Парадоксальный сон: зачем нужны сновидения и почему их помнят не все
Откуда берутся сновидения? Для чего они нужны? Об этом и не только рассказывал в интервью Psychologies.ru первооткрыватель фазы парадоксального сна (быстрый сон, REM-сон. — Прим. Vokrugsveta.ru) профессор Мишель Жуве (1925-2017).
- Фото
- BSIP/Universal Images Group via Getty Images
Мишель Жуве
Нейрофизиолог и невролог, один из трех «отцов-основателей» современной сомнологии, член Национальной академии наук Франции. Жуве руководил исследованиями природы сна и сновидений во Французском национальном институте здоровья и медицинских исследований.
Cновидения появляются во время парадоксального сна. Что это такое и как вам удалось открыть существование этой фазы?
Мишель Жуве: Парадоксальный сон был открыт нашей лабораторией в 1959 году. Изучая формирование условных рефлексов у кошек, мы неожиданно зафиксировали удивительный феномен, нигде ранее не описанный. У спящего животного наблюдались быстрые движения глаз, интенсивная мозговая деятельность, почти как во время бодрствования, при этом мышцы были полностью расслаблены. Это открытие перевернуло все наши представления о снах.
Изучая формирование условных рефлексов у кошек, мы неожиданно зафиксировали удивительный феномен, нигде ранее не описанный. У спящего животного наблюдались быстрые движения глаз, интенсивная мозговая деятельность, почти как во время бодрствования, при этом мышцы были полностью расслаблены. Это открытие перевернуло все наши представления о снах.
Прежде считалось, что сновидение — это серия коротких образов, которые человек видит непосредственно перед пробуждением
Открытое нами состояние организма — это не классический сон и не бодрствование, а особое, третье состояние. Мы назвали его «парадоксальным сном», потому что в нем парадоксально сочетаются полное расслабление мышц тела и интенсивная активность мозга; это активное бодрствование, направленное внутрь себя.
Сколько раз за ночь человек видит сны?
Четыре-пять. Продолжительность первых снов — не больше 18–20 минут, последние два «сеанса» более длительные, по 25–30 минут. Мы обычно запоминаем самый последний сон, который обрывается с нашим пробуждением. Он может быть длинным или состоять из четырех-пяти коротких эпизодов — и тогда нам кажется, будто мы видели сны всю ночь.
Мы обычно запоминаем самый последний сон, который обрывается с нашим пробуждением. Он может быть длинным или состоять из четырех-пяти коротких эпизодов — и тогда нам кажется, будто мы видели сны всю ночь.
В общей сложности все наши ночные сновидения длятся около 90 минут. Их продолжительность зависит от возраста. У новорожденных сновидения составляют 60% от всего времени их сна, а у взрослых — только 20%. Именно поэтому некоторые ученые утверждают, что сон играет важную роль в созревании мозга.
Вы также открыли, что в сновидении задействовано два вида памяти…
Я сделал этот вывод, анализируя свои собственные сны — 6600, между прочим! Было уже известно, что сновидения отражают события прошедшего дня, переживания последней недели. Но вот вы отправляетесь, скажем, на Амазонку.
В первую неделю путешествия действие ваших снов будет происходить в домашних «декорациях»
А их героем вполне может оказаться индеец, расположившийся в вашей квартире. Этот пример показывает, что в создании наших сновидений участвует не только кратковременная память на ближайшие события, но и память долговременная.
Этот пример показывает, что в создании наших сновидений участвует не только кратковременная память на ближайшие события, но и память долговременная.
Исследование сна методом полисомнографии
- Фото
- BSIP/Universal Images Group via Getty Images
Почему некоторые люди не запоминают свои сны?
Среди нас таких процентов двадцать. Человек не помнит своих снов в двух случаях. Первый — если он проснулся через несколько минут после окончания сновидения, за это время оно улетучивается из памяти. Другое объяснение дает психоанализ: человек просыпается, а его «Я» — одна из основных структур личности — подвергает жесткой цензуре образы, «всплывшие» из бессознательного. И все забывается.
Из чего состоит сновидение?
На 40% — из впечатлений прожитого дня, а в остальном — из сцен, связанных с нашими страхами, тревогами, заботами. Бывают особые сны, во время которых спящий осознает, что действие происходит не наяву; бывают — почему бы и нет? — и вещие сновидения.
Бывают особые сны, во время которых спящий осознает, что действие происходит не наяву; бывают — почему бы и нет? — и вещие сновидения.
Недавно я изучал сны двух африканцев. Они во Франции давно, но каждую ночь им снится родная Африка. Тема сновидений наукой далеко не исчерпана, и каждое новое исследование это только подтверждает.
После 40 лет исследований можете ли вы ответить на вопрос, для чего человеку нужны сны?
Разочарую вас — нет! Это по-прежнему загадка. Нейрофизиологи не знают, для чего нужны сны, точно так же как не могут дать точное определение сознанию. Долгое время считалось, что сны нужны, чтобы наполнять кладовые нашей памяти.
Затем обнаружили, что в отсутствие фазы парадоксального сна и сновидений человек не испытывает проблем ни с памятью, ни с мышлением
Английский биофизик Фрэнсис Крик выдвинул противоположную гипотезу: сны помогают забывать! То есть мозг, подобно суперкомпьютеру, использует сны, чтобы стереть незначащие воспоминания. Но в таком случае человек, не видящий снов, имел бы серьезные нарушения памяти. А это не так.
Но в таком случае человек, не видящий снов, имел бы серьезные нарушения памяти. А это не так.
В теории вообще множество белых пятен. Например, во время фазы парадоксального сна наш организм потребляет больше кислорода, чем во время бодрствования. И никто не знает почему!
Мишель Жуве, 1991 год
- Фото
- Louis MONIER/Gamma-Rapho via Getty Images
Вы выдвинули гипотезу о том, что сны поддерживают наш мозг в рабочем состоянии.
Скажу больше: в снах зарождается завтрашний день, они его готовят. Их действие можно сравнить с приемом мысленной визуализации: так, накануне соревнований лыжник с закрытыми глазами мысленно проходит всю трассу. Если замерить деятельность его мозга с помощью приборов, мы получим те же данные, как если бы он уже был на трассе!
Во время фазы парадоксального сна идут те же мозговые процессы, что и у бодрствующего человека.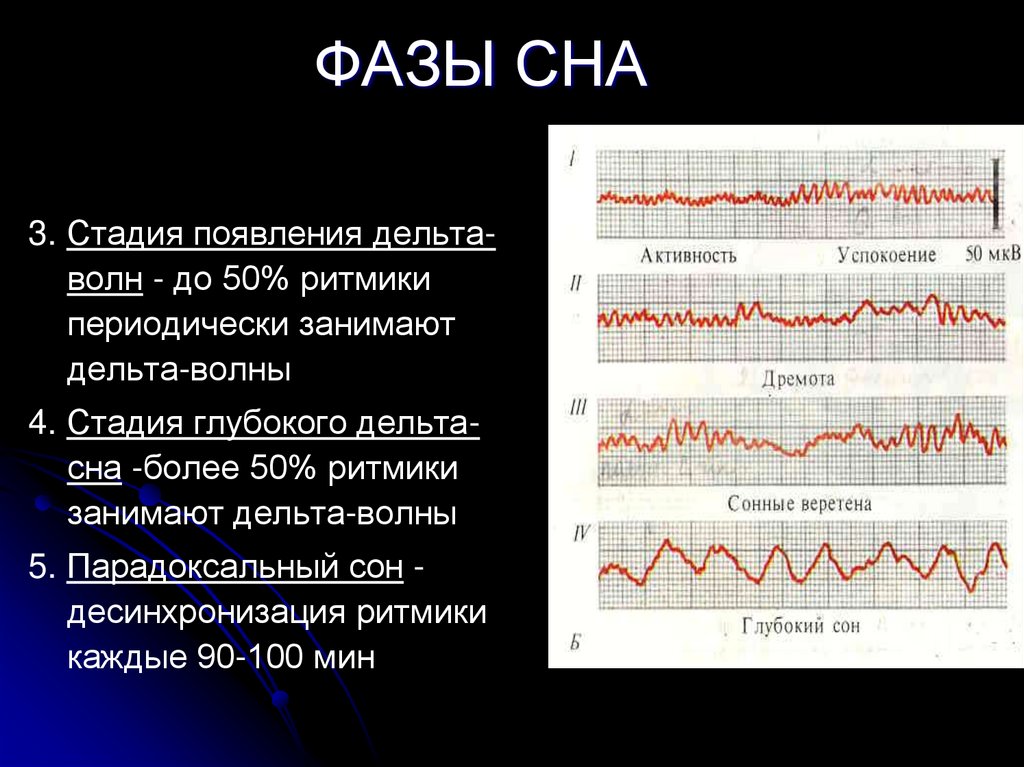 И днем наш мозг быстро активизирует ту часть нейронов, которая была задействована во время ночных сновидений. Таким образом, сны облегчают некоторые процессы обучения и напрямую связаны с нашим будущим. Можно перефразировать афоризм: я вижу сны, следовательно, будущее существует!
И днем наш мозг быстро активизирует ту часть нейронов, которая была задействована во время ночных сновидений. Таким образом, сны облегчают некоторые процессы обучения и напрямую связаны с нашим будущим. Можно перефразировать афоризм: я вижу сны, следовательно, будущее существует!
Редакция
Теги
- наука
Сегодня читают
Тест: посмотрите на свой нос и узнайте важное о своем характере
Тест: линии на запястьях расскажут о сильных сторонах вашей личности
Тест: срочно посмотрите на свои пальцы, вдруг вы скрытый гений?
Тест: ответьте на 5 вопросов, а нейросеть нарисует дом вашей мечты
Тест: попросите человека скрестить руки и узнайте все о его характере
Эволюция сна от простых форм до млекопитающих uMEDp
Cон и бодрствование – два состояния всех живых организмов от простых форм с децентрализованной нервной системой до высших млекопитающих со сложно устроенным головным мозгом. У всех животных состояние сна характеризуется похожими поведенческими, нейронными, молекулярными и физиологическими признаками. У низших позвоночных есть стадия сна, которая по многим параметрам напоминает медленноволновый сон млекопитающих. В то же время две дифференцированные стадии сна – медленноволновый сон и парадоксальный (или быстрый, или фаза быстрого сна) сон – есть только у птиц и млекопитающих. У птиц, а также у примитивных однопроходных и сумчатых млекопитающих медленноволновый сон и парадоксальный сон в целом менее дифференцированы, чем у плацентарных млекопитающих. У морских млекопитающих (китообразных и ластоногих семейства ушастых тюленей) медленноволновый сон может развиваться только в одном полушарии мозга (однополушарный сон). Эпизоды с выраженной межполушарной асимметрией электроэнцефалограммы, имеющие определенное сходство с однополушарным сном, есть и у некоторых птиц. Предполагается, что однополушарный сон позволяет сохранять двигательную активность и поддерживать бдительность во время сна.
