физиология активности — это… Что такое физиология активности?
— концепция, трактующая поведение организма как активное отношение к среде, определяемое потребной организму моделью будущего — искомого результата. Как специальное направление исследований развита Н. А. Бернштейном, разработавшим ее на основе изучения движений.
В ней активность рассматривается как существенное свойство организма животного, определяющее его поведение. Физиология активности была новым шагом в развитии нейрофизиологии, психологии и даже биологии: был совершен переход от рассмотрения организма как реактивной системы к рассмотрению его как активной системы. Идеи физиологии активности оказали влияние на развитие ряда наук, в том числе психологию.
Активность проявляется, когда запрограммированное организмом движение к определенной цели требует преодоления сопротивления среды. На это преодоление организм отпускает энергию, пока не восторжествует над средой или не погибнет. Организм движется негэнтропически, добиваясь понижения уровня энтропии в самом себе и оплачивая это ценой метаболического возрастания энтропии в своем окружении за счет разрушения веществ — участников энергетического метаболизма.
Принцип физиологической и биологической активности альтернативен концепциям, основанным на представлении о дуге рефлекторной для условного и безусловного рефлексов, поскольку стимул, будучи бесспорной формой появления рефлекса, ничего не говорит о значении этого рефлекса — о том, почему на этот стимул организм отвечает именно так, а не иначе. А физиология активности стремится ответить на вопрос, для чего совершается реакция организма, какой потребности она соответствует.
Жизнедеятельность организма — не уравновешение со средой, но активное преодоление среды, определяемое моделью будущего потребного. Активность строится соответственно вероятностному прогнозированию развития событий в среде и положению в ней организма.
Модель будущего потребного строится на базе информации о текущей ситуации и прежнем опыте, но не носит жесткого характера, ибо организм постоянно стоит перед надобностью вероятностного прогноза и выбора самых эффективных путей достижения цели или решения двигательной задачи.
Словарь практического психолога. — М.: АСТ, Харвест. С. Ю. Головин. 1998.
Биологические ритмы — описание часов работы организма
Каждый организм имеет встроенные биологические часы, регулирующие химические процессы, возникающие как реакции на внутренние и внешние сигналы. Активность биологических часов человека не одинакова на протяжении суток. За 24 часа они то замедляются, то ускоряются.
Согласно китайской философии, человеческий организм подвержен трем ритмам: интеллектуальному, эмоциональному, и физическому. Эти три биоритма определяют все состояния тела, захватывая даже состояния молекул.
Суточный биоритм начинает функционировать с работы легких (с 3 до 5 часов – время тиньши), поскольку интенсификация активности бронхов и легких – один из начальных этапов пробуждения и подготовки к деятельному состоянию. Необходимо создать внешние условия (открыть форточку) для поступления большего количества кислорода, поскольку в это время мозг особенно в нем нуждается. При недостатке кислорода мозг «забирает последние накопления», и легкие «ослабевают». Таким образов, создаются предпосылки возникновения патологических измененияй в легких. Это время еще называют первой волной саморегуляции психики и физиологии.
- 3 часа минимальный уровень выработки мелатонина, отвечающего за наш спокойный сон, психические реакции.
 Те, кто не спит в это время в 5 раз больше страдают депрессиями.
Те, кто не спит в это время в 5 раз больше страдают депрессиями.
 Те, кто не спит в это время в 5 раз больше страдают депрессиями.
Те, кто не спит в это время в 5 раз больше страдают депрессиями.В 3 раза повышается склонность к онкологии.
- С 5 до 7 часов (время маоши) – время максимальной активности толстого кишечника, который готов к детоксикации.
Если нет опорожнения кишечника , то все яды и токсины возвращаются в кровеносную систему. Вот почему люди, страдающие запорами часто жалуются на головную боль. 5 утра – минимальная температура тела и минимальная активность почек.
В 6 часов срабатывает «биологический будильник».Оживают все системы организмов, активируется обмен веществ, увеличивается количество Биологические ритмы глюкозы и аминокислот в крови, повышается артериальное давление. На работу здорового организма оказывает благоприятное влияние обливание холодной водой. В 6 – 7 часов особенно сильна именная защита организма.
- С 7 до 9 часов (время чэньши) – время максимальной активности желудка. Все готово для восполнения ежедневной потребности организма в энергии. Завтрак должен быть полноценным, поскольку желудок перерабатывает не только пищу, но и жизненные события, включается деятельность стимулирующая к размышлениям. Если человек голоден, преобладают отрицательные эмоции, дающие плохие советы.
В период с 7 до 11 часов «сытый организм » начинает бороться с возможными еще скрытыми и проявившимися заболеваниями.
В 9 часов отмечается максимальный выброс гормона коры надпочечников – кортизона, которому отводят роль регулятора нашей активности и тонуса. В крови отмечается суточный минимум лимфоцитов, поэтому снижается иммунитет и повышается опасность заболеваний респираторными инфекциями.
- С 9 до 11 часов (время сыши) максимум работы системы селезенка – поджелудочная железа.
Не стоит принимать лекарственные препараты в это время. Можно дать возможность организму самому побороться с болезнью.
- 10 часов – пик кровообращения. Здоровый организм способен выполнять любые нагрузочные функции.

Особенно активно работают отделы мозга, отвечающие за выполнение математических действий.
- 11 – 13 часов (время уши) – время для самых сложных физических и эмоциональных нагрузок.
Организм проявляет наибольшую выносливость. Здоровый миокард справляется со всем, что предлагает ему жизнь. Люди с сердечно – сосудистыми заболеваниями должны быть предельно осторожны, так как в это время чаще всего могут проявляться осложнения. К 12 часам запас энергии начинает таять , и организм плохо справляется со стрессом,в это время можно перекусить.
- С 13 до 15 часов (время вэйши) максимально активна система тонкого кишечника.
Это период большого эмоционального и физического напряжения сил. Таким образом, период с 11 до 14 часов – время предельно возможных для организма нагрузок и способность к запоминанию на длительное время. Кроме того, это время, когда меньше всего ощущается боль – подходящий период сходить к стоматологу. 15 часов время максимальной утомляемости. Необходимо дать организму короткий, но эффективный отдых (провести оздоровительную релаксацию длительностью 5 – 10 минут).
- В 16 часов оживляется кровообращение, можно заниматься спортом.
С 15 до 17 часов (время шэньши) – солнце жизни (максимальная активность в системе мочевого пузыря). Начало второй волны саморегуляции психики и физиологии. Это время неторопливого и мудрого общения со своей душой, встречи с друзьями ( не следует видеться с недоброжелателями).
- С 17 до 19 часов (время юши) максимум активности системы почек. Более глубинная саморегуляция.
Можно вспомнить самое сокровенное, хорошее время для прогулок и решения важных вопросов. 19 – 21 час (время сюйши)– активна система перикарда, отвечающая за вегето – сосудистую нервную регуляцию. Можно снова давать организму высокие нагрузки, активно творить, поскольку система перикарда защищает сердце от эмоциональных бурь.
- С 21 до 23 часов (время цзыши)– солнце жизни в системе тройного обогревателя ) легкие, печень, почки.

Это время предназначено для серьезной работы по решению своих жизненных задач. К 21 часу в желудке постепенно снижается выработка желудочного сока, а около полуночи прекращается совсем. Поэтому обильная белковая пища может принести вред. Период с 19 до 21 часа и с 21 до 23 часов можно рассматривать как время работы организма «во вторую смену», выдерживая большие нагрузки. Это время, когда хорошо обдумывается информация, гениальные люди совершают открытия. К 22 часам усиливается выработка серотонина ( гормон хорошего настроения). Это лучшее время для романтических встреч.
- С 22 до 2 ночи к гениальным людям приходят открытия, хорошо обдумывается информация, а обычный человек страдает повышенным аппетитом.
С 23 до 1 часа ночи максимальная активность желчного пузыря. Эта система контролирует опорно двигательный аппарат и сексуальную сферу. Созданы условия для проявления агрессивности и гнева. Но, поскольку природа умна, она в это время укладывае человека спать, чтобы он не натворил бед. Во сне вся агрессия устремляется на восстановление сил уставшего организма. Однако, «совы» в это время могут работать без ущерба для своего здоровья. Очень полезно лечь спать именно в это время. 00 часов – благоприятное время для «самопочинки» организма. Быстрее заживают раны, повышается иммунитет.
- С 1 до 3 часов (время чоуши) ночи активна системы печени. А поскольку печень энергетически решает любые жизненные проблемы, то в этот период происходит интенсивный обмен веществ, и в организме отмечается «большая чистка».
У бодрствующих в это время людей отмечаются более частые случаи депрессивных состояний, нервные срывы. Таким образом, становится понятно, что сам организм – это точно настроенный механизм, работающий в гармонией с окружающей средой. Он может подстроиться под окружающую среду самостоятельно, нужно только не мешать ему.
(PDF) Принципы физиологии активности Н.А. Бернштейна в психологии восприятия и внимания: проблемы и перспективы [N.
 A. Bernstein’s Principles of Physiology of Activity in Psychology of Perception and Attention: Problems and Perspectives]
A. Bernstein’s Principles of Physiology of Activity in Psychology of Perception and Attention: Problems and Perspectives]КУЛЬТУРНО-ИСТОРИЧЕСКАЯ ПСИХОЛОГИЯ 2016. Т. 12. № 4
CULTURAL-HISTORICAL PSYCHOLOGY. 2016. Vol. 12, no. 4
65
Journal of the Higher School of Economics], 2008, vol. 5, no. 3,
pp. 31—66.
45. Falikman M.V. Effekty prevoskhodstva slova v
zritel’nom vospriyatii i vnimanii [Word superiority effects
in visual perception and attention]. Psikhologicheskii zhurnal
[Psychological journal], 2010, no. 1, pp. 32—40.
46. Falikman M.V., Pechenkova E.V. Strategicheskaya
regulyatsiya resheniya pertseptivnoi zadachi kak klass nisk-
hodyashchikh vliyanii na protsess postroeniya pertseptivnogo
obraza [Strategic regulation of perceptual task accomplishment
as a type of top-down influences on perceptual image construc-
tion]. Pervaya Rossiiskaya konferentsiya po kognitivnoi nauke.
Tezisy dokladov [1st Russian cognitive science conference. Ab-
stracts]. Kazan’: KGU, 2004, pp. 237—239.
47. Falikman M.V., Pechenkova E.V., Stepanov V.Yu.
Vnimanie i pertseptivnye edinitsy [Attention and perceptual
units]. In Chernigovskaya T.V. (eds.), Cognitivnye issledo-
vaniya [Cognitive research]. Vyp. 5. . Moscow: Publ. “Institut
psikhologii RAN”, 2012, pp. 102—116.
48. Feigenberg I.M. Nikolai Bernshtein: ot refleksa k mod-
eli budushchego [Nikolai Bernstein: From a reflex to a model of
future]. Moscow: Smysl, 2004.
49. Feigenberg I.M. Veroyatnostnoe prognozirovanie v
deyatel’nosti cheloveka i povedenii zhivotnykh [Probabilistic
forecasting in human activity and animal behaviour]. Moscow:
N’yudiamed, 2008.
50. Feigenberg I.M., Ivannikov V.A. Veroyatnostnoe prog-
nozirovanie i prednastroika k dvizheniyam [Probabilistic fore-
casting and movement presetting]. Moscow: Publ. Mosk. un-
Moscow: Publ. Mosk. un-
ta, 1978.
