1977). Обратные ассоциации у голубя. Am. J. Psychol. 90, 3–15. DOI: 10.2307 / 1421635
CrossRef Полный текст | Google Scholar
Лазарева, О.Ф., и Вассерман Э. А. (2012). Переходный вывод у голубей: измерение ассоциативных значений стимулов B и D. Behav. Процессы 89, 244–255. DOI: 10.1016 / j.beproc.2011.12.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Линке, М., Брекер, Ф., Рамскар, М., и Баайен, Х. (2017). Обучаются ли павианы «орфографическим» представлениям? Возможно нет. PLoS One 12: e0183876. DOI: 10.1371 / journal.pone.0183876
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лионелло-ДеНольф, К.М. и Уркуиоли П. Дж. (2002). Топографии контроля стимулов и проверка симметрии у голубей. J. Exp. Анальный. Behav. 78, 467–495. DOI: 10.1901 / jeab.2002.78-467
CrossRef Полный текст | Google Scholar
Липкенс Р., Коп П. Ф. и Маттейс В. (1988). Тест на симметрию и транзитивность в условных различительных характеристиках голубей. J. Exp. Анальный. Behav. 49, 395–409. DOI: 10.1901 / jeab.1988.49-395
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Люс, Р.Д. (1995). Четыре противоречия относительно математического моделирования в психологии. Annu. Rev. Psychol. 46, 1–27. DOI: 10.1146 / annurev.ps.46.020195.000245
CrossRef Полный текст | Google Scholar
Макфейл, Э. М. (1982). Мозг и интеллект позвоночных . Оксфорд, Англия: Clarendon Press.
Google Scholar
Макфейл, Э. М. (1985). Интеллект позвоночных: нулевая гипотеза. Philos. Пер. R. Soc. Лондон. B Biol. Sci. 308, 37–51.DOI: 10.1098 / rstb.1985.0008
CrossRef Полный текст | Google Scholar
Макфейл, Э. М. (1987). Сравнительная психология интеллекта. Behav. Brain Sci. 10, 645–695. DOI: 10.1017 / S0140525X00054984
CrossRef Полный текст | Google Scholar
Masure, R.H., и Allee, W.C. (1934). Социальный порядок в стаях обычных кур и голубей. Auk 51, 306–327. DOI: 10.2307 / 4077659
CrossRef Полный текст | Google Scholar
Макилвейн, W.Дж., Серна, Р. В., Дуб, В. В., и Стромер, Р. (2000). «Согласованность топографии контроля стимулов и эквивалентность стимулов: согласование результатов испытаний с теорией» в Экспериментальный и прикладной анализ человеческого поведения . ред. Дж. Лесли и Д. Э. Блэкман (Рино, Невада: Context Press), 85–110.
Google Scholar
Макинтайр, Р. Д., Клири, Дж., И Томпсон, Т. (1987). Условные отношения обезьян: рефлексивность, симметрия и транзитивность. J. Exp. Анальный. Behav. 47, 279–285.DOI: 10.1901 / jeab.1987.47-279
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Морено, А. М., де Соуза, Д. Д. Г., и Рейнхард, Дж. (2012). Сравнительное исследование способности к реляционному обучению у медоносных пчел ( Apis mellifera ) и пчел без жала ( Melipona rufiventris ). PLoS One 7: e51467. DOI: 10.1371 / journal.pone.0051467
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Оден, Д. Л., Томпсон, Р.К. Р. и Премак Д. (1988). Спонтанный перенос совпадения детенышей шимпанзе ( Pan troglodytes ). J. Exp. Psychol. Anim. Behav. Процесс. 14, 140–145. DOI: 10.1037 / 0097-7403.14.2.140
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пас-и-Миньо К., Г., Бонд, А. Б., Камил, А. К., и Балда, Р. П. (2004). Пиньонские сойки используют транзитивный вывод для предсказания социального доминирования. Природа 430, 778–781. DOI: 10.1038 / nature02723
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пенн, Д.К., Холяок, К. Дж., И Повинелли, Д. Дж. (2008). Ошибка Дарвина: объяснение разрыва между человеческим и нечеловеческим разумом. Behav. Brain Sci. 31, 109–130. DOI: 10.1017 / S0140525X08003543
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пиаже, Дж. (1928). Суждение и рассуждение у ребенка . Лондон, Англия: Routledge Chapman & Hall.