У всех животных состояние сна характеризуется похожими поведенческими, нейронными, молекулярными и физиологическими признаками. У низших позвоночных есть стадия сна, которая по многим параметрам напоминает медленноволновый сон млекопитающих. В то же время две дифференцированные стадии сна – медленноволновый сон и парадоксальный (или быстрый, или фаза быстрого сна) сон – есть только у птиц и млекопитающих. У птиц, а также у примитивных однопроходных и сумчатых млекопитающих медленноволновый сон и парадоксальный сон в целом менее дифференцированы, чем у плацентарных млекопитающих. У морских млекопитающих (китообразных и ластоногих семейства ушастых тюленей) медленноволновый сон может развиваться только в одном полушарии мозга (однополушарный сон). Эпизоды с выраженной межполушарной асимметрией электроэнцефалограммы, имеющие определенное сходство с однополушарным сном, есть и у некоторых птиц. Предполагается, что однополушарный сон позволяет сохранять двигательную активность и поддерживать бдительность во время сна.
Рис. 1. Электрофизиологические признаки сна у домашней курицы: электроэнцефалограмма (ЭЭГ), электромиограмма (ЭМГ) шейной мускулатуры, электроокулограмма (ЭОГ), а также импульсная активность нейрона в ретикулярной формации покрышки мозга
Рис. 2. Особенности сна северного морского котика: позы сна на суше и в воде (фото С. Артемьевой и С. Корневой)
Рис. 3. Развитие медленноволнового сна у морского котика во время сна на суше (А) и в воде (Б): электроэнцефалограмма (ЭЭГ) в левом (Л) и правом (П) полушариях и электромиограмма (ЭМГ) шейных мышц
Рис. 4. Суммарная длительность ПС (А), длительность эпизодов ПС (Б), а также пропорция билатерально-симметричного МС (В) у котиков на суше, в воде, а также после возвращения на сушу (средние значения)
У млекопитающих и птиц сон состоит из двух подстадий – медленноволнового сна (МС) и парадоксального сна (ПС) (или быстрого сна, или стадии быстрых движений глаз).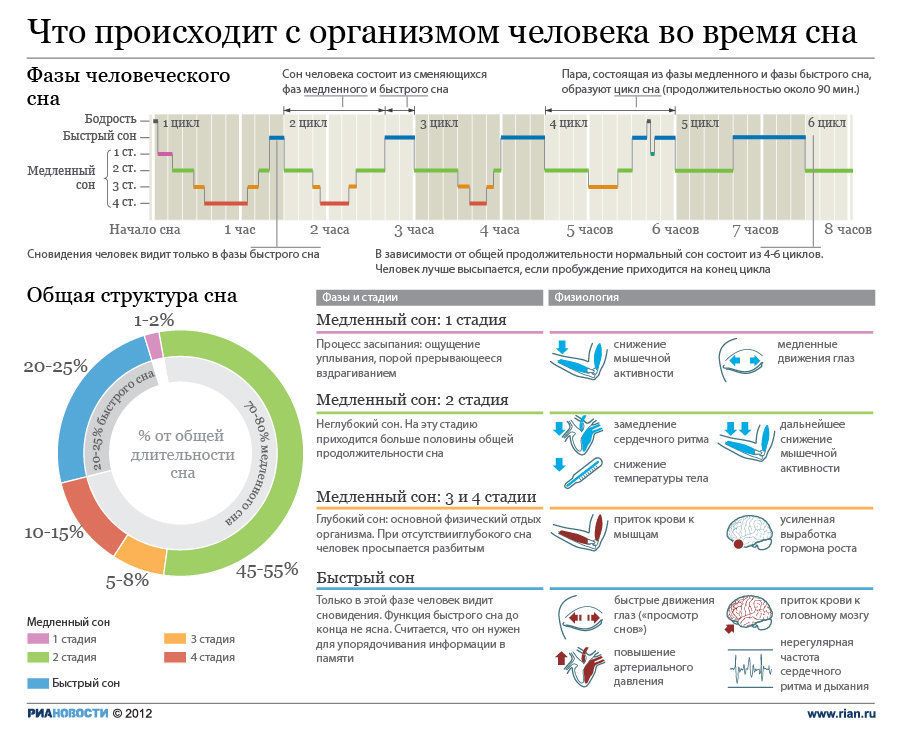 Разные виды млекопитающих проводят во сне от двух до 20 часов в сутки, а птицы – от пяти до 17 часов. Попытки установить корреляции параметров сна с анатомо-физиологическими характеристиками млекопитающих и птиц, а также с условиями сна до сих пор были не очень успешными [1–6]. Во многом это определяется тем, что данные о продолжительности сна и его стадий получены в основном в лабораторных условиях. Они не учитывают все многообразие внешних факторов, влияющих на сон животных в природе. Корреляционные отношения не всегда указывают и на причинно-следственные связи. Приходится признать, что функция и биологическая роль сна до конца не понятны. Задача этого обзора – обобщить результаты наиболее интересных сравнительно-физиологических исследований сна, выполненных в последние два десятилетия. Дополнительная информация может быть найдена в оригинальных работах и других обзорах, ссылки на которые находятся в списке цитируемой литературы.
Разные виды млекопитающих проводят во сне от двух до 20 часов в сутки, а птицы – от пяти до 17 часов. Попытки установить корреляции параметров сна с анатомо-физиологическими характеристиками млекопитающих и птиц, а также с условиями сна до сих пор были не очень успешными [1–6]. Во многом это определяется тем, что данные о продолжительности сна и его стадий получены в основном в лабораторных условиях. Они не учитывают все многообразие внешних факторов, влияющих на сон животных в природе. Корреляционные отношения не всегда указывают и на причинно-следственные связи. Приходится признать, что функция и биологическая роль сна до конца не понятны. Задача этого обзора – обобщить результаты наиболее интересных сравнительно-физиологических исследований сна, выполненных в последние два десятилетия. Дополнительная информация может быть найдена в оригинальных работах и других обзорах, ссылки на которые находятся в списке цитируемой литературы.
Сон у беспозвоночных
На сегодняшний день состояние поведенческого сна изучено у нескольких десятков видов беспозвоночных [7–9].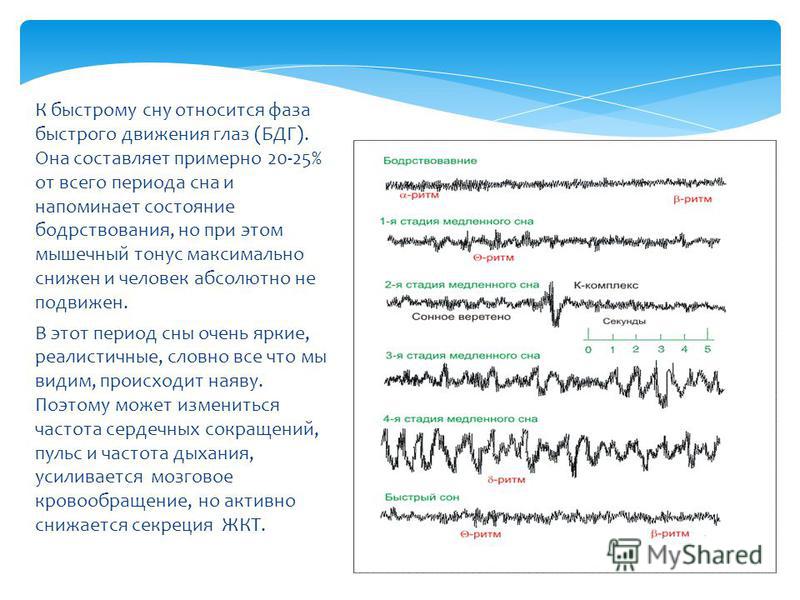 У всех исследованных видов поведенческий сон имеет такие же признаки, как у млекопитающих и птиц, включая неподвижность, типичную позу, сниженную реакцию на раздражители. Продолжительность сна увеличивается после навязанной двигательной активности (признак гомеостатической регуляции). Препараты с седативным эффектом удлиняют время сна, а стимуляторы, наоборот, сокращают. Как и у млекопитающих, наибольшее количество сна у беспозвоночных отмечается на ранних этапах развития и постепенно снижается с возрастом [8–13]. Формальные поведенческие характеристики сна наблюдаются даже у медуз – простых форм с децентрализованной нервной системой, которые существуют на Земле уже более 600 млн лет [14]. Следовательно, основные признаки поведенческого сна у животных сформировались в эволюции еще до появления централизованной нервной системы.
У всех исследованных видов поведенческий сон имеет такие же признаки, как у млекопитающих и птиц, включая неподвижность, типичную позу, сниженную реакцию на раздражители. Продолжительность сна увеличивается после навязанной двигательной активности (признак гомеостатической регуляции). Препараты с седативным эффектом удлиняют время сна, а стимуляторы, наоборот, сокращают. Как и у млекопитающих, наибольшее количество сна у беспозвоночных отмечается на ранних этапах развития и постепенно снижается с возрастом [8–13]. Формальные поведенческие характеристики сна наблюдаются даже у медуз – простых форм с децентрализованной нервной системой, которые существуют на Земле уже более 600 млн лет [14]. Следовательно, основные признаки поведенческого сна у животных сформировались в эволюции еще до появления централизованной нервной системы.
У нескольких видов беспозвоночных были зарегистрированы также электрофизиологические признаки сна. Например, снижение двигательной активности и рост порогов пробуждения у фруктовой мушки происходили параллельно уменьшению амплитуды электрической активности (локальные потенциалы в диапазоне 11–40 Гц) грибовидного тела и замедлению уровня метаболизма.
Состояние сна у беспозвоночных – не однородный процесс. В этом тоже есть сходство со сном млекопитающих и птиц. Например, у пчелы в течение всего ночного периода покоя тонус шейных мышц изменяется циклически, временами практически достигая нуля [10]. У дрозофилы величина порогов пробуждения зависит от длительности эпизода сна [16].
У дрозофилы величина порогов пробуждения зависит от длительности эпизода сна [16].
В литературе встречаются упоминания о стереотипных движениях глаз у находящихся в состоянии поведенческого сна каракатиц [20], а также подергивании антенн и конечностей у пчел [21] и муравьев [22]. Во всех работах обращается внимание на некоторое сходство этих движений со вздрагиваниями у млекопитающих в ПС. Однако подобное поведение у беспозвоночных никогда детально не анализировалось.
Таким образом, по поведенческим и некоторым физиологическим критериям состояние сна есть у всех исследованных беспозвоночных, включая организмы с децентрализованной нервной системой. По формальным признакам это состояние имеет сходство с МС млекопитающих и птиц.
Сон у рыб
Эпизоды неподвижности определенной длительности (поведенческий сон) у рыб характеризуются ухудшением способности реагировать на внешние раздражители. Поведенческий сон рыб имеет признаки гомеостатической и циркадианной регуляции. Его суммарная длительность увеличивается под действием мелатонина и диазепама и уменьшается под действием гипокретина. Наибольшая общая продолжительность эпизодов сна у рыб наблюдается на ранних этапах постнатального развития и постепенно снижается с возрастом [23–25].