51. Adamian N., Kuvaldina M. Be prepared: The effect of
expectations on inattentional blindness. The Russian Journal of
Cognitive Science, 2014, vol. 1, no. 3, pp. 4—12.
52. Bernstein N. Co-ordination and regulation of move-
ments. N.Y.: Pergamon Press, 1967.
53. Brodski A., Paasch G.F., Helbling S., Wibral M. The
faces of predictive coding. Journal of Neuroscience, 2015,
vol. 35, no. 24, pp. 8997—9006.
54. Bruner J.S., Minturn A.L. Perceptual identification and
perceptual organization. The Journal of General Psychology,
1955, vol. 53, pp. 21—28.
55. Cavanagh P., Anstis S. The flash grab effect. Vision Re-
search, 2013, vol. 91, pp. 8—20.
56. Clark A. Whatever next? Predictive brains, situated
agents, and the future of cognitive science. Behavioral and
Brain Sciences, 2013, vol. 36, no. 3, pp. 181—204.
57. Craik F.I.M. Levels of processing: past, present … and
future? Memory, 2002, vol. 10, no. 5/6, pp. 305—318.
58. Craik F.I.M., Lockhart R. Levels of processing: A
framework for memory research. Journal of Verbal Learning
and Verbal Behaviour, 1972, vol. 11, no. 6, pp. 671—684.
59. De Baene W., Brass M. Switch probability context (in)
sensitivity within the cognitive control network. NeuroImage,
2013, vol. 77, pp. 207—214.
60. De Baene W., Brass M. Dissociating strategy-depen-
dent and independent components in task preparation. Neuro-
psychologia, 2014, vol. 62, pp. 331—340.
61. Di Lollo V., Enns J., Rensink R. Competition for con-
sciousness among visual events: the psychophysics of reentrant
visual processes. Journal of Experimental Psychology: General,
2000, vol. 129, no. 4, pp. 481—507.
62. Di Lollo V., Kawahara J., Zuvic S.M., Visser T.A. The
preattentive emperor has no clothes: a dynamic redressing.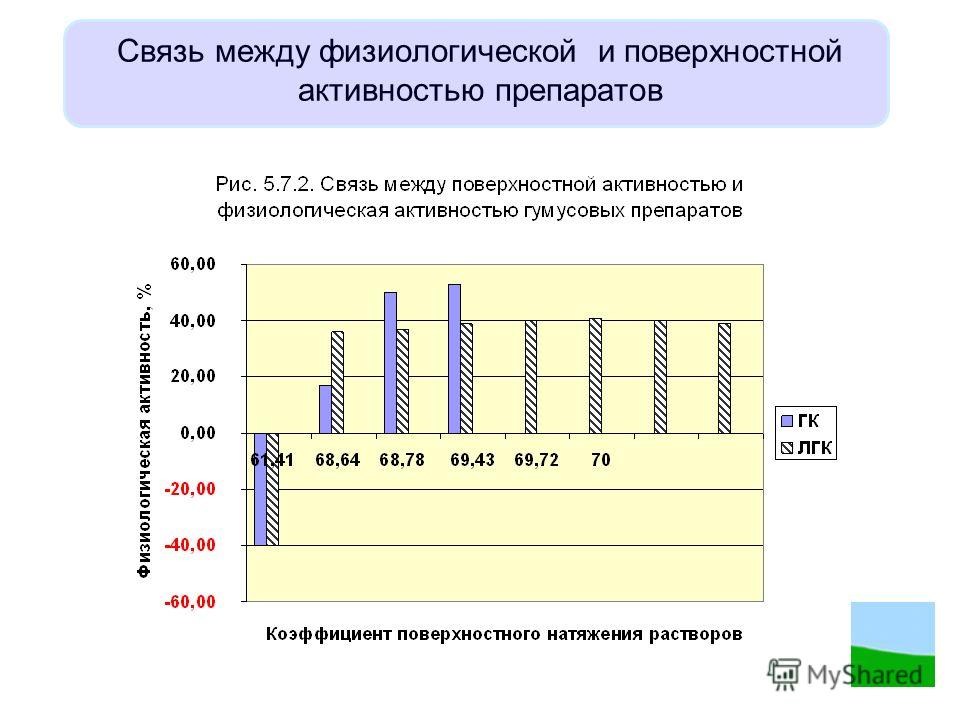
Journal of Experimental Psychology: General, 2001, vol. 130,
no. 3, pp. 479—492.
63. Di Lollo V., Smilek D., Kawahara J., Ghorashi S.M.
System reconfiguration, not resource depletion, determines
the efficiency of visual search. Perception and Psychophysics,
2005, vol. 67, no. 6, pp. 1080—1087.
64. Dux P.E., Visser T.A.W., Goodhew S.C., Lipp O.V.
Delayed re-entrant processing impairs visual awareness: An
object substitution masking study. Psychological Science, 2010,
vol. 21, pp. 1242—1247.
65. Eich T.S., Parker D., Liu D., Oh H., Razlighi Q., Gaz-
es Y., Habeck C., Stern Y. Functional brain and age-related
changes associated with congruency in task switching. Neuro-
psychologia, 2016, vol. 91, pp. 211—221.
66. Enns J.T., Di Lollo V. What’s new in visual masking?
Trends in Cognitive Science, 2000, vol. 4, no. 9, pp. 345—352.
67. Enns J.T., Visser T.A.W., Kawahara J., Di Lollo V. Visual
masking and task switching in the attentional blink. In: Shapiro
K.L. (Ed.) The limits of attention: Temporal constraints on human
information processing. Oxford University Press, 2001, pp. 65—81.
68. Falikman M.V. Word superiority effects across the va-
rieties of attention. Journal of Russian and East European Psy-
chology, 2011, vol. 49, no. 5, pp. 45—61.
69. Goschke T. Intentional reconfiguration and involun-
tary persistence in task set switching. In S. Monsell, J. Driver
(Eds.), Control of cognitive processes: Attention and perfor-
mance XVIII. Cambridge, MA: MIT Press, 2000, pp. 331—355.
70. Grange J., Houghton G. (Eds.) Task switching and cog-
nitive control. Oxford University Press, 2014.
71. Greene M.R., Botros A.P., Beck D.M., Fei-Fei L. What
you see is what you expect: Rapid scene understanding benefits
from prior experience. Attention, Perception and Psychophysics,
Attention, Perception and Psychophysics,
2015, vol. 77, no. 4, pp. 1239—1251.
72. Gregory R. Knowledge in perception and illusion. Phil-
osophical Transactions of the Royal Society B: Biological Sci-
ences, 1997, vol. 352, no. 1358, pp. 1121—1127.
73. Gurfinkel V.S., Cordo P.J. The scientific legacy of
Nikolai Bernstein. In M.L. Latash (Ed.) Progress in motor
control: Volume 1, Bernstein’s traditions in movement studies.
Champaign: Human Kinetics, 1998, pp. 1—20.
74. Hochstein, S., Ahissar, M. View from the top: Hier-
archies and reverse hierarchies in the visual system. Neuron,
2002, vol. 36, pp. 791—804.
75. Hübner M., Kluwe R.H., Luna—Rodriguez A., Peters
A. Task preparation and stimulus—evoked competition. Acta
Psychologica, 2004, vol. 115, pp. 211—234.
76. James W. Principles of Psychology. Vol. 1. New York:
Henry Holt, 1890.
77. Jonides J., Gleitman N. A conceptual category effect in
visual search: O as letter or as digit. Perception & Psychophys-
ics, 1972, vol. 12, pp. 457—460.
78. Kawahara J., Di Lollo V., Enns J.T. Attentional require-
ments in visual detection and identification: Evidence from the
attentional blink. Journal of Experimental Psychology: Human
Perception and Performance, 2001, vol. 27, no. 4, pp. 969—984.
79. Kelso J.A.S. From Bernstein’s psysiology of activity
to coordination dynamics. In Latash M.L. (ed.), Progress in
motor control: Volume 1, Bernstein’s traditions in movement
studies. Champaign: Human Kinetics, 1998, pp. 203—220.
80. Kesner L. The predictive mind and the experience
of visual art work. Frontiers in Psychology, 2014, vol. 5, art.
1417. URL: http://journal.frontiersin.org/article/10.3389/
fpsyg.2014.01417/full (Accessed: 09.11.2016).
81. Kogo N., Trengove C. Is predictive coding theory ar-
Is predictive coding theory ar-
ticulated enough to be testable? Frontiers in Computational
Neuroscience, 2015, vol. 9, art. 111. URL: http://journal.fron-
tiersin.org/article/10.3389/fncom.2015.00111/full (Accessed:
09.11.2016).
82. Latash M.L. (Ed.) Progress in motor control. Vol-
ume 1: Bernstein’s traditions in movement studies. Urbana, IL:
Human Kinetics, 1998.
Биологически активные добавки. Еда или лекарство?
Биологически активные добавки — компоненты натуральных или идентичных натуральным биологически активных веществ – предназначены для непосредственного приёма или введения в состав пищевых продуктов с целью обогащения рациона питания человека биологически активными веществами или их комплексами.
Биологически активные добавки разделяются на 3 группы.
- Нутрицевтики представляют собой эссенциальные биологически активные вещества, которые являются основными компонентами пищи: витамины или их предшественники, макро- и микроэлементы (железо, кальций, селен, цинк, фтор и т.д.), полиненасыщенные жирные кислоты, незаменимые аминокислоты, некоторые моно- и дисахариды, пищевые волокна, применяемые для коррекции химического состава пищи человека.
- Парафармацевтики – биологически активные вещества, обладающие определённой активностью и применяемые для профилактики и поддержки в физиологических границах функциональной активности органов и систем. К ним относятся биофлавоноиды, алкалоиды, гликозиды, сапонины, органические кислоты, эфирные масла, полисахариды, то есть это препараты растительного происхождения.
- Эубиотики или пробиотики — биологически активные добавки к пище, в состав которых входят живые микроорганизмы и (или) их метаболиты, оказывающие нормализующее воздействие на состав и биологическую активность микрофлоры пищеварительного тракта.
В России биологически активные добавки впервые начали появляться в 1995 году, и с каждым годом уровень потребления населением БАД растет. Почему люди охотно их принимают? Наш организм практически никогда не получает в нужной пропорции витамины и минералы, без которых он не в состоянии функционировать и «строить» здоровые клетки и ткани. Выбирая в магазине продукты, мы чаще всего не задумываемся, какой «строительный материал» ему поставляем. И, когда в организме истощается необходимый баланс между минералами и витаминами, микронутриентами наш иммунитет ослабевает, и мы начинаем чаще болеть, возрастает тяжесть простудных и других инфекционных заболеваний.
Почему люди охотно их принимают? Наш организм практически никогда не получает в нужной пропорции витамины и минералы, без которых он не в состоянии функционировать и «строить» здоровые клетки и ткани. Выбирая в магазине продукты, мы чаще всего не задумываемся, какой «строительный материал» ему поставляем. И, когда в организме истощается необходимый баланс между минералами и витаминами, микронутриентами наш иммунитет ослабевает, и мы начинаем чаще болеть, возрастает тяжесть простудных и других инфекционных заболеваний.