Google Scholar
Повинелли, Д. Дж. (1994). Сравнительные исследования атрибуции психического состояния животных: ответ Хейесу. Anim. Behav. 48, 239–241. DOI: 10.1006 / anbe.1994.1233
CrossRef Полный текст | Google Scholar
Повинелли Д. Дж., Нельсон К. Э. и Бойсен С. Т. (1990). Выводы о догадках и знании шимпанзе ( Pan troglodytes ). J. Comp. Psychol. 104, 203–210. DOI: 10.1037 / 0735-7036.104.3.203
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Премак Д. и Вудрафф Г. (1978). Есть ли у шимпанзе теория разума? Behav.Brain Sci. 1, 515–526. DOI: 10.1017 / S0140525X00076512
CrossRef Полный текст | Google Scholar
Ричардс Р. У. (1988). Вопрос о двунаправленных ассоциациях в обучении голубей задачам условной дискриминации. Бык. Психон. Soc. 26, 577–579. DOI: 10.3758 / BF03330126
CrossRef Полный текст | Google Scholar
Сэндс, С. Ф., и Райт, А. А. (1980). Производительность распознавания серийных зондов резус-деньгами и человеком с помощью списков из 10 и 20 пунктов. J. Exp. Psychol. Anim. Behav. Процесс. 6, 386–396. DOI: 10.1037 / 0097-7403.6.4.386
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сантьяго, Х.С., и Райт, А.А. (1984). Память голубя: обучение одной и той же / другой концепции, получение последовательного распознавания датчика и влияние задержки датчика на функцию последовательного позиционирования. J. Exp. Psychol. Anim. Behav. Процесс. 10, 498–512. DOI: 10.1037 / 0097-7403.10.4.498
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шарф, Д., Бой, К., Убер Райнерт, А., Дивайн, Дж., Гюнтюркюн, О., и Коломбо, М. (2016). Ортографическая обработка у голубей ( Columba livia ). Proc. Natl. Акад. Sci. США 113, 11272–11276. DOI: 10.1073 / pnas.1607870113
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шарф Д. и Коломбо М. (2008). Изображение серийного порядка: сравнительный анализ людей, обезьян и голубей. Brain Res. Бык. 76, 307–312. DOI: 10.1016 / j.brainresbull.2008.02.022
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шарф, Д., и Коломбо, М. (2020). Columban Simulation Project 2.0: числовая компетенция и орфографическая обработка у голубей и приматов. Перед. Psychol. 10: 3017. DOI: 10.3389 / fpsyg.2019.03017
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шварц Б. Л., Колон М. Р., Санчес И. К., Родригес И. А. и Эванс С. (2002). Однократное изучение информации «что» и «кто» у гориллы ( Gorilla gorilla gorilla ): последствия для эпизодической памяти. Anim. Cogn. 5, 85–90. DOI: 10.1007 / s10071-002-0132-0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шварц Б. Л., Хоффман М. Л. и Эванс С. (2005). Эпизодическая память у гориллы: обзор и новые открытия. ЖЖ. Мотив. 36, 226–244. DOI: 10.1016 / j.lmot.2005.02.012
CrossRef Полный текст | Google Scholar
Шварц, Б. Л., Мейснер, К. М., Хоффман, М., Эванс, С., и Фрейзер, Л. Д. (2004). Эффекты памяти событий и дезинформации у гориллы ( Gorilla gorilla gorilla ). Anim. Cogn. 7, 93–100. DOI: 10.1007 / s10071-003-0194-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шеттлворт, С. Дж. (1998). Познание, эволюция и поведение . Нью-Йорк, Нью-Йорк: Издательство Оксфордского университета.
Google Scholar
Сидман М. (1994). Отношения эквивалентности и поведение: исследование . Бостон, Массачусетс: Кооператив авторов.