Его суммарная длительность увеличивается под действием мелатонина и диазепама и уменьшается под действием гипокретина. Наибольшая общая продолжительность эпизодов сна у рыб наблюдается на ранних этапах постнатального развития и постепенно снижается с возрастом [23–25].
Длительность поведенческого сна у рыб варьируется в большом диапазоне. Например, у обитающих в пещерных водоемах мексиканских рыбок ( Astyanax mexicanus) на поведенческий сон приходится в пять раз меньше времени, чем у рыб того же вида, живущих в реках [26]. Меньшая продолжительность сна у рыб пещерных популяций определяется большим числом гипокретинсодержащих нейронов в гипоталамусе. Торможение активности нейронов приводит к увеличению времени сна [27]. Разрушение рецепторов боковой линии, наоборот, повышает продолжительность сна у пещерных форм до уровня речных рыбок, что указывает на важную роль сенсорной информации в регуляции количества сна [8]. Результаты этих исследований раскрывают молекулярно-генетические и нейронные механизмы регуляции продолжительности сна, а также консервативность нейрохимических систем мозга, регулирующих фенотип сна у разных животных. Эволюция сна пещерной формы мексиканской рыбки, которая происходила в течение примерно 2–3 млн лет, – прекрасный пример влияния экологических факторов (дефицит корма, обитание в темноте) на продолжительность сна и его суточную динамику.
Эволюция сна пещерной формы мексиканской рыбки, которая происходила в течение примерно 2–3 млн лет, – прекрасный пример влияния экологических факторов (дефицит корма, обитание в темноте) на продолжительность сна и его суточную динамику.
Данные об электрофизиологических аспектах сна у рыб противоречивы. Медленные волны и острые спайки были зафиксированы во время поведенческого сна у сома [28]. В других работах однонаправленных изменений электрической активности не регистрировалось [7]. В одном из первых исследований сна у рыб были описаны стереотипные движения глаз и сделано предположение о наличии у них ПС [29]. Однако позже эти результаты не подтвердились [28, 30]. Анатомические и молекулярные исследования показали большое сходство в организации активирующих (холинергической, моноаминергической, гипокретиновой) и сомногенных (мелатониновой) систем мозга у рыб и позвоночных [24, 31].
Сон у амфибий
Исследований сна у амфибий крайне мало.
Сон у рептилий
Высокоамплитудные спайки на электроэнцефалограмме (ЭЭГ), напоминающие медленные волны у млекопитающих, были зарегистрированы на фоне поведенческого сна у многих видов пресмыкающихся, включая каймана, игуану, черепаху и змею [7, 32]. Фармакологические исследования продемонстрировали, что реакция ЭЭГ-спайков у рептилий на сомногенные препараты такая же, как и гиппокампальных спайков (аналогов корковой медленноволновой ЭЭГ) у кошки. Частота ЭЭГ-спайков у рептилий также увеличивалась после навязанного бодрствования [33].
Признаки ПС у рептилий в ранних исследованиях отмечались редко. Одним из аргументов в пользу отсутствия этой стадии сна у рептилий было и наше исследование динамики разряда изолированных нейронов в цикле «сон – бодрствование» у черепах. Нам не удалось обнаружить признаки повторяющейся активации нейронов ретикулярной формации – ключевого признака ПС у млекопитающих ‒ у черепах во время периодов естественного сна [34]. Тем не менее как минимум у двух видов черепах сон был разделен на две стадии – МС и ПС [35, 36]. Недавно у двух видов ящериц были зарегистрированы повторяющиеся движения глаз во время поведенческого сна. Эпизоды наблюдались с интервалом 80 секунд и сопровождались снижением амплитуды ЭЭГ-спайков. Флуоксетин (ингибитор обратного транспорта серотонина, укорачивает время ПС у млекопитающих) уменьшал амплитуду ЭЭГ-спайков [37, 38]. По мнению авторов, такие эпизоды имеют признаки ПС млекопитающих. Природа регулярного чередования двух стадий сна с подобным коротким периодом не понятна и не имеет аналогов у млекопитающих и птиц.
Одним из аргументов в пользу отсутствия этой стадии сна у рептилий было и наше исследование динамики разряда изолированных нейронов в цикле «сон – бодрствование» у черепах. Нам не удалось обнаружить признаки повторяющейся активации нейронов ретикулярной формации – ключевого признака ПС у млекопитающих ‒ у черепах во время периодов естественного сна [34]. Тем не менее как минимум у двух видов черепах сон был разделен на две стадии – МС и ПС [35, 36]. Недавно у двух видов ящериц были зарегистрированы повторяющиеся движения глаз во время поведенческого сна. Эпизоды наблюдались с интервалом 80 секунд и сопровождались снижением амплитуды ЭЭГ-спайков. Флуоксетин (ингибитор обратного транспорта серотонина, укорачивает время ПС у млекопитающих) уменьшал амплитуду ЭЭГ-спайков [37, 38]. По мнению авторов, такие эпизоды имеют признаки ПС млекопитающих. Природа регулярного чередования двух стадий сна с подобным коротким периодом не понятна и не имеет аналогов у млекопитающих и птиц.
Таким образом, у рептилий, по всей видимости, есть стадия сна, напоминающая по параметрам МС млекопитающих. Убедительных данных о существовании у рептилий ПС нет. Рептилии дали начало общему предку млекопитающих и птиц. Если ПС у рептилий отсутствует, значит, он появился в эволюции этих двух групп независимо.
Убедительных данных о существовании у рептилий ПС нет. Рептилии дали начало общему предку млекопитающих и птиц. Если ПС у рептилий отсутствует, значит, он появился в эволюции этих двух групп независимо.
Сон у птиц
Электрофизиологическим методом сон изучен примерно у 30 видов класса птиц, большинство из которых принадлежат к надотряду новонёбных (Neognathae), и лишь два вида (африканский страус и тинаму) относятся ко второму надотряду – бескилевых, или древненёбных (Palaeognathae). Новонёбные насчитывают более 10 тыс. видов современных птиц, а бескилевые, или древненёбные, – базовая группа, включающая всего 60 видов современных птиц. Их предки отделились от основной ветви, давшей начало новонёбным птицам, примерно 110 млн лет назад [39].
Только у птиц и млекопитающих сон состоит из двух подстадий – МС и ПС. Главный признак МС у птиц – характерные медленные колебания в ЭЭГ в диапазоне 1–4 Гц. Другие ритмы (например, сонные веретена) у птиц обычно не выделяются. У большинства исследованных видов медленные волны в ЭЭГ развиваются в двух полушариях синхронно (билатерально-симметричный МС). Как минимум у семи видов в МС были зарегистрированы эпизоды межполушарной асимметрии ЭЭГ, или однополушарный сон. В то же время длительность этих эпизодов, как правило, составляет несколько секунд [3, 40–44] и только у некоторых видов более одной минуты [43, 45]. Эпизоды межполушарной асимметрии ЭЭГ у птиц часто сочетаются с асимметричным состоянием глаз. Например, утки, которые спят на краю группы, проводят больше времени с одним открытым глазом по сравнению с птицами, которые спят в центре группы в окружении других особей. При этом глаз, который направлен от центра группы, находится в открытом состоянии больше времени, чем глаз, обращенный на других птиц. Асимметричное состояние глаз коррелирует с эпизодами межполушарной асимметрии ЭЭГ: в полушарии, противоположном закрытому глазу, медленные волны ЭЭГ имели более высокую амплитуду, чем в полушарии, противоположном открытому глазу [44].
У большинства исследованных видов медленные волны в ЭЭГ развиваются в двух полушариях синхронно (билатерально-симметричный МС). Как минимум у семи видов в МС были зарегистрированы эпизоды межполушарной асимметрии ЭЭГ, или однополушарный сон. В то же время длительность этих эпизодов, как правило, составляет несколько секунд [3, 40–44] и только у некоторых видов более одной минуты [43, 45]. Эпизоды межполушарной асимметрии ЭЭГ у птиц часто сочетаются с асимметричным состоянием глаз. Например, утки, которые спят на краю группы, проводят больше времени с одним открытым глазом по сравнению с птицами, которые спят в центре группы в окружении других особей. При этом глаз, который направлен от центра группы, находится в открытом состоянии больше времени, чем глаз, обращенный на других птиц. Асимметричное состояние глаз коррелирует с эпизодами межполушарной асимметрии ЭЭГ: в полушарии, противоположном закрытому глазу, медленные волны ЭЭГ имели более высокую амплитуду, чем в полушарии, противоположном открытому глазу [44]. Фрегаты во время полетов над океаном на кормовые поля спят в воздухе, планируя по спирали. Вместе с тем полушарие мозга, противоположное направлению вращения, больше времени находится в состоянии бодрствования или низкоамплитудного МС. Косвенно это говорит о том, что во время сна птицы открывают глаз, который направлен в сторону вращения [45]. Поэтому функция МС с межполушарной асимметрией ЭЭГ у птиц состоит, по-видимому, в обработке информации, поступающей от открытого в сторону опасности глаза, и, следовательно, поддержании бдительности во время сна [40–43]. Считается, что основные признаки ПС млекопитающих (быстрые движения глаз, мышечные вздрагивания, торможение скелетной мускулатуры) есть и у птиц. Однако у большинства видов птиц ПС не сопровождается заметным снижением мышечного тонуса по сравнению с МС. У всех исследованных видов птиц, кроме африканского страуса, эпизоды ПС длятся всего лишь несколько секунд [3, 40–42]. Применяя традиционные полиграфические критерии, сложно точно определить границы таких эпизодов ПС.
Фрегаты во время полетов над океаном на кормовые поля спят в воздухе, планируя по спирали. Вместе с тем полушарие мозга, противоположное направлению вращения, больше времени находится в состоянии бодрствования или низкоамплитудного МС. Косвенно это говорит о том, что во время сна птицы открывают глаз, который направлен в сторону вращения [45]. Поэтому функция МС с межполушарной асимметрией ЭЭГ у птиц состоит, по-видимому, в обработке информации, поступающей от открытого в сторону опасности глаза, и, следовательно, поддержании бдительности во время сна [40–43]. Считается, что основные признаки ПС млекопитающих (быстрые движения глаз, мышечные вздрагивания, торможение скелетной мускулатуры) есть и у птиц. Однако у большинства видов птиц ПС не сопровождается заметным снижением мышечного тонуса по сравнению с МС. У всех исследованных видов птиц, кроме африканского страуса, эпизоды ПС длятся всего лишь несколько секунд [3, 40–42]. Применяя традиционные полиграфические критерии, сложно точно определить границы таких эпизодов ПС.
Сон африканского страуса отличается от сна всех исследованных птиц [46], включая еще одного представителя древненёбных птиц – тинаму [47]. У взрослых страусов эпизоды ПС могут достигать пяти минут, более чем в десять раз превышая максимальную продолжительность эпизода ПС у других птиц. При этом на ПС у страусов приходится в среднем 24% от времени ночного периода, или 26% от общего времени сна, то есть значительно больше, чем у других птиц [3]. Интересно, что характерные для ПС вздрагивания глаз и головы регистрируются на фоне как низкоамплитудной, так и высокоамплитудной ЭЭГ. Подобные особенности сна у страуса послужили основанием для гипотезы о том, что его недифференцированный сон является предшественником «эволюционно более молодого» ПС млекопитающих и птиц, который сопровождается активацией коры переднего мозга. Поэтому, возможно, ПС сформировался в эволюции птиц и млекопитающих независимо [46]. Однако отсутствие такой же недифференцированной стадии сна у тинаму и короткие эпизоды их ПС (как у всех новонёбных птиц) ставят эту гипотезу под сомнение.