Как считает Б.Суханов, профессор кафедры гигиены питания и токсикологии Московской медицинской академии им. Сеченова, применение БАДов по рекомендации врача, согласно прилагаемой инструкции, принесёт человеку только пользу. Биологически активные добавки сами по себе являются концентрированным источником дозировано подобранных дефицитных веществ, участвующих в строительстве мышечных, костных, клеточных тканях нашего организма. С проблемой неправильного питания и возникновения в последствие такого рода дефицита сталкивается подавляющее большинство населения России.
Не являясь по своей сути едой, а только витаминно-минеральной добавкой к ней, БАДы по определению не являются и лекарством, какие бы чудеса исцеления ни обещала при этом хорошо продуманная реклама продукта. Наличие веществ растительного и животного происхождения в продукте обладает только определённой фармакологической активностью и не может заменить собой лекарственные препараты.
Рынок продаж БАДов в России расширяется, вместе с этим строже становятся и требования к ним: продукт должен иметь свидетельство о государственной регистрации и не содержать в себе сильнодействующие, наркотические и ядовитые вещества.
В свою очередь, приобретая полезный и эффективный «добавочный» продукт, нам надо отдавать себе отчёт, что БАД, как и лекарство, обязательно нам помогут, если применяются по установленным правилам и потребность нашего организма в их приёме действительно существует.
Использовать БАД без консультации с врачом не рекомендуется, поскольку компоненты БАД могут взаимодействовать с лекарственными средствами, изменяя их активность. Консультации необходимы и для определения показаний, выявления противопоказаний у каждого конкретного человека, объяснения рекомендаций по применению в зависимости от пола, возраста, наличия сопутствующих заболеваний, дачи характеристики отдельных компонентов, входящих в состав БАД. Особое значение имеет взаимодействие БАД с лекарственными препаратами, которые одновременно принимают больные люди.
Будьте предельно внимательны, при употреблении БАД. Во избежание развития нежелательных эффектов, консультация с врачом является необходимой!
Заместитель начальника ЦГСЭН МСЧ Е.А.Лебедева.
Двигательная активность и здоровье человека.
Двигательная активность и здоровье человека.
Здоровый образ жизни неотделимо связан с высокой двигательной активностью человека. В настоящее время на людей влияют многие неблагоприятные факторы внешней среды, большой поток информации, сложные социальные условия жизни. Что неизменно приводит к эмоциональному напряжению и снижению двигательной активности.
Развитие наших предков происходило в каждодневной борьбе за жизнь и сопровождалось огромными мышечными напряжениями. В настоящее время доказано, что опорно-двигательный аппарат, органы кровообращения и дыхания, функции нервной системы и даже железы секреции смогут правильно развиваться и функционировать, лишь при условии достаточной и регулярной мышечной нагрузки.
Современный образ жизни ведет к снижению двигательной активности
Работа, связанная с физическими нагрузками, требующая выносливости и длительных мышечных напряжений, в современной промышленности, транспорте и сельском хозяйстве исчезает. Ей на смену приходит механизация труда. Снижает сумму мышечных усилий «кнопочное» управление различными механизмами и распространение личного транспорта. Облегчая нашу жизнь и бытовые условия, это лишает наш организм мышечных усилий и тем самым оказывает на него неблагоприятное влияние.
Облегчая нашу жизнь и бытовые условия, это лишает наш организм мышечных усилий и тем самым оказывает на него неблагоприятное влияние.
Нехватку мышечных напряжений нужно корректировать при помощи специальных оздоровительных мероприятий
Для организма становится особенно важным значение мышечных нагрузок при таких заболеваниях как гипокинезия (снижении двигательной активности) и гиподинамия (снижении мышечных усилий). Сопутствовать недостатку двигательной активности могут атрофия и дегенерацией скелетных мышц. Происходит истончение мышечных волокон, снижение веса и тонус мышц.
К чему приводит недостаток движения?
Результатом гипокинезии или гиподинамии становятся существенные изменения и нарушения координации движений, ухудшается состояние зрительного, вестибулярного и двигательного аппаратов. Происходят изменения в кровеносной системе: уменьшается размер сердца, учащается пульс, уменьшается масса циркулирующей крови, увеличивается время ее кругооборота. Снижаются функции надпочечников.
Для нашего организма двигательная активность является физиологической потребностью. Лишённый движения организм теряет способность накапливать энергию, необходимую для противостояния стрессу. Мышечные напряжения, воздействие контрастных температур, принятие солнечных ванн в разумной мере полезны организму.
Для восполнения подвижности незаменимы занятия физической культурой, способствующие улучшению деятельности нервных центров, процессов мышления, памяти, концентрации внимания, точной ориентации человека в пространстве, повышению резервов многих систем организма. При регулярных занятиях физическими упражнениями повышается емкость легких, объем и глубина дыхания, нормализуется деятельность желез внутренней секреции.
Повышенная мышечная деятельность может стать стрессом. Если физические нагрузки на организм будут чрезмерными, может появиться первая или вторая стадия стресса, которая при длительном действии напряжений способна перейти в третью — истощение.
Исследования показали, что при постепенном увеличении интенсивности и длительности упражнений в организме не наблюдается патологических изменений.
Утренние физические упражнения имеют достаточно важное значение для повышения работоспособности человека после сна, для укрепления здоровья и закаливания организма. Способствуют восстановлению нормального функционирования центральной нервной системы утренние занятия гимнастикой.
Солнце, воздух и вода — наши лучшие друзья!
Восстановление нервной системы ускоряется, если сочетать утреннюю зарядку с водными процедурами. Воздействие таких раздражителей как температурный фактор, действие свежего воздуха, солнца или воды способствует закаливанию организма человека, помогая снять отечность тканей, а именно вен, которая бывает после пробуждения.
Важность физических упражнений по утрам не ограничивается устранением последствий сна. Они способствуют совершенствованию силы, выносливости, координации и быстроты нашего тела, а также укреплению сердечно-сосудистой, выделительной, дыхательной и других вегетативных систем.
Ежедневно выполняя упражнения, вы будете поддерживать на должном уровне резервную щелочность крови и способствовать сохранению специфических свойств мышц, которые развиваются при регулярной мышечной деятельности.
Чтобы не ухудшать последующую работоспособность, надо избежать большой активности при выполнении физических упражнений до начала вашего рабочего дня.
Не менее важны по значению упражнения, выполняемые на протяжении рабочего дня и даже после его окончания. Они запускают механизм активного отдыха, способствующий скорому восстановлению разных функций в организме, нарушенных из-за утомления. Упражнения являются самым эффективным средством снимающим нервное и психическое напряжение. В настоящее время для эффективной борьбы с гиподинамией разработаны несколько методик физических упражнений. Стоит особенно выделить шейпинг, спортивные игры, ритмическую гимнастику, плавание, велосипедные прогулки и бег. Каждый делает выбор системы исходя из интереса, возможностей и запросов.
Стоит особенно выделить шейпинг, спортивные игры, ритмическую гимнастику, плавание, велосипедные прогулки и бег. Каждый делает выбор системы исходя из интереса, возможностей и запросов.
Правильная организация процесса — очень важна.
Важно понимать, что сама физическая активность при неправильном использовании не даст оздоровительного результата. Степень физической нагрузки должна быть оптимальной для каждого отдельного человека. Необходимо соблюдение принципов, гарантирующих оздоровительный эффект. И главными из них выступают последовательность, постепенность, регулярность и систематичность физических тренировок. Тренированность организма появляется только в процессе занятий.
Главное отличие тренированного организма — умением быстро включить резервы в действие и экономно обеспечивать их координацию. Сдвиги происходящие в организме, при мышечной деятельности, являются фазовыми и не сохраняются долгое время. Именно поэтому, чтобы развить тренированность важно не допускать больших интервалов отдыха между упражнениями.
Информация: http://vsezdorovo.com/2011/11/activity-11/2/
Направления научно-исследовательской деятельности кафедры «Ботаника, физиология и защита растений»
Цифровые технологии в агрономии (Крутова Е.К.)
Цифровая трансформация сельского хозяйства позволит увеличить производительность труда в АПК, повысить уровень автоматизации основных производственных процессов, повысить привлекательность работы в сельском хозяйстве и, тем самым, позволит решить проблему кадров агрономов, сохранить плодородие почв.
Агрономический факультет предлагает:
- Создание цифровых карт, картограмм и формирование агроэкологических паспортов полей
- Составление цифровых тематических карт полей – культур, гумус и др.
- Составление агроэкологических паспортов полей
- Мониторинг посевов при помощи БЛА
- Составление картограмм индекса NDVI
- Составление систем земледелия на основе анализа полученных данных и сервисов дистанционного мониторинга
Утилизация отходов АПК методом термодинамической бифуркации (Михалев Е. В.)
В.)
Наши специалисты готовы оказать помощь в проектировании и налаживании производства новых видов органических удобрений (сложных компостов) на основе термодинамической бифуркации компостируемой массы.
- Объекты применения – крупные животноводческие комплексы (птицефабрики, свинокомплексы, фермы КРС).
- Сроки компостирования каждой партии – до 1 месяца (производство работает постоянно в течение круглого года).
- Эффективность производства – до 300%.
- КПД технологии возрастает до 50-500%.
- Экологическая значимость – улучшается экологическая ситуация на территориях, прилегающих к животноводческим объектам.
- Социальная значимость – создание новых рабочих мест.
Состав, содержание и особенности биосинтеза масла в плодах с сочным околоплодником (Бережная Г.А.)
Изучение процесса маслообразования в плодах с сочными околоплодниками представляет несомненный теоретический интерес и практическую ценность. Биологическое значение масла семян известно и состоит в его использовании в качестве источника энергии и органических веществ при прорастании семян. Физиологическая роль масел плодовой мякоти не установлена, однако интерес к их изучению постоянно возрастает, благодаря высокой физиологической активности таких масел, необычному жирнокислотному составу и присутствию многочисленных биологически активных веществ.
В последние годы интерес к подобным исследованиям возрастает в связи с использованием различных органов растений, в том числе и плодов с сочными околоплодниками для получения медицинских препаратов и БАДов. Этому способствует внедрение новых технологий, позволяющих выделять из растительных тканей высокоэффективные, нетоксичные препараты, обладающие высокой физиологической активностью.
Цель и задачи работы — продолжить изучение липидозапасающей функции околоплодников и семян сочных плодов, как целостной биологической системы, а также формирование анатомических структур, накопления масла в клетках этих органов и взаимосвязь между околоплодником и семенем в процессе формирования и развития плода. Расширить изучение числа плодов с сочными околоплодниками, используемых в медицинской практике, что может способствовать пониманию закономерностей липидообразования. Выделить, идентифицировать и изучить, биологически активные вещества околоплодников, и их физиологическую активность для использования в медицине и пищевой промышленности.
Расширить изучение числа плодов с сочными околоплодниками, используемых в медицинской практике, что может способствовать пониманию закономерностей липидообразования. Выделить, идентифицировать и изучить, биологически активные вещества околоплодников, и их физиологическую активность для использования в медицине и пищевой промышленности.
Изучение эндофитных бактерий картофеля как ресурс профилактики фитофтороза (Ионова Г.Б.)
Эндофитными называются бактерии, способные колонизировать внутренние ткани растения, не вызывая его заболеваний и не оказывая отрицательного влияния на развитие.
Эндофитные бактерии занимают те же экологические ниши, что и болезнетворные микроорганизмы, поэтому могут быть использованы в качестве перспективного биологического средства борьбы с фитопатогенами.