Google Scholar
Сидман М., Раузин Р., Лазарь Р., Каннингем С., Тейлби В. и Карриган П. (1982). Поиск симметрии в условном различении макак-резусов, бабуинов и детей. J. Exp. Анальный. Behav. 37, 23–44. DOI: 10.1901 / jeab.1982.37-23
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Симанн М., Делиус Дж. Д. и Райт А. А. (1996). Переходные реакции у голубей: влияние частоты стимулов и истории подкрепления. Behav. Процессы 37, 185–195.DOI: 10.1016 / 0376-6357 (96) 00020-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сков-Ракетт, С. И., Миллер, Н. Ю., и Шеттлворт, С. Дж. (2006). Что-где-когда память у голубей. J. Exp. Psychol. Anim. Behav. Процесс. 32, 345–358. DOI: 10.1037 / 0097-7403.32.4.345
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Смит, Дж. Д., Закжевски, А. К., и Черч, Б. А. (2016). Формальные модели в исследованиях метапознания животных: проблема интерпретации поведения животных. Психон. Бык. Rev. 23, 1341–1353. DOI: 10.3758 / s13423-015-0985-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Стейрн, Дж. Н., Уивер, Дж. Э. и Зенталл, Т. Р. (1995). Переходный вывод у голубей: упрощенные процедуры и проверка теории переноса ценностей. Anim. Учиться. Behav. 23, 76–82. DOI: 10.3758 / BF03198018
CrossRef Полный текст | Google Scholar
Штернберг Р. Дж. (1987). Трудности сравнения интеллекта разных видов. Behav. Brain Sci. 10, 679–680. DOI: 10.1017 / S0140525X00055278
CrossRef Полный текст | Google Scholar
Штрауб, Р. О., и Террас, Х. С. (1981). Обобщение серийного обучения голубя. Anim. Учиться. Behav. 9, 454–468. DOI: 10.3758 / BF03209775
CrossRef Полный текст | Google Scholar
Suddendorf, T., и Corballis, M.C. (2007). Эволюция предвидения: что такое мысленные путешествия во времени и уникальны ли они для людей? Behav.Brain Sci. 30, 299–313. DOI: 10.1017 / S0140525X07001975
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Террас, Х. С. (1993). Филогения и онтогенез серийной памяти: список обучения голубей и обезьян. Psychol. Sci. 4, 162–169. DOI: 10.1111 / j.1467-9280.1993.tb00481.x
CrossRef Полный текст | Google Scholar
Томонага М., Мацузава Т., Фудзита К. и Ямамото Дж. (1991). Появление симметрии в визуальном условном различении шимпанзе ( Pan troglodytes ). Psychol. Rep. 68, 51–60. DOI: 10.2466 / pr0.1991.68.1.51
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тулвинг, Э. (1972). «Эпизодическая и смысловая память» в Организация памяти . ред. Э. Талвинг и У. Дональдсон (Нью-Йорк: Academic Press), 382–403.
Google Scholar
Vaughan, W. (1988). Формирование множеств эквивалентности у голубей. J. Exp. Psychol. Anim. Behav. Процесс. 14, 36–42. DOI: 10.1037 / 0097-7403.14.1.36
CrossRef Полный текст | Google Scholar
Воки, Дж. Р., и Джеймисон, Р. К. (2015). Отчет о визуальном знакомстве свидетельств орфографической обработки павианов ( Papio papio ). Psychol. Sci. 25, 991–996. DOI: 10.1177 / 0956797613516634
PubMed Аннотация | CrossRef Полный текст | Google Scholar
фон Ферзен, Л., Винн, К. Д. Л., Делиус, Дж. Д. и Стаддон, Дж. Э. Р. (1991). Формирование переходного вывода у голубей. Дж.Exp. Psychol. Anim. Behav. Процесс. 17, 334–341. DOI: 10.1037 / 0097-7403.17.3.334
CrossRef Полный текст | Google Scholar
Вассерман, Э.А., Хугарт, Дж. А., и Киркпатрик-Стегер, К. (1995). Голуби демонстрируют совершенно разные представления после тренировки со сложными визуальными стимулами. J. Exp. Psychol. Anim. Behav. Процесс. 21, 248–252. DOI: 10.1037 // 0097-7403.21.3.248
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вайнштейн, Б.(1941). Подбор по образцу макаками-резусами и детьми. J. Comp. Psychol. 31, 195–213. DOI: 10,1037 / h0063449
CrossRef Полный текст | Google Scholar
Уилсон, Б., Макинтош, Н. Дж., И Боукс, Р. А. (1985a). Сопоставление и необычное обучение у голубя: эффекты переноса и отсутствие реляционного обучения. Q. J. Exp. Psychol. В 37, 295–311. DOI: 10.1080 / 14640748508401172
CrossRef Полный текст | Google Scholar
Уилсон, Б., Макинтош, Н. Дж., И Боукс, Р. А. (1985b). Передача реляционных правил в сопоставлении и изучении странностей голубями и врановыми. Q. J. Exp. Psychol. В 37, 313–332. DOI: 10.1080 / 14640748508401173
CrossRef Полный текст | Google Scholar
Райт, А.А. (1997). Концептуальное обучение и стратегии обучения. Psychol. Sci. 8, 119–123. DOI: 10.1111 / j.1467-9280.1997.tb00693.x
CrossRef Полный текст | Google Scholar
Ямамото Дж. И Асано Т.(1995). Эквивалентность стимулов у шимпанзе ( Pan troglodytes ). Psychol. Рек. 45, 3–21.