Большинство нейронов в ретикулярной формации ствола мозга млекопитающих разряжаются с наибольшей частотой во время бодрствования и ПС, снижая частоту разряда в МС. Другими словами, активация подкорковых структур мозга млекопитающих – один из важнейших признаков ПС. Нами была исследована особенность импульсной активности нейронов ствола мозга во время цикла «сон – бодрствование» у домашних кур. Оказалось, что большинство таких нейронов разряжались с наибольшей частотой во время эпизодов ПС. Периоды разряда нейронов не всегда точно соответствовали другим признакам ПС [48]. В наибольшей степени учащение частоты разряда совпадало с быстрыми движениями глаз и в наименьшей – с изменением мышечного тонуса. Поэтому в этом случае правильнее использовать название «сон с быстрыми движениями глаз», так как именно этот параметр является определяющим для этого состояния у птиц. Некоторые эпизоды быстрых движений глаз у кур регистрировались на фоне медленных волн в ЭЭГ, то есть без активации переднего мозга (рис. 1). Это означает, что даже у кур – типичных представителей новонёбных ПС менее дифференцирован, чем у наземных млекопитающих. Таким образом, сложившееся представление о том, что суммарная длительность ПС/сна с быстрыми движениями глаз у птиц меньше, чем у млекопитающих, возможно, ошибочно, если в дополнение к полиграфическим критериям применять параметр активации нейронов ретикулярной формации.
1). Это означает, что даже у кур – типичных представителей новонёбных ПС менее дифференцирован, чем у наземных млекопитающих. Таким образом, сложившееся представление о том, что суммарная длительность ПС/сна с быстрыми движениями глаз у птиц меньше, чем у млекопитающих, возможно, ошибочно, если в дополнение к полиграфическим критериям применять параметр активации нейронов ретикулярной формации.
Многие птицы совершают сезонные миграции, а также полеты на кормовые поля, причем они находятся в воздухе несколько дней или даже недель. Птицы могут спать в полете, но общая продолжительность сна при этом, скорее всего, значительно меньше таковой на земле. Например, у фрегатов общее время сна в полете составляет менее 7% от продолжительности сна в гнездах [45]. Это один из примеров адаптивного сокращения продолжительности сна в ситуации, когда сон препятствует реализации более важной стратегии поведения, а именно кормлению и обеспечению безопасности. В других случаях птицы сокращают количество сна в период спаривания. В результате самцы, которые спят меньше, спариваются с большим число самок. Судя по частоте встречаемости птиц и их выживаемости, сокращение продолжительности сна в период размножения не имеет последствий для их физиологического состояния и здоровья [49].
В результате самцы, которые спят меньше, спариваются с большим число самок. Судя по частоте встречаемости птиц и их выживаемости, сокращение продолжительности сна в период размножения не имеет последствий для их физиологического состояния и здоровья [49].
Сон у млекопитающих
На Земле обитает примерно 3,5 тыс. видов млекопитающих, включая представителей плацентарных, сумчатых и однопроходных. Сон исследован более чем у 100 видов. Существование МС у однопроходных (ехидны, утконоса) не вызывает сомнений. В то же время ситуация с ПС у этих животных не понятна. Одни считают, что ПС у ехидны нет [50]. Другие придерживаются мнения, что ПС у ехидны есть [51]. Третьи, основываясь на особенностях разряда нейронов в стволе мозга, предполагают, что у ехидны есть недифференцированная стадия сна, сочетающая признаки МС и ПС плацентарных млекопитающих [52]. Интересно, что у утконоса мышечные вздрагивания и подергивания регистрируются одновременно с высоковольтной ЭЭГ. На основании этих данных была выделена стадия спокойного сна (гомологичная МС наземных млекопитающих) и стадия, которая сочетает признаки дифференцированного МС и ПС плацентарных [52, 53]. Очевидные признаки недифференцированного ПС есть и у сумчатых млекопитающих. Например, у опоссума характерные для ПС быстрые движения глаз и мышечные вздрагивания фиксируются на фоне не только активированного паттерна в ЭЭГ, но и низкоамплитудных медленных волн [54]. Таким образом, сон (в первую очередь ПС) у эволюционно «примитивных» млекопитающих (однопроходных, сумчатых) имеет признаки недифференцированности, как и у африканского страуса.
На основании этих данных была выделена стадия спокойного сна (гомологичная МС наземных млекопитающих) и стадия, которая сочетает признаки дифференцированного МС и ПС плацентарных [52, 53]. Очевидные признаки недифференцированного ПС есть и у сумчатых млекопитающих. Например, у опоссума характерные для ПС быстрые движения глаз и мышечные вздрагивания фиксируются на фоне не только активированного паттерна в ЭЭГ, но и низкоамплитудных медленных волн [54]. Таким образом, сон (в первую очередь ПС) у эволюционно «примитивных» млекопитающих (однопроходных, сумчатых) имеет признаки недифференцированности, как и у африканского страуса.
Все наземные млекопитающие во время сна неподвижны, глаза при этом закрыты, а дыхание регулярно. Медленные волны в ЭЭГ двух полушарий коры мозга развиваются одновременно. ПС обнаружен у всех наземных млекопитающих. Обычно он развивается после МС. Избирательная депривация ПС приводит к увеличению числа попыток заснуть ПС, то есть предполагается, что продолжительность ПС регулируется гомеостатическим образом. Депривация ПС сопровождается нарушением когнитивных функций, может иметь последствия для здоровья и даже приводить к смерти [55].
Депривация ПС сопровождается нарушением когнитивных функций, может иметь последствия для здоровья и даже приводить к смерти [55].
Среди млекопитающих наибольшее количество ПС регистрируется у однопроходных и сумчатых, то есть у животных с наиболее примитивно устроенным мозгом. В целом продолжительность ПС больше у незрелорождающихся видов, что говорит о его возможной роли в развитии мозга. Кроме того, существует положительная корреляция между количеством ПС у млекопитающих и степенью их защищенности во время сна [3–6]. Безусловно, нужно учитывать, что наши представления о сне животных сложились в первую очередь по результатам исследований, выполненных в лабораторных условиях. В природе животные не спят каждый день в одно и то же время, в том числе и ПС [56, 57]. Кроме того, снижение продолжительности ПС – не единственный способ сокращения времени пребывания в наиболее глубокой (судя по порогам пробуждения) стадии сна. Другой способ – изменение циркадной организации паттерна сна и смещение активности на время, когда степень опасности ниже [58].
Сон морских млекопитающих отличается от сна наземных млекопитающих по некоторым признакам, включая те, что считаются ключевыми для определения самого состояния сна. Особенности сна водных млекопитающих – прекрасный пример, в какой степени сон животных зависит от условий их обитания. Предки современных водных млекопитающих должны были «решить» несколько важных проблем, обусловленных условиями сна в воде. Во-первых, в воде невозможна длительная неподвижность. Животным нужно регулярно всплывать к поверхности воды для дыхания или поддерживать голову (ноздри) над водой. Во-вторых, в воде нет убежищ для сна, а всплытие к поверхности делает животных максимально уязвимыми для хищников (косаток, акул). Значит, во время сна животные должны постоянно поддерживать высокий уровень бдительности. В-третьих, теплопроводность воды в десятки раз выше теплопроводности воздуха.
Важнейшие особенности сна китообразных – однополушарный МС и способность спать во время движения, закрывая только один глаз [59–62].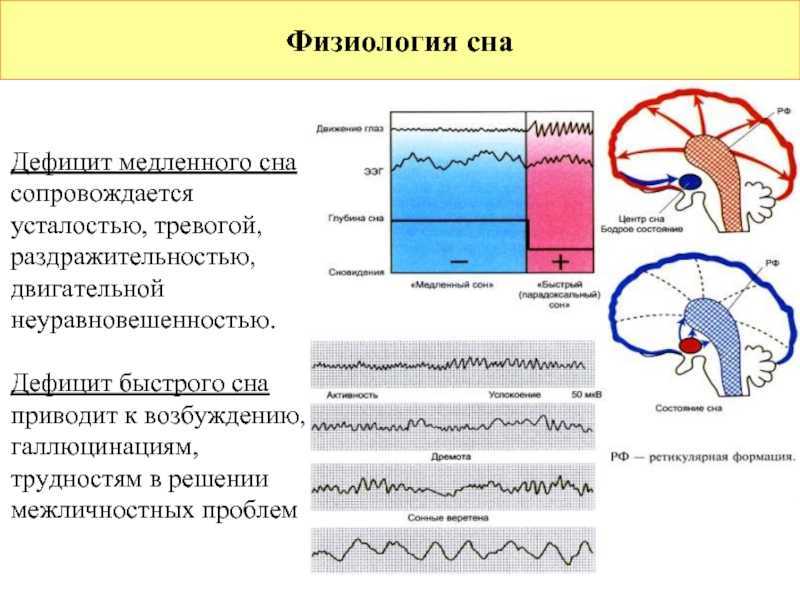 Считается, что именно однополушарный сон (будучи при этом и однополушарным бодрствованием) позволяет китообразным сочетать сон и движение, обеспечивая возможность всплытия к поверхности воды для дыхания без пробуждения. Движение также способствует мышечному термогенезу, препятствуя переохлаждению в холодной воде. Вторая функция однополушарного сна у китообразных состоит в визуальном контроле за состоянием окружающей среды с помощью одного открытого во время сна глаза. Это снижает риск неожиданного нападения хищников и помогает координировать движение с другими особями в группе (например, матерей с детенышами [60–63]).
Считается, что именно однополушарный сон (будучи при этом и однополушарным бодрствованием) позволяет китообразным сочетать сон и движение, обеспечивая возможность всплытия к поверхности воды для дыхания без пробуждения. Движение также способствует мышечному термогенезу, препятствуя переохлаждению в холодной воде. Вторая функция однополушарного сна у китообразных состоит в визуальном контроле за состоянием окружающей среды с помощью одного открытого во время сна глаза. Это снижает риск неожиданного нападения хищников и помогает координировать движение с другими особями в группе (например, матерей с детенышами [60–63]).
Причины отсутствия ПС у китообразных (во всяком случае в том в виде, в каком он регистрируется у наземных млекопитающих) не понятны [61, 62]. В то же время у китообразных есть элементы поведения (в частности, вздрагивания во время МС), которые напоминают поведенческие признаки ПС у наземных млекопитающих. Не исключено, что ПС у дельфинов и китов может протекать в модифицированной форме (например, в виде коротких эпизодов длительностью всего в несколько секунд), что затрудняет его идентификацию. Даже в этом случае продолжительность ПС у китообразных была бы значительно меньше, чем у наземных млекопитающих и птиц. Считается, что ПС выполняет витальную (жизнеподдерживающую) функцию. В чем бы эта функция ни состояла, она оказывается менее важной для китообразных по сравнению с теми рисками, которые возникают при развитии длинных эпизодов ПС.