Задачи исследования:
- выделение и описание эндофитных бактерий из клубней картофеля;
- изучение фунгицидных (возможно, и бактерицидных) свойств эндофитных бактерий;
- изучение ростостимулирующей активности эндофитных бактерий.
Снижение ущерба, наносимого патогенными грибами (остальное надо считать). Ущерб агрономический – снижение урожая, экономический – денежные потери.
Разработка приемов агротехники возделывания конопли среднерусской в условиях Нижегородской области (Михалев Е.В., Насонова Л.В.)
Конопля является экономически значимой технической культурой. Она представляет большой интерес для предприятий АПК области и практическую значимость для перерабатывающей промышленности.
Для успешного развития коноплеводства в условиях Нижегородской области необходимо разработать адаптированную технологию культивирования конопли с учетом ее сортовых особенностей, для чего необходимо изучить новые сорта, разработаны на абазе Пензенского НИИСХ.
Задачи: Изучить сроки, схемы, нормы и способы посева конопли трех сортов — Вера, Сурская, Надежда; разработать элементы защиты растений конопли от вредителей и болезней; изучить различные фоны минерального питания при выращивании сортов конопли; определить оптимальный режим полива и внесения органических и минеральных удобрений.
За счет внедрения в производство конопли среднерусской ожидается оптимизация структуры посевных площадей, при этом резко вырастет выход конечной продукции с единицы площади, что позволит существенным образом повысить эффективность производства за счет получения дополнительной прибыли.
Разработка приемов адаптивной системы защиты картофеля в условиях Нижегородской области (Михалев Е.В., Мухина О.В., Насонова Л.В.)
В настоящее время перед предприятиями АПК Нижегородской области остро встает вопрос об эффективности возделывания картофеля в условиях повышенной нагрузки, которая приводит к значительному недобору урожая и к последующим потерям при хранении. Это обусловлено тем, что большинство сортов картофеля недостаточно адаптированы к постоянно изменяющимся условиям внешней среды.
Сравнительное изучение различных сортов картофеля в условиях опытного участка НГСХА позволит изучить различные способы посадки картофеля (глубина, норма, ширина междурядий), влияние внесения органических удобрений (отходов) и полива на урожайность картофеля, проведение учета вредителей и болезней, а так же применение приемов сидерации (до посадки, уплотнения и после уборки).
Ожидаемый эффект заключается в увеличении урожайности и качества продукции, а так же в повышении устойчивости растений к вредителям и болезням.
Разработка новых агробиотехнологических приемов и биопрепаратов для нужд сельского хозяйства (Веселов А.П.)
Биотехнологические работы по микроклональному размножению растений, выявлению особенностей синтеза и накопления в них хозяйственно – ценных веществ.
Разработка приемов агротехники возделывания льна в условиях Нижегородской области (Михалев Е.В., Насонова Л.В.)
Лен является экономически значимой технической культурой. Он представляет большой интерес для предприятий АПК области (особенно в северных районах) и практическую значимость для перерабатывающей промышленности.
Для успешного развития льноводства в условиях Нижегородской области необходимо разработать адаптированную технологию культивирования льна с учетом его сортовых особенностей, для чего необходимо изучить новые сорта, сроки, схемы, нормы и способы посева; разработка элементов защиты растений льна от вредителей и болезней, изучение различных фонов минерального питания; изучение режимов полива.
За счет внедрения в производство льна ожидается оптимизация структуры посевных площадей, при этом резко вырастет выход конечной продукции с единицы площади, что позволит существенным образом повысить эффективность производства за счет получения дополнительной прибыли.
Исследование эффективности применения новых, комплексных органо-минеральных удобрений на основе отходов грибоводства (Тарасов С.С.)
Грибы ключевые организмы в экосистемах, являясь рудуцентами, они перерабатывают органические вещества в минеральные, т.е. обеспечивают круговорот веществ в природе. Проблема применения удобрений важна с одной стороны для получения высокопродуктивной продукции, а с другой снижение их негативного воздействия на экосистемы. Важно разработать такие удобрения, которые бы, с одной стороны максимально эффективно обеспечивали продуктивность растений, а с другой бы минимизировали негативное воздействие на окружающую среду. Изучается возможность применения отходов выращивания целлюлозоразрушающих и сапротрофных грибов в качестве органических удобрений, а так же их влияние на физилого-биохимические и продуктивные показатели растений. Использование отходов грибоводства позволит перевести в полный замкнутый цикл аграрное производство.
Свободно-радикальные процессы в хозяйственно значимых биообъектах в зависимости от модифицирующих и стресс факторов (Тарасов С.С.)
Свободно-радикальные процессы наиболее важные физиолого-биохимические показатели состояния живых организмов, в том числе и сельскохозяйственных биообъектов, их изучение значимо для понимания фундаментальных механизмов жизнедеятельности с целью получения максимальной и безопасной продукции. Проводятся эксперименты по влиянию различных экологических факторов на показатели окислительного гомеостаза и антиоксидантного статуса биообъектов в зависимости от модифицирующих и стресс факторах. Как итог можно выявить наиболее оптимальные условия жизнедеятельности живых организмов и установить наиболее благоприятные диапазоны свободно-радикальной толерантности к действию экологических факторов.
Проводятся эксперименты по влиянию различных экологических факторов на показатели окислительного гомеостаза и антиоксидантного статуса биообъектов в зависимости от модифицирующих и стресс факторах. Как итог можно выявить наиболее оптимальные условия жизнедеятельности живых организмов и установить наиболее благоприятные диапазоны свободно-радикальной толерантности к действию экологических факторов.
Разработка и исследования кормов на основе нетрадиционного растительного сырья (Тарасов С.С.)
Производства экологически чистых кормов одна из ключевых задач современного сельского хозяйства. Для решения этой проблемы стоит исследовать возможность применения нетрадиционных растений в качестве кормового сырья. Что может уменьшить затраты на кормление животных, сохраняя при этом их высокую продуктивность и безопасность итоговой продукции.
Дальневосточный научный центр физиологии и патологии дыхания
Федеральное государственное бюджетное научное учреждение «Дальневосточный научный центр физиологии и патологии дыхания» (ДНЦ ФПД) — это крупнейшее на Дальнем Востоке динамично развивающееся научно-медицинское учреждение. ДНЦ ФПД занимает одно из лидирующих мест по продуктивности научно-исследовательской работы, о чем свидетельствуют показатели публикационной активности, в том числе и в международных базах данных Web of Science и Scopus.
В составе Центра работают 2 академических филиала: Владивостокский филиал — Научно-исследовательский институт медицинской климатологии и восстановительного лечения и Хабаровский филиал — Научно-исследовательский институт охраны материнства и детства.
Расширяются международные связи: с ВОЗ по реализации национального проекта Глобального альянса по борьбе с болезнями органов дыхания; с профессиональными международными научными сообществами врачей; с зарубежными фармацевтическими компаниями путем участия в многоцентровых клинических исследованиях новых лекарственных препаратов; с учеными Китая в рамках реализации межгосударственной программы научно-технического сотрудничества.
Сотрудники центра регулярно принимают участие в крупнейших международных научных форумах. Опыт работы широко представлен в отечественных и зарубежных научных журналах и монографиях. ДНЦ ФПД проводит крупные научно-практические мероприятия и на собственной базе: съезды, конференции, школы для врачей.
Как научно-образовательный центр ДНЦ ФПД осуществляет подготовку научно-педагогических кадров высшей квалификации в аспирантуре по направлениям фундаментальная и клиническая медицина. На базе ДНЦ ФПД функционирует диссертационный совет по специальностям «Пульмонология» и «Патологическая физиология», выпускается научно-практический журнал «Бюллетень физиологии и патологии дыхания», входящий в перечень ВАК.
ДНЦ ФПД является уникальным по своим кадровым и техническим возможностям учреждением и занимает важное место в системе оказания специализированной пульмонологической помощи населению Амурской области. Центр располагает высококвалифицированными кадрами, среди которых доктора и кандидаты медицинских наук. Наличие пульмонологического стационара и современного лечебно-диагностического комплекса дает возможность пациентам в одном месте получить полный комплекс услуг по диагностике и лечению заболеваний органов дыхания.
Физиологическое поглощение в FDG PET, моделирующем болезнь
Biomed Imaging Interv J. 2006 окт-декабрь; 2 (4): e59.
Кафедра биомедицинской визуализации медицинского факультета Малайского университета, Куала-Лумпур, Малайзия
* Автор, ответственный за переписку. Нынешний адрес: Отделение биомедицинской визуализации медицинского факультета Малайского университета, 50603 Куала-Лумпур, Малайзия. Тел .: + 603-79492091; Факс: + 603-79581973; Электронная почта: ym.ude.mu@dar_halizas (Сазила Ахмад Сарджи).Поступило 20 сентября 2006 г .; Пересмотрено 8 ноября 2006 г .; Принята в печать 24 декабря 2006 г.
Copyright © 2006 Biomedical Imaging and Intervention Journal Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы.
Abstract
Многие потенциальные ловушки и артефакты были описаны в ПЭТ-визуализации, в которой используется фтордезоксиглюкоза F-18 (FDG).Нормальное поглощение ФДГ происходит во многих частях тела и может вызвать путаницу в интерпретации, особенно при онкологической визуализации. Клиническая корреляция, осведомленность об областях нормального поглощения ФДГ в организме и информация об изменениях в поглощении, а также о доброкачественных процессах, которые активно используются в ФДГ, необходимы, чтобы избежать потенциальных ошибок при интерпретации изображений. В этом контексте может быть начата оптимальная подготовка пациентов к сканированию, чтобы попытаться уменьшить проблему. Многие проблемы и подводные камни, связанные с областями нормального поглощения ФДГ, можно решить с помощью ПЭТ-КТ.ПЭТ-КТ позволяет правильно отнести активность ФДГ к структурно нормальному органу на КТ. Однако разработка комбинированных ПЭТ-КТ-сканеров также сопряжена со своими специфическими проблемами, связанными с комбинированной ПЭТ-КТ техникой. К ним относятся артефакты неправильной регистрации из-за дыхания и присутствие веществ с высокой плотностью, которые могут привести к артефактам завышенной оценки активности, если данные КТ используются для коррекции ослабления.
Ключевые слова: ФДГ ПЭТ, физиологическое поглощение, подводные камни, артефакты
ВВЕДЕНИЕ
Фтордезоксиглюкоза (ФДГ) F-18 является радиоиндикатором, наиболее часто используемым для ПЭТ-визуализации.Молекулу легче всего понять, интерпретируя ее в обратном направлении, начиная с молекулы глюкозы. Дезоксигруппа подразумевает отщепление гидроксильной группы от глюкозы. Присоединение индикатора F-18 к глюкозе заменяет гидроксильную группу.