Google Scholar
Янг, М. Э., Вассерман, Э. А., и Гарнер, К. Л. (1997). Влияние количества предметов на различение голубей одного и того же на разных визуальных дисплеях. J. Exp. Psychol. Anim. Behav. Процесс. 23, 491–501. DOI: 10.1037 / 0097-7403.23.4.491
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Зенталл, Т.Р. и Хоган Д. Э. (1974). Абстрактное обучение концепции в голуби. J. Exp. Psychol. 102, 393–398. DOI: 10,1037 / h0035970
CrossRef Полный текст | Google Scholar
Зенталл Т. Р. и Хоган Д. Э. (1978). Одно и то же / другое концептуальное обучение у голубя: влияние негативных примеров и предшествующая адаптация к передаче стимулов. J. Exp. Анальный. Behav. 30, 177–186. DOI: 10.1901 / jeab.1978.30-177
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Развитие высокоскоростного метания
Мы обнаружили, что люди могут метать с такой скоростью, сохраняя упругую энергию в своих плечах.Это достигается за счет размещения руки таким образом, чтобы масса руки сопротивлялась движениям туловища и плеча и вращалась назад от цели. Этот «взвод» руки растягивает сухожилия, связки и мышцы, пересекающие плечо, и накапливает энергию упругости (как в рогатке). Когда эта энергия затем высвобождается, она обеспечивает очень быстрое вращение плеча, что является самым быстрым движением человеческого тела — до 9000 градусов в секунду у профессиональных питчеров! Это быстрое вращение также приводит к быстрому выпрямлению локтя и выпуску снаряда с очень высокой скоростью.
Мы обнаружили, что люди могут метать с такой скоростью, сохраняя упругую энергию в своих плечах. Это достигается за счет размещения руки таким образом, чтобы масса руки сопротивлялась движениям туловища и плеча и вращалась назад от цели. Этот «взвод» руки растягивает сухожилия, связки и мышцы, пересекающие плечо, и накапливает энергию упругости (как в рогатке). Когда эта энергия затем высвобождается, она обеспечивает очень быстрое вращение плеча, что является самым быстрым движением человеческого тела — до 9000 градусов в секунду у профессиональных питчеров! Это быстрое вращение также приводит к быстрому выпрямлению локтя и выпуску снаряда с очень высокой скоростью.
Бывший питчер MLB Билли Вагнер показывает огромное вращение руки во время фазы «взведения», когда сохраняется упругая энергия. (Изображение предоставлено: sportsofboston.com)
Мы также обнаружили, что три ключевых анатомических изменения, произошедших в ходе эволюции человека, сделали возможным этот новый механизм накопления энергии: расширение талии, опускание плеч и низкий перекрут плечевой кости. Расширение талии позволяет туловищу вращаться независимо от бедер.Это вращение туловища создает большие силы, необходимые для растяжения эластичных сухожилий и связок плеча. Опускание плеча изменяет ориентацию многих мышц плеча, включая большую грудную мышцу (большую грудную мышцу), которая имеет решающее значение для накопления энергии. Наконец, мы обнаружили, что низкий перекрут плечевой кости (перекручивание кости плеча) позволяет нам накапливать больше энергии и, таким образом, быстрее метать.
_________________________________________________________________________________
.