Даже в этом случае продолжительность ПС у китообразных была бы значительно меньше, чем у наземных млекопитающих и птиц. Считается, что ПС выполняет витальную (жизнеподдерживающую) функцию. В чем бы эта функция ни состояла, она оказывается менее важной для китообразных по сравнению с теми рисками, которые возникают при развитии длинных эпизодов ПС.
Ластоногие – группа полуводных животных (отряд хищных), которые могут спать и на суше, и в воде. Сон ластоногих сочетает признаки сна наземных млекопитающих и китообразных. Один из типичных представителей ластоногих, в частности семейства ушастых тюленей, и одна из моделей для изучения механизмов сна у млекопитающих – северный морской котик. Морские котики обитают преимущественно в умеренной и субтропической климатических зонах. Во время летнего периода они проводят большую часть времени на суше, где размножаются и линяют. В зимний период котики мигрируют на расстояние до 2000 км и не выходят на сушу до десяти месяцев. В воде они спят на поверхности в характерной позе, поддерживая голову над водой и совершая гребковые движения одним передним ластом (рис.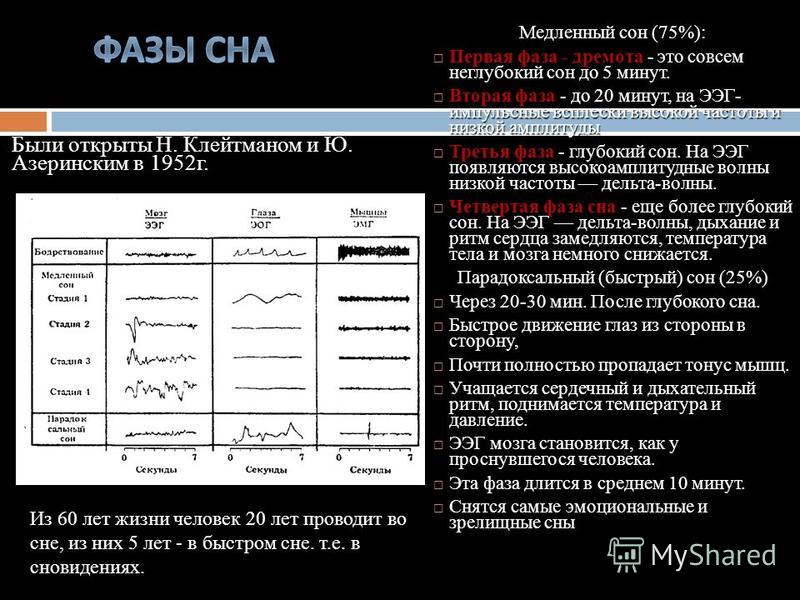 2). Такая поза позволяет снизить потери тепла (через находящиеся в воздухе ласты) и обеспечивает регулярное дыхание. Во время сна в воде котики, как и китообразные, закрывают на продолжительное время только один глаз, тогда как второй направлен в сторону воды, что повышает вероятность обнаружения хищников [63–65].
2). Такая поза позволяет снизить потери тепла (через находящиеся в воздухе ласты) и обеспечивает регулярное дыхание. Во время сна в воде котики, как и китообразные, закрывают на продолжительное время только один глаз, тогда как второй направлен в сторону воды, что повышает вероятность обнаружения хищников [63–65].
Наши исследования показали, что переход от наземного к водному образу жизни сопровождается у котиков «переключением» с «типичного наземного» (билатерально-симметричный МС, неподвижность, ПС) на характерный для китообразных «водный» тип сна (однополушарный сон, сохранение двигательной активности, прерывистый тип дыхания, отсутствие ПС) (рис. 3 и 4) [65, 66]). Способность котиков естественным образом фактически полностью исключить ПС из жизни на как минимум одну-две недели без каких бы то ни было очевидных последствий представляет большой интерес для понимания функции и биологической роли ПС. Исчезновение ПС при переходе от сна на суше ко сну в воде говорит о важности экологических факторов в регуляции ПС у этого вида. При этом исчезновение ПС происходит в условиях, когда животным необходимо поддерживать высокий уровень двигательной активности и бдительности. Находясь в воде две недели, морские котики накапливают огромный дефицит ПС в количестве 15–20 часов. После возвращения на сушу у них не наблюдается значимого превышения ПС над контрольными значениями на суше. Полученные данные ставят под сомнение представления о гомеостатической регуляции ПС. В то же время они хорошо согласуются с ранее высказанной гипотезой, что ПС необходим для смягчения возможных негативных последствий глубокого билатерально-симметричного МС (например, снижения температуры мозга [67]). Это могло бы объяснить не только сокращение ПС в воде у котиков (параллельно с исчезновением билатерально-симметричного МС), но и отсутствие ПС у дельфинов (у которых высокоамплитудный билатерально-симметричный МС не регистрируется). Особенности сна морских млекопитающих необходимо каким-то образом согласовать с представлением о роли ПС в поддержании когнитивных функций, или следует признать, что эта стадия сна выполняет различные функции у разных видов и групп.
При этом исчезновение ПС происходит в условиях, когда животным необходимо поддерживать высокий уровень двигательной активности и бдительности. Находясь в воде две недели, морские котики накапливают огромный дефицит ПС в количестве 15–20 часов. После возвращения на сушу у них не наблюдается значимого превышения ПС над контрольными значениями на суше. Полученные данные ставят под сомнение представления о гомеостатической регуляции ПС. В то же время они хорошо согласуются с ранее высказанной гипотезой, что ПС необходим для смягчения возможных негативных последствий глубокого билатерально-симметричного МС (например, снижения температуры мозга [67]). Это могло бы объяснить не только сокращение ПС в воде у котиков (параллельно с исчезновением билатерально-симметричного МС), но и отсутствие ПС у дельфинов (у которых высокоамплитудный билатерально-симметричный МС не регистрируется). Особенности сна морских млекопитающих необходимо каким-то образом согласовать с представлением о роли ПС в поддержании когнитивных функций, или следует признать, что эта стадия сна выполняет различные функции у разных видов и групп.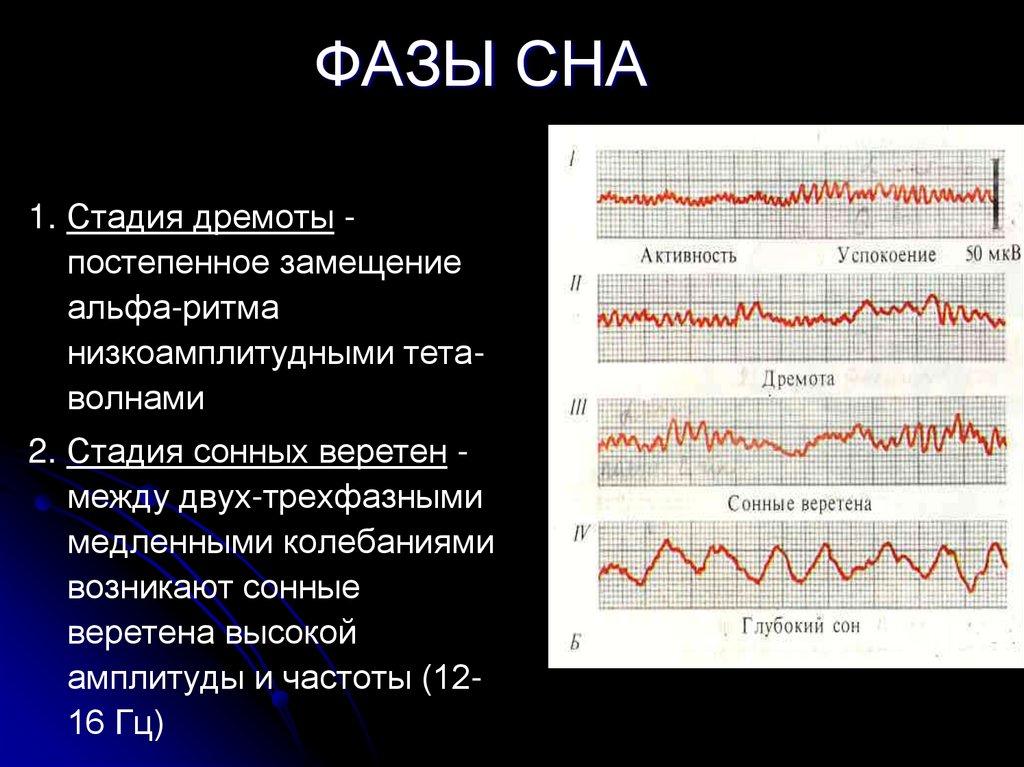
Отсутствие/резкое сокращение ПС у дельфинов и морских котиков вовсе не означает, что ПС несовместим с водным образом жизни, а однополушарный МС на фоне постоянной двигательной активности – единственная возможность для млекопитающих спать в водной среде. Существует и другой паттерн (стратегия) сна, который сформировался в эволюции у настоящих тюленей. Большинство настоящих тюленей, а также моржи обитают в холодных водах субполярных и полярных районов вдоль кромки льдов или в замерзающих озерах и внутренних морях в условиях ограниченного доступа к открытой воде. Задерживая дыхание, тюлени могут спать под водой на глубине [68]. Подобный паттерн сна не требует поддержания двигательной активности. Поэтому МС у тюленей билатерально-симметричный, как у неподвижно спящих наземных млекопитающих. Сохранение тепла у тюленей достигается посредством толстого подкожного жирового слоя. Эффективная защита от переохлаждения не препятствует развитию ПС. При этом они должны пробуждаться для движения при всплытии к поверхности воды [63].
Таким образом, имеющиеся данные об особенностях сна водных млекопитающих указывают на важность экологических факторов в формировании паттерна сна. Необходимость всплывать к поверхности воды для дыхания, постоянно контролировать окружающую среду и осуществлять эффективную терморегуляцию – основные факторы, определившие формирование структуры сна у современных водных млекопитающих [61–66]. Однополушарный МС – одна из главных особенностей сна китообразных и ушастых тюленей сформировалась в эволюции этих групп независимо в условиях действия одних и тех же экологических факторов. Чрезвычайно интересно, что однополушарный МС у морских млекопитающих и птиц фенотипически похож, несмотря на различия в анатомической организации их переднего мозга. Не исключено, что в будущем такая форма сна обнаружится и у других представителей животного мира, для которых условия сна требуют поддержания высокого уровня бдительности или сохранения двигательной активности во время сна.
Заключение
Наиболее интересные и значимые результаты в нейробиологии сна (сомнологии) последних 20 лет были получены при изучении сна у простых организмов, а также при исследовании сна животных в природе. Оказалось, что сон – универсальное состояние всех животных, от простых форм с децентрализованной нервной системой до высших млекопитающих со сложно устроенным головным мозгом. У всех животных состояние сна характеризуется удивительно похожими поведенческими, нейронными, молекулярными и физиологическими признаками. В то же время две дифференцированные стадии сна – МС и ПС (или быстрый сон) существуют только у млекопитающих и птиц. Поэтому сравнительные исследования сна у млекопитающих и птиц особенно важны для осмысления эволюции сна и механизмов его регуляции. Для понимания природы и функции (функций) сна и его стадий важно изучать сон животных в условиях, предельно приближенных к естественным, и в дикой природе. При этом необходимо учитывать влияние на поведение и сон максимально возможного числа внешних факторов, динамику их изменений и особенности биологического цикла данного вида животных.