Таким образом, молекула ФДГ действует как глюкоза во время начальных ферментативных реакций внутри клеток, но измененная структура препятствует дальнейшему метаболизму. Это по существу захватывает ФДГ внутри клеток, и ФДГ накапливается в большинстве тканей со скоростью, пропорциональной гликолизу [1]. Злокачественные клетки имеют повышенное содержание белков-переносчиков глюкозы на своей клеточной поверхности, а также повышенную скорость гликолиза. Повышенная гликолитическая скорость злокачественных клеток облегчает их обнаружение с использованием изображений ПЭТ-ФДГ. Именно этот атрибут заставляет метаболически активные опухоли выглядеть «горячими» на снимках ПЭТ. ПЭТ измеряет удержание ФДГ на объем ткани. Известно, что некоторые опухоли имеют высокое удержание ФДГ, в то время как другие имеют различное удерживание ФДГ. К сожалению, ФДГ не является агентом, специфичным для рака, и его поглощение было описано при ряде воспалительных поражений, включая саркоидоз, туберкулез, грибковую инфекцию и церебральный абсцесс.Повышенное накопление, вероятно, связано с заметно увеличенной скоростью гликолиза в активированных воспалительных клетках [2,3,4]. Тальковый плевродез вызывает висцеральное и париетальное гранулематозное воспаление плевры. Сообщалось об увеличении активности ФДГ в участках плеврального талька в течение трех лет после процедуры, предположительно вторично по отношению к воспалению плевры [5, 6].
Злокачественные клетки имеют повышенное содержание белков-переносчиков глюкозы на своей клеточной поверхности, а также повышенную скорость гликолиза. Повышенная гликолитическая скорость злокачественных клеток облегчает их обнаружение с использованием изображений ПЭТ-ФДГ. Именно этот атрибут заставляет метаболически активные опухоли выглядеть «горячими» на снимках ПЭТ. ПЭТ измеряет удержание ФДГ на объем ткани. Известно, что некоторые опухоли имеют высокое удержание ФДГ, в то время как другие имеют различное удерживание ФДГ. К сожалению, ФДГ не является агентом, специфичным для рака, и его поглощение было описано при ряде воспалительных поражений, включая саркоидоз, туберкулез, грибковую инфекцию и церебральный абсцесс.Повышенное накопление, вероятно, связано с заметно увеличенной скоростью гликолиза в активированных воспалительных клетках [2,3,4]. Тальковый плевродез вызывает висцеральное и париетальное гранулематозное воспаление плевры. Сообщалось об увеличении активности ФДГ в участках плеврального талька в течение трех лет после процедуры, предположительно вторично по отношению к воспалению плевры [5, 6].
ПЭТ-сканирование с ФДГ (коронарный вид) у пациента, перенесшего плевродез. Повышенная активность наблюдается вдоль плевры и трещины в правом гемитораксе [Изображение любезно предоставлено Центром молекулярной визуализации, Онкологический центр Питера МакКаллума, Восточный Мельбурн, Виктория, Австралия].
Было описано много потенциальных ловушек и артефактов, связанных с визуализацией FDG PET. Важно, чтобы мы узнали об этих ловушках и осознали важные области нормального поглощения ФДГ или отсутствия поглощения, которые могут не иметь значения. Это необходимо, чтобы пациенты могли быть оптимально подготовлены к сканированию и могли сделать точную интерпретацию.
ПОТРЕБЛЕНИЕ ФДГ В ОБЛАСТИ ГОЛОВКИ И ШЕИ
В головном мозге очень интенсивное поглощение метки происходит в нормальной коре головного мозга и базальных ганглиях, при этом глюкоза является преобладающим субстратом для метаболизма мозга. Общее поглощение в головном мозге составляет примерно 6% от введенной дозы. Нормальная лимфатическая ткань может демонстрировать поглощение ФДГ от низкого до умеренного в области головы и шеи. Это наблюдается в язычных и небных миндалинах и у основания языка из-за физиологической активности, связанной с лимфатической тканью в кольце Вальдейера. Симметрия помогает оценить поглощение ФДГ в голове и шее. Захват должен быть симметричным в небных и язычных миндалинах. Мягкое небо также может показывать поглощение метки.Переменное, но обычно низкое поглощение можно увидеть в слюнных железах. Во время разговора в гортани может накапливаться индикатор. Асимметричное поглощение ФДГ может наблюдаться в мышцах гортани у пациентов с параличом гортанного нерва, противоположным стороне дисфункции нерва, и его не следует неправильно интерпретировать как патологическое ().
Общее поглощение в головном мозге составляет примерно 6% от введенной дозы. Нормальная лимфатическая ткань может демонстрировать поглощение ФДГ от низкого до умеренного в области головы и шеи. Это наблюдается в язычных и небных миндалинах и у основания языка из-за физиологической активности, связанной с лимфатической тканью в кольце Вальдейера. Симметрия помогает оценить поглощение ФДГ в голове и шее. Захват должен быть симметричным в небных и язычных миндалинах. Мягкое небо также может показывать поглощение метки.Переменное, но обычно низкое поглощение можно увидеть в слюнных железах. Во время разговора в гортани может накапливаться индикатор. Асимметричное поглощение ФДГ может наблюдаться в мышцах гортани у пациентов с параличом гортанного нерва, противоположным стороне дисфункции нерва, и его не следует неправильно интерпретировать как патологическое ().
ПЭТ-сканирование с ФДГ (осевой вид) через гортань. Наблюдается асимметричный захват индикатора в мышцах гортани, в то время как у этого пациента с параличом левого гортанного нерва наблюдается немного более интенсивное поглощение с правой стороны (стрелка) [Изображение любезно предоставлено Центром молекулярной визуализации, Онкологический центр Питера МакКаллума, Восточный Мельбурн, Виктория, Австралия].
Умеренное поглощение можно увидеть в передней части дна рта из-за подбородочно-язычной мышцы, которая не позволяет языку откидываться назад у пациентов, находящихся в положении лежа на спине. Шейный отдел спинного мозга также может демонстрировать нормальное поглощение, что лучше всего видно на сагиттальных изображениях. Диффузное симметричное поглощение можно увидеть в нормальной щитовидной железе примерно в 2% сканирований. Диффузное поглощение тироидом может происходить в связи с тиреоидитом или болезнью Грейвса. Очаговое поглощение щитовидной железы может происходить при автономно функционирующих узлах щитовидной железы и злокачественных новообразованиях щитовидной железы.Пациентов с очаговым захватом следует дополнительно обследовать из-за более высокого риска того, что результат будет связан со злокачественными новообразованиями [7, 8] ().
ПЭТ с ФДГ и слитые ПЭТ КТ изображения (вид в осевом направлении) на уровне щитовидной железы. Очаговое интенсивное поглощение индикатора было случайно обнаружено в левой доле щитовидной железы [Изображение любезно предоставлено Центром молекулярной визуализации, Онкологический центр Питера МакКаллума, Восточный Мельбурн, Виктория, Австралия].
ПОНИМАНИЕ ФДГ В ТОЛАКЕ, ЖИВОТНОМ И ТАЗЕ
Поглощение в тимусе обычно наблюдается у детей.Однако не у всех детей будет видна активность тимуса на изображениях ПЭТ с ФДГ. Причина такой изменчивости накопления ФДГ в тимусе у детей неясна, но, вероятно, связана с физическими и эмоциональными стрессорами, которые влияют на метаболизм тимуса. В целом, физиологический захват ФДГ вилочковой железой исчезает в подростковом возрасте вместе с инволюцией вилочковой железы.
Отскок гиперплазии тимуса наблюдается у молодых пациентов, леченных от злокачественных новообразований. После химиотерапии поглощение ФДГ можно увидеть в тимусе 75% детей и от 5% до 16% взрослых, а увеличение может сохраняться до шести месяцев после завершения химиотерапии.Есть несколько признаков, свидетельствующих о непатологическом накоплении ФДГ в тимусе. Нормальная активность тимуса будет иметь треугольную или V-образную форму (двулопастную) и обычно имеет низкий или средний уровень поглощения ().
ПЭТ-сканирование с ФДГ (коронарный вид) у молодого взрослого. Нормальный тимус выглядит как перевернутая V-область умеренного поглощения (стрелки) [Изображение любезно предоставлено Центром молекулярной визуализации, Онкологический центр Питера МакКаллума, Восточный Мельбурн, Виктория, Австралия].
Отсутствие поглощения при сканировании перед началом терапии должно быть индикатором гиперплазии тимуса после лечения.Накопление ФДГ в тимусе указывает на патологию, когда он не имеет типичной треугольной формы или если активность очень интенсивна [9,10]. Железистая ткань молочной железы может демонстрировать умеренное поглощение ФДГ и относительно повышенное у субъектов в пременопаузе и у субъектов в постменопаузе, принимающих заместительную гормональную терапию (). Заметное поглощение можно увидеть в кормящих грудях.
Заметное поглощение можно увидеть в кормящих грудях.
ПЭТ-сканирование с ФДГ (коронарный вид) у молодой самки. Наблюдается двусторонний симметричный умеренный захват индикатора в груди [Изображение любезно предоставлено Центром молекулярной визуализации, Онкологический центр Питера МакКаллума, Восточный Мельбурн, Виктория, Австралия].
Сердечная активность варьируется от отсутствия заметной активности выше фоновой активности пула до интенсивной активности в миокарде левого желудочка. Активность предсердия или правого желудочка не наблюдается, если только эти камеры не поражены сердечным заболеванием. В состоянии голодания, когда уровень инсулина низкий, поглощение ФДГ сердечной мышцей должно быть низким. Поглощение миокардом повышается при высоком уровне глюкозы в крови, поэтому сердечная активность отмечается в постпрандиальном состоянии.В голодном состоянии обычно отмечается небольшая активность миокарда, поскольку миокард преимущественно использует жирные кислоты для выработки энергии. Однако потребление может быть различным даже в состоянии голодания. Мы можем видеть активность в аорте и магистральных сосудах, особенно при атеросклеротическом заболевании.
В мочевыводящих путях ФДГ фильтруется клубочками и не абсорбируется почечными канальцами. Таким образом, значительная активность может проявляться в любой части мочевыводящих путей или хирургических отведениях мочи, таких как подвздошные каналы.Печень, селезенка и костный мозг обычно демонстрируют однородное низкосортное поглощение. Костный мозг и селезенка обычно поглощают менее интенсивно, чем печень. Однако у пациентов, получающих фактор, стимулирующий рост колоний (GCSF), повышенная активность костного мозга и селезенки показана на сканировании FDG PET ().
ПЭТ-сканирование с FDG (коронарное изображение) у пациента, получавшего фактор, стимулирующий рост колоний (GCSF). Наблюдается повышенная активность костного мозга в позвоночнике и тазу и повышенная активность селезенки [Изображение любезно предоставлено Центром молекулярной визуализации, Онкологический центр Питера МакКаллума, Восточный Мельбурн, Виктория, Австралия].
Поглощение в желудочно-кишечном тракте может сильно варьироваться. Пищевод обычно не проявляет значительной активности. Однако желудочно-пищеводный переход может показывать нормальное поглощение. Гомогенное низкое поглощение стенкой желудка является относительно обычным явлением. Если желудок сокращен, это может выглядеть как круглая очаговая область умеренной активности. Поглощение в тонком кишечнике непостоянно и обычно низкого уровня. Активность толстой кишки может быть весьма выраженной, особенно в области слепой кишки и ректосигмовидного перехода.Как правило, наиболее высокий уровень поглощения наблюдается в толстой кишке, за которой следует тонкий кишечник, а в желудке наблюдается наименьшая интенсивность поглощения. Поглощение кишечником может быть диффузным, но не очаговым.
Захват яичками у самцов обычно наблюдается и симметрично диффузный. Однако у женщин захват яичниками обычно не наблюдается. Если поглощение яичниками наблюдается у пациентки в постменопаузе, следует исключить злокачественное новообразование. Слабая активность матки является обычным явлением. Сообщалось о проникновении в матку во время менструации и овуляции у женщин в пременопаузе и в отношении миомы, но на практике это редкость ().
ФДГ ПЭТ КТ (аксиальный вид) таза. Видна активность в полости эндометрия у этой менструирующей пациентки (стрелка). На совмещенном изображении матка видна ретровертированной [Изображение любезно предоставлено Центром молекулярной визуализации, Онкологический центр Питера МакКаллума, Восточный Мельбурн, Виктория, Австралия].
УПРАВЛЕНИЕ ФДГ В МЯГКИХ ТКАНЯХ
Общая область ошибок интерпретации связана с захватом ФДГ активными скелетными мышцами. У расслабленных и отдохнувших пациентов не наблюдается значительного поглощения скелетными мышцами.Мышечный дисбаланс, например, послеоперационный сколиоз, может привести к увеличению поглощения ФДГ пораженными мышцами (). Большую часть активности скелетных мышц можно легко распознать как таковую.
ПЭТ-сканирование с ФДГ (вид с коронки). Наблюдается повышенная активность мышц шеи и плеч. Умеренное поглощение также наблюдается в поясничных мышцах у этого пациента со сколиозом (стрелки) [Изображение любезно предоставлено Центром молекулярной визуализации, Онкологический центр Питера МакКаллума, Восточный Мельбурн, Виктория, Австралия].
Заметное поглощение индикаторов также было описано в надключичной жировой клетчатке при сканировании FDG PET примерно у 2% — 4% пациентов.Этиология не совсем понятна, но считается, что она связана с присутствием «бурого жира» (коричневой жировой ткани). Бурый жир наиболее заметен у новорожденных, и его количество с возрастом уменьшается. В отличие от белой жировой ткани, она способна выделять тепло. Его стимулируют несколько факторов, в том числе переохлаждение. Частота попадания индикаторов в бурый жир также увеличивается у женщин. Области, в которых наблюдается заметное поглощение метки коричневым жиром, находятся в надключичных областях, за которыми следуют подмышечные впадины, средостение, межреберные, паравертебральные и перинефрические области ().Даже если он распознан как доброкачественный вариант, степень поглощения может легко скрыть злокачественную лимфаденопатию в этом регионе. С помощью ПЭТ-КТ теперь легче дифференцировать жировую ткань и патологическую ткань () [11].
ПЭТ-сканирование с ФДГ (коронарный вид). Интенсивное поглощение индикаторов наблюдается в буром жире, наблюдаемом в надключичной, межреберной и паравертебральной областях [Изображение любезно предоставлено Центром молекулярной визуализации, Онкологический центр Питера МакКаллума, Восточный Мельбурн, Виктория, Австралия].
Слева направо — аксиальное изображение КТ, изображение ПЭТ и объединенное изображение ПЭТ КТ на одном уровне, показывающие области интенсивного поглощения жира в надключичных областях (стрелки).Объединенные изображения ПЭТ / КТ способствовали различению жировой ткани и патологической ткани [Изображение любезно предоставлено Центром молекулярной визуализации, Онкологический центр Питера МакКаллума, Восточный Мельбурн, Виктория, Австралия].
Важно знать о клинической корреляции других участков поглощения или отсутствия поглощения, что может не иметь значения. К ним относятся заживляющие переломы (менее трех месяцев) и заживление участков хирургических разрезов, участки предыдущей лучевой терапии (без или с низким уровнем облучения), суставные протезы (не инфицированные), дегенеративное заболевание суставов, участки стомы (колостома, илеостома, трахеостомия), грудная клетка. место дренажа трубки, места биопсии и места порта-катетера.
ПОДГОТОВКА ПАЦИЕНТА И ДРУГИЕ ТЕХНИЧЕСКИЕ ВОПРОСЫ
Надлежащая подготовка пациента необходима для сведения к минимуму появления потенциальных артефактных паттернов поглощения, затрудняющих интерпретацию. В день сканирования следует избегать упражнений, чтобы избежать поглощения мышцами. Пациенты должны быть полностью расслаблены и согреты. Если пациенту холодно и нервно, сжимание рук увеличит мышечную активность предплечий, что приведет к увеличению поглощения метки. Вызванное стрессом мышечное напряжение часто наблюдается в трапециевидных и параспинальных мышцах.Тем, у кого есть мышечные спазмы, можно вводить бензодиазепины перед инъекцией ФДГ. Это сводит к минимуму поглощение нормальными скелетными мышцами, особенно проксимальными мышцами, что затрудняет оценку шеи и надключичных узлов. Пациентам рекомендуется избегать чрезмерного разговора, жевания и глотания, чтобы уменьшить накопление метки в жевательных мышцах и гортани. Бурый жир обладает способностью выделять тепло. Известно, что потребление глюкозы увеличивается, когда симпатическая нервная система активируется холодовой стимуляцией.Следовательно, согревание пациента может помочь снизить поглощение бурого жира.
Пациентов готовят к сканированию ПЭТ-КТ так же, как и к сканированию ПЭТ. КТ выполняется перед ПЭТ. По онкологическим показаниям обычно проводится расширенное обследование тела, обычно включающее изображения от основания черепа до проксимальных отделов бедер. Типичное время сбора данных для компьютерной томографии составляет менее минуты. Компонент КТ выполняется как неконтрастное сканирование с низкой дозой облучения.Это выполняется в первую очередь для коррекции затухания и анатомической корреляции. Затем, без движения или изменения положения пациента, выполняется ПЭТ-сканирование, охватывающее то же поле изображения. Полученные в результате исследования ПЭТ-КТ интерпретируются на компьютерной рабочей станции со специальным программным обеспечением, позволяющим просматривать изображения ПЭТ, КТ и термоядерной ПЭТ с поправкой на затухание в трех ортогональных плоскостях. И КТ, и ПЭТ КТ изображения слияния используются для локализации аномалий поглощения ПЭТ. Многие из проблем, связанных с ловушками и областями нормального восприятия, описанными выше, решаются с использованием ПЭТ-КТ изображений.ПЭТ-КТ уменьшит диагностическую неопределенность в отношении физиологической активности, позволяя более уверенно интерпретировать области анатомически нормальных структур. Он объединяет анатомические и молекулярные данные с целью создания единого интегрированного диагноза.
Компонент КТ выполняется как неконтрастное сканирование с низкой дозой облучения.Это выполняется в первую очередь для коррекции затухания и анатомической корреляции. Затем, без движения или изменения положения пациента, выполняется ПЭТ-сканирование, охватывающее то же поле изображения. Полученные в результате исследования ПЭТ-КТ интерпретируются на компьютерной рабочей станции со специальным программным обеспечением, позволяющим просматривать изображения ПЭТ, КТ и термоядерной ПЭТ с поправкой на затухание в трех ортогональных плоскостях. И КТ, и ПЭТ КТ изображения слияния используются для локализации аномалий поглощения ПЭТ. Многие из проблем, связанных с ловушками и областями нормального восприятия, описанными выше, решаются с использованием ПЭТ-КТ изображений.ПЭТ-КТ уменьшит диагностическую неопределенность в отношении физиологической активности, позволяя более уверенно интерпретировать области анатомически нормальных структур. Он объединяет анатомические и молекулярные данные с целью создания единого интегрированного диагноза.
Разработка комбинированных ПЭТ-КТ-сканеров сопряжена со своими специфическими проблемами, связанными с комбинированной ПЭТ-КТ техникой [8]. Различия в паттернах дыхания при КТ и ПЭТ могут привести к неправильной регистрации, например.g., легочный узелок, расположенный на периферии и в основании легких. Ошибку регистрации можно свести к минимуму, выполняя КТ во время выдоха вместо вдоха. Вещества с высокой плотностью, например контрастные вещества или металлические предметы, могут привести к искусственно завышенной оценке активности, если данные КТ используются для коррекции затухания. Артефакты можно распознать, изучив нескорректированные данные изображения. Использование внутривенного контраста во время КТ может привести к чрезмерной коррекции артефактических горячих областей ослабления на изображении с коррекцией ослабления и количественной переоценке активности / поглощения ФДГ.Отдельное КТ-исследование с контрастным усилением показано, если использование внутривенного контраста необходимо для повышения точности диагностики.
Вкратце;
ФДГ локализуется в любой части тела, где наблюдается высокая физиологическая активность.
Основой для правильной интерпретации изображений ПЭТ-КТ является четкое понимание нормальных вариантов восприятия и осведомленность о доброкачественных процессах, которые активно используются ФДГ, чтобы избежать потенциальных ловушек при интерпретации изображений.
ССЫЛКИ
1. Хьюмс С. Онкологический ПЭТ / КТ: учебник для радиологов [веб-страница] Доступно на www.auntminnie.com.2. Повар Г.Дж., Фогельман И., Мейси М.Н. Нормальные физиологические и доброкачественные патологические варианты сканирования с позитронно-эмиссионной томографией 18-фтор-2-дезоксиглюкозы: возможность ошибки в интерпретации. Semin Nucl Med. 1996. 26 (4): 308–14. [PubMed] [Google Scholar] 3. Кук Г.Дж., Мейси М.Н., Фогельман И. Нормальные варианты, артефакты и ошибки интерпретации в ПЭТ-визуализации с 18-фтор-2-дезоксиглюкозой и углерод-11-метионином.Eur J Nucl Med. 1999. 26 (10): 1363–78. [PubMed] [Google Scholar] 4. Шрив П.П., Анзай Й., Валь Р.Л. Подводные камни в онкологической диагностике с помощью ФДГ-ПЭТ: физиологические и доброкачественные варианты. Рентгенография. 1999. 19 (1): 61–77. викторина 150-1. [PubMed] [Google Scholar] 5. Нгуен М., Варма В., Перес Р., Шустер Д.М. КТ с гистопатологической корреляцией поглощения ФДГ у пациента с легочной гранулемой и плевральной бляшкой, вызванной удаленным тальковым плевродезом. AJR Am J Roentgenol. 2004. 182 (1): 92–4. [PubMed] [Google Scholar] 6.Мюррей JG, Эразмус JJ, Bahtiarian EA, Goodman PC. Тальковый плевродез, моделирующий метастазы в плевре, на позитронно-эмиссионной томографии с 18F-фтордезоксиглюкозой. AJR Am J Roentgenol. 1997. 168 (2): 359–60. [PubMed] [Google Scholar] 7. Коэн М.С., Арслан Н., Дехдашти Ф. и др. Риск злокачественного новообразования при инциденталомах щитовидной железы, выявленных с помощью фтордезоксиглюкозо-позитронно-эмиссионной томографии. Операция. 2001. 130 (6): 941–6. [PubMed] [Google Scholar] 8. Кук Г.Дж., Вегнер Е.А., Фогельман И. Ловушки и артефакты при онкологической визуализации 18FDG PET и PET / CT.Semin Nucl Med. 2004. 34 (2): 122–33. [PubMed] [Google Scholar] 9. Бринк I, Райнхардт MJ, Hoegerle S, Altehoefer C, Moser E, Nitzsche EU. Повышенная метаболическая активность вилочковой железы, изученная с помощью ПЭТ с 18F-FDG: возрастная зависимость и частота после химиотерапии. J Nucl Med. 2001. 42 (4): 591–5. [PubMed] [Google Scholar] 10. Фердинанд Б., Гупта П., Крамер Э. Спектр поглощения тимуса при ПЭТ 18F-ФДГ. Рентгенография. 2004. 24 (6): 1611–6. [PubMed] [Google Scholar]
2001. 130 (6): 941–6. [PubMed] [Google Scholar] 8. Кук Г.Дж., Вегнер Е.А., Фогельман И. Ловушки и артефакты при онкологической визуализации 18FDG PET и PET / CT.Semin Nucl Med. 2004. 34 (2): 122–33. [PubMed] [Google Scholar] 9. Бринк I, Райнхардт MJ, Hoegerle S, Altehoefer C, Moser E, Nitzsche EU. Повышенная метаболическая активность вилочковой железы, изученная с помощью ПЭТ с 18F-FDG: возрастная зависимость и частота после химиотерапии. J Nucl Med. 2001. 42 (4): 591–5. [PubMed] [Google Scholar] 10. Фердинанд Б., Гупта П., Крамер Э. Спектр поглощения тимуса при ПЭТ 18F-ФДГ. Рентгенография. 2004. 24 (6): 1611–6. [PubMed] [Google Scholar]Физиологическая активность в спокойных тепловых помещениях
Общие шкалы теплового комфорта
Экспериментатор контролировал и поддерживал на одном уровне комнатную температуру и относительную влажность.Для подтверждения условия мы провели статистический анализ данных, измеренных в начале экспериментов. Не было значительной разницы в температуре в помещении между средой с кондиционированием воздуха (AC) и средой системы лучистого охлаждения и обогрева (RS) (охлаждение: Z = -1,4, p = 0,16, обогрев: Z = 0,67, p = 0,50 по Тест Вилкоксона) (рис.2, левые панели) и отсутствие значимых различий в относительной влажности (охлаждение: Z = -1,9, p = 0,056, нагрев: Z = -0,088, p = 0.93) (рис.2, средние панели).