Оказалось, что сон – универсальное состояние всех животных, от простых форм с децентрализованной нервной системой до высших млекопитающих со сложно устроенным головным мозгом. У всех животных состояние сна характеризуется удивительно похожими поведенческими, нейронными, молекулярными и физиологическими признаками. В то же время две дифференцированные стадии сна – МС и ПС (или быстрый сон) существуют только у млекопитающих и птиц. Поэтому сравнительные исследования сна у млекопитающих и птиц особенно важны для осмысления эволюции сна и механизмов его регуляции. Для понимания природы и функции (функций) сна и его стадий важно изучать сон животных в условиях, предельно приближенных к естественным, и в дикой природе. При этом необходимо учитывать влияние на поведение и сон максимально возможного числа внешних факторов, динамику их изменений и особенности биологического цикла данного вида животных.
Исследования сна птиц поддержаны грантом Российского фонда фундаментальных исследований (18-04-01252).
Исследования сна морских млекопитающих поддержаны грантами Национального научного фонда США, Национального института здоровья США, Российского фонда фундаментальных исследований и ООО «Утришский дельфинарий» (Россия).
Парадоксальный сон: стадия сна или расстройство?
Специалисты по сну должны действительно любить слово «парадокс», потому что они используют его для описания четырех уникальных концепций сна, которые все очень разные: парадоксальный сон, парадоксальное лишение сна, парадоксальная бессонница и парадоксальное намерение.
Это может показаться запутанным, но не бойтесь. Мы познакомим вас со всеми четырьмя из них, чтобы помочь вам лучше понять, что эти концепции означают для вас, и как получить Zzz, который вам нужен.
Что такое парадоксальный сон?
Парадоксальный сон широко известен как сон с быстрыми движениями глаз (БДГ), но он имеет много других названий, включая активированный сон, десинхронизированный сон, сон во сне и D-сон.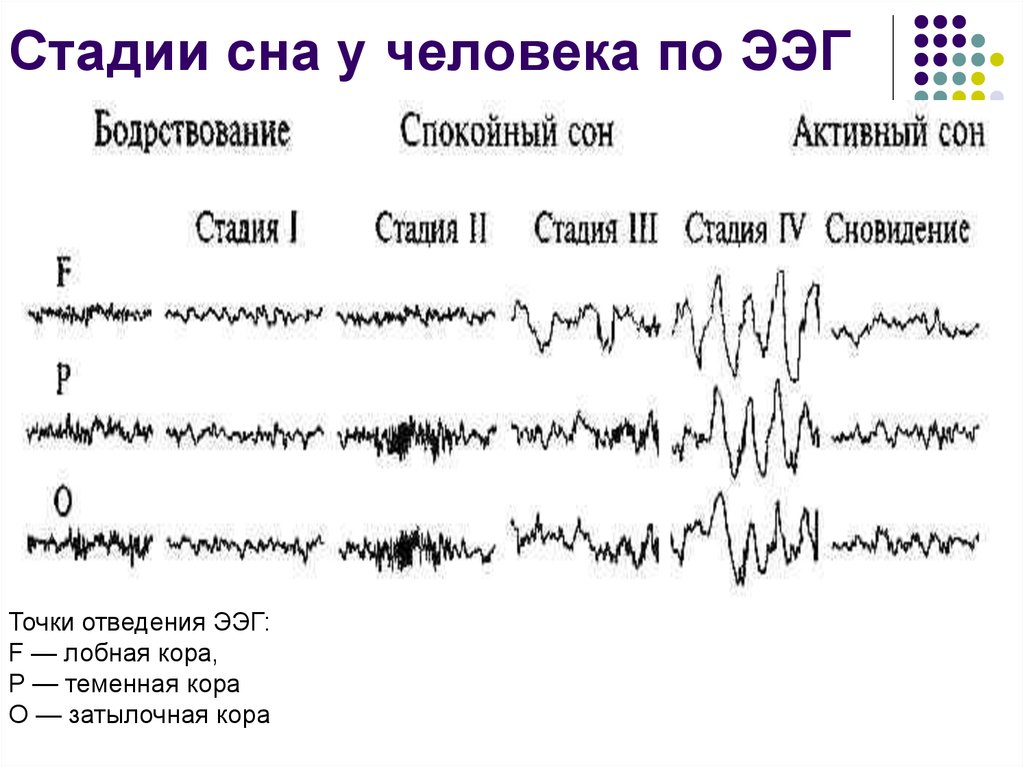 Если эти термины слишком просты для вас, вы также можете назвать это ромбэнцефальным сном.
Если эти термины слишком просты для вас, вы также можете назвать это ромбэнцефальным сном.
Парадоксальный сон — это , а не расстройство сна. Это фаза сна, когда происходит большая часть ваших сновидений, и это также очень важное время для восстановления вашего мозга и мышц.
Поделиться на PinterestIbai Acevedo/Stocksy United — Изображение для иллюстративных целей.Согласно Merriam-Webster, парадокс — это «утверждение, которое кажется противоречащим или противоречащим здравому смыслу, но, возможно, является истинным».
Некоторыми простыми примерами этого являются высказывания «Меньше значит больше» и «Если я что-то и знаю, так это то, что я ничего не знаю».
Во время парадоксального (он же БДГ-сна) все становится тривиальным. Ваш сердечный ритм увеличивается, и ваше тело перестает регулировать температуру. Ваша мозговая деятельность сильна (как если бы вы бодрствовали), поэтому у вас могут быть яркие сны, но ваше тело по существу парализовано.
В общем, ваш мозг устраивает большую старую вечеринку, на которую ваше тело не приглашено! И в этом парадокс. 🥳
🥳
Спите на этом
Несмотря на то, что мы тратим на это треть своей жизни, феномен сна по-прежнему окружает много загадок.
Большинство экспертов считают, что наш организм автоматически отключает произвольную мышечную активность, чтобы мы не причиняли вреда себе или другим во сне. Но наша непроизвольная мышечная деятельность продолжается, потому что, ну, дыхание.
Парадоксальное недосыпание — это когда вы недостаточно времени проводите в стадии парадоксального сна. Таким образом, вы можете заснуть и спать, но вы застряли в стадии легкого сна.
Поскольку восстановление мышц, выведение отходов, построение мозга и энергетическая подзарядка — среди прочего — происходят на более глубоких стадиях сна, пребывание в режиме поверхностного сна может иметь серьезные долгосрочные последствия для здоровья.
Некоторые симптомы парадоксальной депривации сна:
- hormonal issues
- increased appetite and weight gain
- excessive tiredness
- lack of concentration and poor memory
- difficulty regulating body temperature
- lower immune system function
- puffy eyes and droopy eyelids
- anxiety and depression
Для постановки диагноза парадоксального недосыпания может потребоваться остаться на ночь для проведения анализов в лаборатории сна или носить устройство для мониторинга сна в течение нескольких недель.
Устройства для мониторинга сна могут дать вам точную оценку общего времени сна, но они не так точны в определении различных стадий сна, включая быстрый сон.
Методы лечения различаются, поскольку они зависят от первопричины. Некоторые распространенные методы лечения:
- Лекарства, отпускаемые по рецепту. Снотворные, такие как золпидем и суворексант, следует принимать с осторожностью, а не в качестве долгосрочного решения. И они не решат основную проблему.
- Терапия контроля стимулов. Ограничение деятельности, которую вы выполняете в спальне, может помочь укрепить связь между спальней и сном.
- Релаксация и терапия с биологической обратной связью. Доктора используют датчики на вашей коже для записи мышечного напряжения и мозговых ритмов, чтобы помочь вам манипулировать своим мозгом, чтобы он заснул. Эти методы не очень практичны и обычно используются в исследовательских целях.
- Терапия ограничения сна.
 Это включает в себя ограничение времени, которое вы проводите в своей постели.
Это включает в себя ограничение времени, которое вы проводите в своей постели. - Ароматерапия. Вы используете растительные масла, чтобы теоретически активировать мозговые сенсоры, вызывающие сон.
 Это включает в себя ограничение времени, которое вы проводите в своей постели.
Это включает в себя ограничение времени, которое вы проводите в своей постели.Парадоксальная бессонница — это отдельная тема. Это редкое расстройство сна, при котором вы чувствуете , что не спите, но не проявляете физических признаков недосыпания. Это также известно как искажение состояния сна.
Это может быть разочаровывающим и одиноким опытом, потому что окружающие могут подумать, что вы лжете или пытаетесь привлечь внимание. Если вы пройдете обследование, результаты покажут, что вы действительно спите, поэтому поставить диагноз может быть сложно.
Парадоксальная бессонница может привести к:
- ненужному (и неэффективному) самолечению
- низкой самооценке из-за ощущения, что тебе никто не верит
- напряженности в отношениях
- тревоге и депрессии , парадоксальная бессонница лучше всего лечится многими из тех же методов, которые используются для бессонницы, такими как:
- успокоение пациента
- когнитивно-поведенческая реструктуризация (включая парадоксальное намерение)
- обучение гигиене сна
- методы медитации и релаксации
- лекарства (краткосрочное решение)
парадоксальное намерение результат.
Таким образом, вместо того, чтобы лежать в постели, заставляя себя заснуть (что никогда не срабатывает, кстати), вы заставите себя бодрствовать.Это может уменьшить давление и тревогу, связанные с ощущением себя таким, как вы есть , чтобы заснуть и, в некотором смысле, заставить ваш мозг чувствовать себя сонным. Если вы когда-нибудь пробовали , а не , чтобы заснуть во время учебы, вождения или просмотра фильма, то вы можете увидеть, как работает этот метод!
Парадоксальный сон (также называемый БДГ-сном) характеризуется сверхактивным мозгом и почти парализованным телом, и в нем также происходят многие восстановительные процессы.
Слово «парадокс» также появляется в других понятиях, связанных со сном, в том числе:
- Парадоксальная депривация сна: недостаточное количество сна на стадии парадоксального (БДГ) сна
- Парадоксальная бессонница: ощущение, что вы не спите, когда вы на самом деле
- Парадоксальная терапия: когнитивно-поведенческая терапия: 9006 метод лечения бессонницы, предполагающий преднамеренные попытки бодрствовать вместо попытки заснуть
Парадоксальный сон не является расстройством сна.