Мы также сравнили скорости воздуха в экспериментальной комнате (рис. 2, правые панели). Были значительные различия в скорости воздуха между условиями AC и RS. Медиана (межквартильный диапазон) скорости воздуха в эксперименте с охлаждением составляла 1,1 (0,46) м / с в условиях переменного тока и 0,0033 (0,22) м / с в условиях RS (Z = -4,2, p <0,001, критерий Вилкоксона). (Рис. 2, вверху справа). Средняя скорость воздуха в эксперименте по нагреву составляла 0,61 (0,18) м / с в условиях переменного тока и 0.071 (0,053) м / с в состоянии RS (Z = -4,1, p <0,001) (рис.2, справа внизу).
Для сравнения результатов ЭЭГ с субъективными оценками теплового комфорта и ощущений мы использовали две тепловые шкалы: шкалы PMV и прогнозируемого процента неудовлетворенных (PPD) 5, 7 (рис. 3). PMV используется для выражения теплового комфорта как суммы баллов, основанных на нескольких типах переменных окружающей среды, скорости метаболизма участника и уровне теплоизоляции одежды 6 .Семибалльная шкала тепловых ощущений Американского общества инженеров по отоплению, охлаждению и кондиционированию воздуха (ASHARE) определяется PMV (−3: холод, −2: холод, −1: слегка прохладно, 0: нейтральный, +1 : слегка теплый, +2: теплый и +3: горячий). Зона теплового комфорта, указанная в стандарте ASHARE Standard 55 5, 7 , находится в диапазоне от -0,5 до +0,5.
3). PMV используется для выражения теплового комфорта как суммы баллов, основанных на нескольких типах переменных окружающей среды, скорости метаболизма участника и уровне теплоизоляции одежды 6 .Семибалльная шкала тепловых ощущений Американского общества инженеров по отоплению, охлаждению и кондиционированию воздуха (ASHARE) определяется PMV (−3: холод, −2: холод, −1: слегка прохладно, 0: нейтральный, +1 : слегка теплый, +2: теплый и +3: горячий). Зона теплового комфорта, указанная в стандарте ASHARE Standard 55 5, 7 , находится в диапазоне от -0,5 до +0,5.
Значения PMV и PPD, полученные в результате коллапса между сессиями в эксперименте с охлаждением (верхние панели) и эксперименте по нагреванию (нижние панели).Звездочками отмечены достоверные различия (*** p <0,001). Серые области на левой панели обозначают зону теплового комфорта, определенную стандартом ASHARE.
В эксперименте с охлаждением медиана (межквартильный размах) PMV в условиях переменного тока (с воздушным потоком) составляла -2,0 (0,44), а в условиях RS (без воздушного потока) составляла -0,10 (0,29). Эти значения значительно различались (Z = 3,5, p <0,001, критерий Вилкоксона) (рис. 3, вверху слева). PPD, который измеряет отношение количества людей, которые чувствуют себя неудовлетворенными, к родительской популяции, также значительно отличался в условиях с потоком воздуха и без него (медиана (межквартильный размах)) PPD: 6.0 (4,2)% в RS, 77 (15)% в AC, Z = -3,5, p <0,001) (рис. 3, вверху справа). В эксперименте по нагреванию медиана PMV в состоянии RS значительно отличалась от таковой в состоянии AC (0,23 (0,23) и 0,84 (0,43) соответственно; Z = -3,8, p <0,001) (рис. 3, внизу слева ). PPD также значительно различалась в двух условиях воздушного потока (медиана (межквартильный размах)) в AC = 23 (17)%, медиана (межквартильный размах) в RS = 6,2 (1,8)%; Z = −4,0, p <0,001) (рис. 3, внизу справа). Средние значения PMV и их межквартильные диапазоны в состоянии RS находились в пределах комфортного диапазона (от -0.От 5 до 0,5) как в экспериментах по охлаждению, так и по нагреву.
Средние значения PMV и их межквартильные диапазоны в состоянии RS находились в пределах комфортного диапазона (от -0.От 5 до 0,5) как в экспериментах по охлаждению, так и по нагреву.
Нейрофизиологическая активность
Чтобы сравнить условия с потоком воздуха и без него, для каждой полосы частот были рассчитаны изменения амплитуд ЭЭГ в каждом сеансе относительно первого сеанса. Эти полосы включают тета (4–8 Гц), альфа (8–14 Гц), бета (14–30 Гц) и гамма (30–55 Гц) колебания. Расположение записывающих электродов было выбрано таким образом, чтобы можно было измерить ответы, основанные как на психическом состоянии, так и на ощущениях (см. Методы, анализ ЭЭГ).
Амплитуды гамма-диапазона на височных и теменных участках (Pz, T3 и T4) имели меньшие относительные изменения в состоянии RS, чем в состоянии AC в обоих экспериментах (рис. 4). Двусторонний дисперсионный анализ (ANOVA) показал значительное влияние воздушного потока на амплитуды гамма-диапазона на каждом фокусном электроде в каждом эксперименте (охлаждение, Pz: F (1,72) = 4,4, p = 0,040, T3: F ( 1,72) = 8,8, p = 0,0040, T4: F (1,72) = 9,1, p = 0,0036; нагревание, Pz: F (1,72) = 11, p = 0,0016, T3: F (1,72 ) = 20, p <0.001, T4: F (1,72) = 16, p <0,001). Основной эффект сеанса также был значимым или незначительно значимым (охлаждение, Pz: F (4,72) = 5,4, p <0,001, T3: F (4,72) = 2,9, p = 0,028, T4: F (4, 72) = 5,0, p = 0,0013; нагревание, Pz: F (4,72) = 2,2, p = 0,076, T3: F (4,72) = 4,2, p = 0,0040, T4: F (4,72) = 7.1, р <0,001). Не было значительного взаимодействия между условиями воздушного потока и сеансами (охлаждение, Pz: F (4,72) = 1,5, p = 0,20, T3: F (4,72) = 0,85, p = 0,50, T4: F (4,72) ) = 0,88, p = 0,48; нагрев, Pz: F (4,72) = 0.83, p = 0,51, T3: F (4,72) = 1,4, p = 0,24, T4: F (4,72) = 1,3, p = 0,30). Мы подтвердили, что значение PMV и гамма-амплитуды коррелировали в обоих экспериментах (охлаждение: r = -0,24, p = 0,031, нагрев: r = 0,36, p = 0,0031 при T3) (см. Рисунки S1a и S2a в дополнительной информации). Анализ гамма и PPD также подтвердил незначительную корреляцию с охлаждением (r = 0,21, p = 0,065) и значительную корреляцию с нагревом (r = 0,31, p = 0,013 при T3) (рис. S3a и S4a в дополнительной информации).Хотя эти результаты подтверждают взаимосвязь между гаммой и PMV или PPD, аналитические данные измерялись неоднократно. Чтобы решить эту проблему, мы проанализировали корреляции, используя данные, усредненные по сеансам. Однако каких-либо существенных корреляций обнаружить не удалось. Это может быть из-за небольшой степени свободы (рисунки S5a – S8a и таблицы S1 – S4 в дополнительной информации). Мы ввели новый индекс «двухточечного наклона», чтобы исследовать взаимосвязь между амплитудами ЭЭГ и PMV или PPD в каждом сеансе (см. Дополнительные методы).Медианные значения двухточечных наклонов гаммы, которые значительно отличались от нуля, потенциально указывали на наличие некоторой взаимосвязи выше уровня вероятности между PPD и EEG в качестве независимых измерений. Двухточечные наклоны, рассчитанные на основе гамма и PMV, имели значимые отличия от нуля на сеансе 2 и незначительно значимые различия на сеансе 3 при охлаждении (сеанс 2: Z = -16, p = 0,037; сеанс 3: Z = -14, p = 0,065; рис. S9a и таблица S5 в дополнительной информации), как определено с помощью одностороннего теста Вилкоксона (односторонний).При нагревании наблюдались значимые отличия от нуля на сеансе 4 и предельные значения на сеансах 2 и 5 (сеанс 2: Z = 12, p = 0,055; сеанс 4: Z = 14, p = 0,027; сеанс 5, Z = 12, p = 0,055; рис. S11a и таблица S7 в дополнительной информации). Мы также оценили двухточечные наклоны между гамма и PPD и обнаружили существенные различия и предельные значения в охлаждении (сеанс 2: Z = 16, p = 0,037; сеанс 3: Z = 14, p = 0,065; рис. S10a и таблица S6. в дополнительной информации) и нагревание (сеанс 2: Z = 12, p = 0.055; сеанс 3: Z = 10, p = 0,098; сессия 4: Z = 14, p = 0,027; сеанс 5: Z = 12, p = 0,055; Рис. S12a и таблица S8 в дополнительной информации). Эти результаты показывают, что более высокие гамма-амплитуды могут быть факторами-кандидатами для объяснения субъективных оценок, сделанных с использованием PMV и PPD.