Но если у вас когда-нибудь возникнут проблемы со сном, в котором вы нуждаетесь, обратитесь к медицинскому работнику, чтобы обсудить способы улучшения вашего сна. Хороший сон абсолютно необходим для вашей лучшей и здоровой жизни.Быстрый сон: биологический и психологический парадокс
1. Siegel JM. Сон рассматривается как состояние адаптивной бездеятельности. Нат Рев Нейроски. 2009; 10: 747–753. [Бесплатная статья PMC] [PubMed] [Google Scholar]
2. Мэтьюз П., Уайт К. Прерывистый газообмен у насекомых: все в их головах? Американский натуралист. 2011; 177:130–134. [PubMed] [Google Scholar]
3. Швейцер П.К. В: Лекарства, нарушающие сон и бодрствование в принципах и практике медицины сна. Крайгер М.Х., Рот Т., Демент В.К., редакторы. Амстердам: Эльзевир Сондерс; 2011. стр. 542–560. [Академия Google]
4. Rasch B, Pommer J, Diekelmann S, Born J. Фармакологическое подавление быстрого сна парадоксально улучшает, а не ухудшает память навыков. Нат Нейроски.
2009; 12: 396–397. [PubMed] [Google Scholar]5. Lavie P, Pratt H, Scharf B, Peled R, Brown J. Локализованное поражение моста: почти полное отсутствие быстрого сна. Неврология. 1984; 34: 118–120. [PubMed] [Google Scholar]
6. Rechtschaffen A, Bergmann BM. Лишение сна у крыс: обновление статьи 1989 г. Спать. 2002; 25:18–24. [PubMed] [Академия Google]
7. Сигел Дж.М. Все ли животные спят? Тренды Нейроси. 2008; 31: 208–213. [Бесплатная статья PMC] [PubMed] [Google Scholar]
8. Roth TC, Lesku JA, Amlaner CJ, Lima SL. Филогенетический анализ коррелятов сна у птиц. Дж. Сон Рез. 2006; 15: 395–402. [PubMed] [Google Scholar]
9. Holty JE, Guilleminault C. Синдром брадиаритмии, связанный с REM. Обзоры медицины сна. 2010 г. В печати, исправленное доказательство. [PubMed] [Google Scholar]
10. Luppi P-H, Clément O, Sapin E, Gervasoni D, Peyron C, Léger L, et al. Нейронная сеть, ответственная за парадоксальный сон и ее дисфункции, вызывающие нарколепсию и расстройство поведения с быстрыми движениями глаз (БДГ).
Sleep Med Rev. 2010 В печати, исправленное доказательство. [PubMed] [Академия Google]11. Маллик Б.Н., Сингх А. Потеря быстрого сна повышает возбудимость мозга: роль норадреналина и его механизм действия. Sleep Med Rev. 2011 [PubMed] [Google Scholar]
12. Палагини Л., Розенлихт Н. Сон, сновидения и психическое здоровье: обзор исторических и нейробиологических перспектив. Sleep Med Rev. 2010 В печати, исправленное доказательство. [PubMed] [Google Scholar]
13. Plazzi G, Moghadam KK, Maggi LS, Donadio V, Vetrugno R, Liguori R, et al. Вегетативные расстройства при нарколепсии. Sleep Med Rev. 2010 [PubMed] [Google Scholar]
14. Синтон CM. Орексин/гипокретин играет роль в реакции на физиологическое нарушение равновесия. Sleep Med Rev. 2011 [PubMed] [Google Scholar]
15. Асеринский Э., Клейтман Н. Регулярно возникающие периоды моторики глаз и сопутствующие явления во время сна. Наука. 1953; 118: 273–274. [PubMed] [Google Scholar]
16. Siegel JM.
Ключи к функциям сна млекопитающих. Природа. 2005; 437:1264–1271. [Бесплатная статья PMC] [PubMed] [Google Scholar]17. Солмс М. Нейропсихология сновидений: клинико-анатомическое исследование. Махва, Нью-Джерси: Lawrence Erlbaum Associates; 1997. [Google Scholar]
18. Pagel JF. Немечтатели. Сон Мед. 2003; 4: 235–241. [PubMed] [Google Scholar]
19. Фоулкс Д. Детское сновидение и развитие сознания. Кембридж, Массачусетс: Издательство Гарвардского университета; 2002. [Google Scholar]
20. Siegel JM, Manger PR, Nienhuis R, Fahringer HM, Shalita T, Pettigrew JD. Сон в утконосе. Неврологи. 1999; 91: 391–400. [PMC free article] [PubMed] [Google Scholar]
21. Асеринский Е. Связь плотности быстрых движений глаз с предшествующим накоплением сна и бодрствования. Психофизиол. 1973;10:545–558. [PubMed] [Google Scholar]
22. Leclair-Visonneau L, Oudiette D, Gaymard B, Leu-Semenescu S, Arnulf I. Сканируют ли глаза изображения сновидений во время быстрого сна? Данные модели расстройства поведения во сне с быстрыми движениями глаз.
Мозг. 2010; 133:1737–1746. [PubMed] [Google Scholar]23. Hobson JA, McCarley RW. Мозг как генератор состояния сна: гипотеза активации-синтеза процесса сна. Am J психиатрии. 1977; 134: 1335–1348. [PubMed] [Академия Google]
24. Задра А., Домхофф Г.В. В: Содержание сновидения: количественные данные в «Принципах и практике медицины сна». Крайгер М.Х., Рот Т., Демент В.К., редакторы. Сент-Луис, Миссури: Эльзевир Сондерс; 2011. стр. 585–594. [Google Scholar]
25. Гузман-Марин Р., Сунцова Н., Башир Т., Ниенхуйс Р., Шимусяк Р., МакГинти Д. Депривация сна с быстрым движением глаз способствует снижению нейрогенеза в зубчатой извилине гиппокампа взрослой крысы. Спать. 2008; 31: 167–175. [Бесплатная статья PMC] [PubMed] [Google Scholar]
26. Сигель Дж. М., Ниенхуис Р., Томашевски К. С. Признаки быстрого сна ростральные по отношению к хроническим разрезам в понтомедуллярном соединении. Неврологи. лат. 1984; 45: 241–246. [PMC бесплатная статья] [PubMed] [Google Scholar]
27.
Siegel JM, Tomaszewski KS, Nienhuis R. Поведенческие состояния при хроническом медуллярном и среднем мостовом состоянии кошки. электроэнцеф. клин. Нейрофизиол. 1986; 63: 274–288. [Бесплатная статья PMC] [PubMed] [Google Scholar]28. Siegel JM. Нейробиология сна. Семинары по неврологии. 2009 г.;29:277–296. [Бесплатная статья PMC] [PubMed] [Google Scholar]
29. Siegel JM, Manger P, Nienhuis R, Fahringer HM, Pettigrew J. Ехидна Tachyglossus aculeatus сочетает в себе фазы REM и nonREM в одном состоянии сна: значение для эволюции сна. Дж. Нейроски. 1996;16:3500–3506. [Бесплатная статья PMC] [PubMed] [Google Scholar]
30. George R, Haslett WL, Jenden DJ. Холинергический механизм в ретикулярной формации ствола мозга: индукция парадоксального сна. Междунар. Дж. Нейрофармакол. 1964;3:541–552. [PubMed] [Google Scholar]
31. Широмани П., Сигел Дж.М., Томашевски К.С., МакГинти Д.Дж. Изменения артериального давления и фазы быстрого сна после микроинфузии карбахола понтина.
Эксп. Нейрол. 1986; 91: 285–292. [Бесплатная статья PMC] [PubMed] [Google Scholar]32. Lai YY, Siegel JM. Изменения сердечно-сосудистого и мышечного тонуса, вызванные микроинъекцией холинергических и глутаматергических агонистов в дорсолатеральный мост и медиальный мозг. Мозг Res. 1990; 514: 27–36. [Бесплатная статья PMC] [PubMed] [Google Scholar]
33. Широмани П.Дж., Лай Ю.Ю., Сигел Дж.М. Нисходящие проекции от дорсолатеральной покрышки моста к парамедианному ретикулярному ядру каудального отдела продолговатого мозга у кошки. Мозг Res. 1990; 517: 224–228. [Бесплатная статья PMC] [PubMed] [Google Scholar]
34. Lai YY, Siegel JM. Понтомедуллярные рецепторы глутамата, опосредующие локомоцию и подавление мышечного тонуса. Дж. Нейроски. 1991; 11: 2931–2937. [Бесплатная статья PMC] [PubMed] [Google Scholar]
35. Kodama T, Lai YY, Siegel JM. Усиление высвобождения ацетилхолина во время быстрого сна в каудомедиальном отделе продолговатого мозга, измеренное с помощью микродиализа in vivo.
Мозг Res. 1992;580:348–350. [Бесплатная статья PMC] [PubMed] [Google Scholar]36. Lai YY, Clements J, Siegel J. Глутаматергические и холинергические проекции на зону торможения моста, идентифицированную с помощью ретроградного транспорта пероксидазы хрена и иммуногистохимии. Дж. Комп. Нейрол. 1993; 336: 321–330. [Бесплатная статья PMC] [PubMed] [Google Scholar]
37. Kodama T, Lai YY, Siegel JM. Повышенное высвобождение глутамата во время быстрого сна в ростромедиальном мозговом веществе, измеренное с помощью микродиализа in vivo. Мозг Res. 1998;780:178–181. [Бесплатная статья PMC] [PubMed] [Google Scholar]
38. Reid MS, Nishino S, Siegel JM, Dement WC, Mignot E, et al. Нейрофармакологическая характеристика базальной холинергической катаплексии переднего мозга у нарколептических собак. Эксп. Нейрол. 1998; 151:89–104. [Бесплатная статья PMC] [PubMed] [Google Scholar]
39. Hajnik T, Lai YY, Siegel JM. Области, связанные с атонией, в мосту и мозговом веществе грызунов.
J Нейрофизиол. 2000; 84: 1942–1948. [Бесплатная статья PMC] [PubMed] [Google Scholar]40. Милейковский Б.Ю., Киященко Л.И., Кодама Т., Лай Ю.Ю., Сигель Дж.М. Активация мостовых и медуллярных двигательных тормозных областей уменьшает разряд в нейронах, расположенных в голубом пятне и анатомическом эквиваленте локомоторной области среднего мозга. Дж. Нейроски. 2000;20:8551–8558. [Бесплатная статья PMC] [PubMed] [Google Scholar]
41. Swanson LW. Карты мозга: структура мозга крысы. Elsevier Science: Амстердам; 1998. [Google Scholar]
42. Nitz D, Siegel JM. Высвобождение ГАМК в дорсальном ядре шва: роль в контроле быстрого сна. амер. Дж. Физиол. 1997;273:R451–R455. [Бесплатная статья PMC] [PubMed] [Google Scholar]
43. Nitz D, Siegel JM. Высвобождение ГАМК в голубом пятне кошки в зависимости от состояния сна/бодрствования. Неврологи. 1997; 78: 795–801. [Бесплатная статья PMC] [PubMed] [Google Scholar]
44. Siegel JM, Rogawski MA. Функция быстрого сна: регуляция чувствительности норадренергических рецепторов.
Мозг Res. 1988; 13:213–233. [Бесплатная статья PMC] [PubMed] [Google Scholar]45. Маллик Б.Н., Каур С., Саксена Р.Н. Взаимодействия между холинергическими и ГАМКергическими нейротрансмиттерами в голубом пятне и вокруг него для индукции и поддержания сна с быстрыми движениями глаз у крыс. Неврологи. 2001; 104: 467–485. [PubMed] [Академия Google]
46. Ву М.Ф., Гуляни С.А., Яу Э., Миньо Э., Фан Б., Сигель Дж.М. Нейроны голубого пятна: прекращение активности при катаплексии. Неврологи. 1999; 91: 1389–1399. [Бесплатная статья PMC] [PubMed] [Google Scholar]
47. Siegel JM, et al. Нейронная активность при нарколепсии: идентификация клеток, связанных с катаплексией, в медиальном мозговом веществе. Наука. 1991; 252:1315–1318. [Бесплатная статья PMC] [PubMed] [Google Scholar]
48. Rechtschaffen A, Wolpert EA, Dement WC, Mitchel SA, Fisher C. Ночной сон нарколептиков. электроэнцеф. клин. Нейрофизиол. 1963;15:599–609. [PubMed] [Google Scholar]
49. Peyron C, Faraco J, Rogers W, Ripley B, Overeem S, Charnay Y, et al.
Мутация в случае ранней нарколепсии и общее отсутствие гипокретиновых пептидов в нарколептическом мозге человека. Нац. Мед. 2000; 6: 991–997. [PubMed] [Google Scholar]50. Thannickal TC, Moore RY, Nienhuis R, Ramanathan L, Gulyani S, Aldrich M, et al. Снижение числа гипокретиновых нейронов при нарколепсии человека. Нейрон. 2000; 27: 469–474. [Бесплатная статья PMC] [PubMed] [Google Scholar]
51. Милейковский Б.Я., Киященко Л.И., Зигель Ю.М. Поведенческие корреляты активности идентифицированных гипокретин/орексиновых нейронов. Нейрон. 2005; 46: 787–798. [Бесплатная статья PMC] [PubMed] [Google Scholar]
52. Ву М.Ф., Джон Дж., Бомер Л.Н., Яу Д., Нгуен Г.Б., Сигел Дж.М. Активность клеток дорсального шва в цикле сон-бодрствование и во время катаплексии у нарколептических собак. Дж. Физиол. 2004; 554: 202–215. [Бесплатная статья PMC] [PubMed] [Google Scholar]
53. Siegel JM, Nienhuis R, Fahringer HM, Chiu C, Dement WC, Mignot E, Lufkin R. Активность медиальных мезопонтинных единиц во время катаплексии и состояний сна-бодрствования у нарколептическая собака.
Дж. Нейроски. 1992;12:1640–1646. [Бесплатная статья PMC] [PubMed] [Google Scholar]54. John J, Wu MF, Boehmer LN, Siegel JM. Катаплексия-активные нейроны в заднем гипоталамусе: влияние гистамина на поведение во сне и бодрствовании. Нейрон. 2004; 42: 619–634. [Бесплатная статья PMC] [PubMed] [Google Scholar]
55. Kayaba Y, Nakamura A, Kasuya Y, Ohuchi T, Yanagisawa M, Komuro I, et al. Ослабленная защитная реакция и низкое базальное кровяное давление у мышей с нокаутом орексина. Являюсь. J Физиол Регул. интегр. Комп Физиол. 2003; 285: Р581–Р593. [PubMed] [Google Scholar]
56. Siegel JM. Гипокретин (орексин): роль в нормальном поведении и невропатологии. Ежегодный выпуск журнала Psychol. 2004; 55: 125–148. [Бесплатная статья PMC] [PubMed] [Google Scholar]
57. Willie JT, Chemelli RM, Sinton CM, Tokita S, Williams SC, Kisanuki YY, et al. Различные синдромы нарколепсии у мышей с нулевым рецептором орексина-2 и с нулевым орексином: молекулярно-генетическое рассечение процессов регуляции не-БДГ и БДГ-сна.
 Таким образом, вместо того, чтобы лежать в постели, заставляя себя заснуть (что никогда не срабатывает, кстати), вы заставите себя бодрствовать.
Таким образом, вместо того, чтобы лежать в постели, заставляя себя заснуть (что никогда не срабатывает, кстати), вы заставите себя бодрствовать. Но если у вас когда-нибудь возникнут проблемы со сном, в котором вы нуждаетесь, обратитесь к медицинскому работнику, чтобы обсудить способы улучшения вашего сна. Хороший сон абсолютно необходим для вашей лучшей и здоровой жизни.
Но если у вас когда-нибудь возникнут проблемы со сном, в котором вы нуждаетесь, обратитесь к медицинскому работнику, чтобы обсудить способы улучшения вашего сна. Хороший сон абсолютно необходим для вашей лучшей и здоровой жизни. 2009; 12: 396–397. [PubMed] [Google Scholar]
2009; 12: 396–397. [PubMed] [Google Scholar] Sleep Med Rev. 2010 В печати, исправленное доказательство. [PubMed] [Академия Google]
Sleep Med Rev. 2010 В печати, исправленное доказательство. [PubMed] [Академия Google] Ключи к функциям сна млекопитающих. Природа. 2005; 437:1264–1271. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Ключи к функциям сна млекопитающих. Природа. 2005; 437:1264–1271. [Бесплатная статья PMC] [PubMed] [Google Scholar] Мозг. 2010; 133:1737–1746. [PubMed] [Google Scholar]
Мозг. 2010; 133:1737–1746. [PubMed] [Google Scholar] Siegel JM, Tomaszewski KS, Nienhuis R. Поведенческие состояния при хроническом медуллярном и среднем мостовом состоянии кошки. электроэнцеф. клин. Нейрофизиол. 1986; 63: 274–288. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Siegel JM, Tomaszewski KS, Nienhuis R. Поведенческие состояния при хроническом медуллярном и среднем мостовом состоянии кошки. электроэнцеф. клин. Нейрофизиол. 1986; 63: 274–288. [Бесплатная статья PMC] [PubMed] [Google Scholar] Эксп. Нейрол. 1986; 91: 285–292. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Эксп. Нейрол. 1986; 91: 285–292. [Бесплатная статья PMC] [PubMed] [Google Scholar] Мозг Res. 1992;580:348–350. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Мозг Res. 1992;580:348–350. [Бесплатная статья PMC] [PubMed] [Google Scholar] J Нейрофизиол. 2000; 84: 1942–1948. [Бесплатная статья PMC] [PubMed] [Google Scholar]
J Нейрофизиол. 2000; 84: 1942–1948. [Бесплатная статья PMC] [PubMed] [Google Scholar]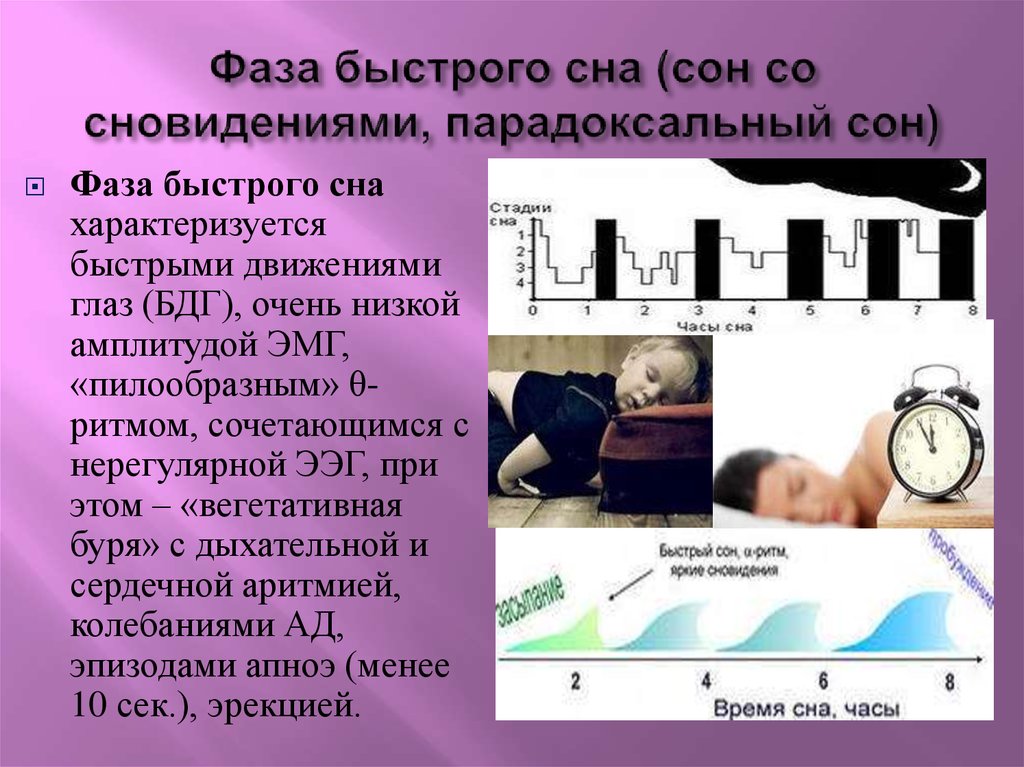 Мозг Res. 1988; 13:213–233. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Мозг Res. 1988; 13:213–233. [Бесплатная статья PMC] [PubMed] [Google Scholar] Мутация в случае ранней нарколепсии и общее отсутствие гипокретиновых пептидов в нарколептическом мозге человека. Нац. Мед. 2000; 6: 991–997. [PubMed] [Google Scholar]
Мутация в случае ранней нарколепсии и общее отсутствие гипокретиновых пептидов в нарколептическом мозге человека. Нац. Мед. 2000; 6: 991–997. [PubMed] [Google Scholar]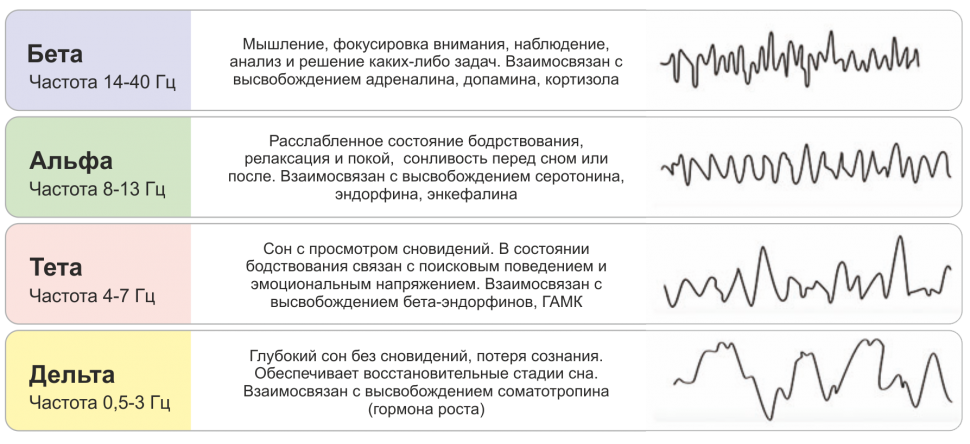 Дж. Нейроски. 1992;12:1640–1646. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Дж. Нейроски. 1992;12:1640–1646. [Бесплатная статья PMC] [PubMed] [Google Scholar]