Анализ гамма и PPD также подтвердил незначительную корреляцию с охлаждением (r = 0,21, p = 0,065) и значительную корреляцию с нагревом (r = 0,31, p = 0,013 при T3) (рис. S3a и S4a в дополнительной информации).Хотя эти результаты подтверждают взаимосвязь между гаммой и PMV или PPD, аналитические данные измерялись неоднократно. Чтобы решить эту проблему, мы проанализировали корреляции, используя данные, усредненные по сеансам. Однако каких-либо существенных корреляций обнаружить не удалось. Это может быть из-за небольшой степени свободы (рисунки S5a – S8a и таблицы S1 – S4 в дополнительной информации). Мы ввели новый индекс «двухточечного наклона», чтобы исследовать взаимосвязь между амплитудами ЭЭГ и PMV или PPD в каждом сеансе (см. Дополнительные методы).Медианные значения двухточечных наклонов гаммы, которые значительно отличались от нуля, потенциально указывали на наличие некоторой взаимосвязи выше уровня вероятности между PPD и EEG в качестве независимых измерений. Двухточечные наклоны, рассчитанные на основе гамма и PMV, имели значимые отличия от нуля на сеансе 2 и незначительно значимые различия на сеансе 3 при охлаждении (сеанс 2: Z = -16, p = 0,037; сеанс 3: Z = -14, p = 0,065; рис. S9a и таблица S5 в дополнительной информации), как определено с помощью одностороннего теста Вилкоксона (односторонний).При нагревании наблюдались значимые отличия от нуля на сеансе 4 и предельные значения на сеансах 2 и 5 (сеанс 2: Z = 12, p = 0,055; сеанс 4: Z = 14, p = 0,027; сеанс 5, Z = 12, p = 0,055; рис. S11a и таблица S7 в дополнительной информации). Мы также оценили двухточечные наклоны между гамма и PPD и обнаружили существенные различия и предельные значения в охлаждении (сеанс 2: Z = 16, p = 0,037; сеанс 3: Z = 14, p = 0,065; рис. S10a и таблица S6. в дополнительной информации) и нагревание (сеанс 2: Z = 12, p = 0.055; сеанс 3: Z = 10, p = 0,098; сессия 4: Z = 14, p = 0,027; сеанс 5: Z = 12, p = 0,055; Рис. S12a и таблица S8 в дополнительной информации). Эти результаты показывают, что более высокие гамма-амплитуды могут быть факторами-кандидатами для объяснения субъективных оценок, сделанных с использованием PMV и PPD.
Дифференциальные гамма-амплитуды (среднее ± SEM) из первого сеанса.
Амплитуды бета-полосы в центральных точках (C3 и C4) также имели меньшие относительные изменения в состоянии RS, чем в условиях переменного тока, как в экспериментах по нагреванию, так и по охлаждению (рис.5). ANOVA показал значительное влияние воздушного потока на амплитуды бета-диапазона (охлаждение, C3: F (1,72) = 5,0, p = 0,030, C4: F (1,72) = 6,0, p = 0,017; нагрев, C3: F (1,72) = 10, p = 0,023, C4: F (1,72) = 8,5, p = 0,0048). Основные эффекты сеанса также были обнаружены в C3 и C4 в эксперименте с охлаждением (C3: F (4,72) = 3,1, p = 0,022, C4: F (4,72) = 4,3, p = 0,0036) и в Сайт C4 в эксперименте по нагреванию (C3: F (4,72) = 1,8, p = 0,15, C4: F (4,72) = 3,1, p = 0,021). Однако не было значительного взаимодействия между состоянием воздушного потока и сеансом (охлаждение, C3: F (4,72) = 0.33, p = 0,86, C4: F (4,72) = 0,95, p = 0,44; нагревание, C3: F (4,72) = 0,71, p = 0,59, C4: F (4,72) = 1,0, p = 0,41). Корреляционный анализ между PMV и бета-амплитудами показал значительную корреляцию в охлаждении (r = -0,23, p = 0,041 в C3) (см. Рис. S1b в дополнительной информации) и незначительную значимую корреляцию в эксперименте по нагреванию (r = 0,23, p = 0,062 в C3) (см. Рис. S2b в дополнительной информации). Анализы бета и PPD также подтвердили значительную корреляцию в охлаждении (r = 0.22, p = 0,049 для C3), но не при нагревании (r = 0,15, p = 0,24 для C3) (рис. S3b и S4b в дополнительной информации). Эти результаты предполагают, что бета-активность в некоторой степени коррелирует с субъективными ощущениями в состоянии охлаждения, тогда как в состоянии нагрева корреляции нет. Мы также выполнили анализ двухточечных наклонов, как мы это сделали для гамма-активности, чтобы более глубоко изучить взаимосвязь между PMV или PPD и бета (дополнительные методы). Двухточечные наклоны PMV не имели достоверных отличий от нуля ни в одном из сеансов в состоянии охлаждения (p ≥ 0. 10, рис. S9b и таблица S5 в дополнительной информации), хотя была незначительно значимая разница в сеансе 5 при нагревании (сеанс 5: Z = 10, p = 0,098, рис. S11b и таблица S7 в дополнительной информации), по оценке с помощью одностороннего теста Вилкоксона (односторонний). Кроме того, двухточечные наклоны PPD не имели значительных различий при охлаждении и нагревании (P ≥ 0,10, рис. S10b и таблица S6 в дополнительной информации), хотя на сеансе 5 при нагревании была незначительная разница (сеанс 5: Z = 10, p = 0.098, рис. S12b и таблица S8 в дополнительной информации). Анализ двухточечных наклонов не дал нам никаких доказательств, подтверждающих взаимосвязь между бета- и температурными масштабами, PMV или PPD.
10, рис. S9b и таблица S5 в дополнительной информации), хотя была незначительно значимая разница в сеансе 5 при нагревании (сеанс 5: Z = 10, p = 0,098, рис. S11b и таблица S7 в дополнительной информации), по оценке с помощью одностороннего теста Вилкоксона (односторонний). Кроме того, двухточечные наклоны PPD не имели значительных различий при охлаждении и нагревании (P ≥ 0,10, рис. S10b и таблица S6 в дополнительной информации), хотя на сеансе 5 при нагревании была незначительная разница (сеанс 5: Z = 10, p = 0.098, рис. S12b и таблица S8 в дополнительной информации). Анализ двухточечных наклонов не дал нам никаких доказательств, подтверждающих взаимосвязь между бета- и температурными масштабами, PMV или PPD.
Дифференциальные бета-амплитуды (среднее ± стандартная ошибка среднего) из первого сеанса.
Амплитуды альфа-полосы имели разную картину в экспериментах по охлаждению и нагреванию (рис. 6). В эксперименте с охлаждением относительные изменения в альфа-диапазоне не указывали на основной эффект воздушного потока (C3: F (1,72) = 0.67, p = 0,42, C4: F (1,72) = 0,42, p = 0,52). Однако основной эффект воздушного потока наблюдался в эксперименте по нагреванию (C3: F (1,72) = 13, p <0,001, C4: F (1,72) = 7,4, p = 0,0080). Кроме того, основной эффект сеанса наблюдался в эксперименте с охлаждением (C3: F (4,72) = 3,8, p = 0,0079, C4: F (4,72) = 3,5, p = 0,011), но не в эксперименте с охлаждением. нагревательный эксперимент (C3: F (4,72) = 1,0, p = 0,40, C4: F (4,72) = 1,1, p = 0,37). Никаких взаимодействий между этими факторами обнаружено не было (охлаждение, C3: F (4,72) = 0,54, p = 0,71, C4: F (4,72) = 0.46, р = 0,76; нагревание, C3: F (4,72) = 1,1, p = 0,36, C4: F (4,72) = 0,80, p = 0,53).
Рисунок 6Дифференциальные альфа-амплитуды (среднее ± стандартная ошибка среднего) из первого сеанса.
Также были проанализированы амплитуды тета-диапазона, но не было обнаружено основного эффекта воздушного потока (охлаждение, Fz: F (1,72) = 1,3, p = 0,25; нагрев, Fz: F (1,72) = 0,081, p = 0,15). ). Основной эффект сеанса был обнаружен как в экспериментах по нагреванию, так и по охлаждению (охлаждение, Fz: F (4,72) = 9,3, p <0.001, нагрев, Fz: F (4,72) = 3,2, p = 0,019), но не было обнаружено значительного взаимодействия между потоком воздуха и сеансом (охлаждение, Fz: F (4,72) = 0,20, p = 0,93, нагрев, Fz: F (4,72) = 0,29, p = 0,89).
). Основной эффект сеанса был обнаружен как в экспериментах по нагреванию, так и по охлаждению (охлаждение, Fz: F (4,72) = 9,3, p <0.001, нагрев, Fz: F (4,72) = 3,2, p = 0,019), но не было обнаружено значительного взаимодействия между потоком воздуха и сеансом (охлаждение, Fz: F (4,72) = 0,20, p = 0,93, нагрев, Fz: F (4,72) = 0,29, p = 0,89).
Вкратце, гамма- и бета-амплитуды указывают на основные эффекты в условиях воздушного потока как в охлаждающей, так и в нагревающей среде. Таким образом, гамма- и бета-активности могут отражать ощущение самого воздушного потока, поскольку различия наблюдались в обоих экспериментах независимо от сезона. Напротив, альфа-амплитуды указали на основной эффект воздушного потока только в эксперименте по нагреванию.Следовательно, на характер альфа-активности могут влиять различия в тепловых ощущениях. Амплитуды тета-диапазона не указывают на какие-либо основные эффекты условий воздушного потока в интересующих местах расположения электродов.
Задача с точностью до времени
Мы проанализировали общую продолжительность (TTL, приблизительно 60 секунд) периода задачи для каждого участника. TTL в условиях RS было больше, чем в среде переменного тока, как в экспериментах по охлаждению, так и по нагреву (рис. 7). Основное влияние воздушного потока было обнаружено на TTL как в охлаждающей, так и в нагревающей среде (охлаждение: F (1,70) = 10, p = 0.0027, нагревание: F (1,56) = 4,4, p = 0,040). Эти результаты показывают, что, по оценкам участников, в обоих экспериментах время в условиях RS, в которых отсутствовал воздушный поток, было больше. Основного влияния сеанса на TTL не было (охлаждение: F (3,70) = 0,27, p = 0,85, нагревание: F (3,56) = 0,23, p = 0,87).
Рисунок 7Разница общей продолжительности времени (TTL, среднее ± SEM) от первого сеанса.
Данные термографии температуры кожи
Было обнаружено, что поток воздуха напрямую влияет на температуру кожи 15 . В начале и в конце каждого эксперимента температуру кожи каждого участника измеряли с помощью термографии (рис. 8).
В начале и в конце каждого эксперимента температуру кожи каждого участника измеряли с помощью термографии (рис. 8).
Температура кожи (среднее ± SEM) в эксперименте с охлаждением ( a ) и эксперименте с нагреванием ( d ), а также примеры термографии во время охлаждения ( b , c ) и нагревания ( e , f ) эксперименты.
В эксперименте с охлаждением измеренные температуры кожи в условиях переменного тока в конце эксперимента были ниже, чем в начале (t (22) = −6.3, p <0,001, двусторонний критерий Стьюдента t ). Напротив, температура кожи в состоянии RS не различалась между первым и последним сеансами (t (22) = 1,1, p = 0,30). В эксперименте по нагреванию измеренные температуры кожи в состоянии переменного тока на последнем сеансе были выше, чем на первом сеансе (t (16) = 5,9, p <0,001). Напротив, температура кожи в состоянии RS не различалась между первым и последним сеансами (t (16) = -0,24, p = 0,59). Эти результаты подтверждают, что условия воздушного потока влияют только на температуру поверхности тела.
Физиологическая активность и активные компоненты потенциально лекарственных растений, используемых дикими шимпанзе гор Махале, Танзания
Асака, Ю., Кубота, Т., и Кулкарни, А. Б. (1977). Исследования на горьком принципе из Vernonia anthelmintica . Фитохимия 16: 1838–1839.
Google Scholar
Беркилл, Х. М. (1985). Полезные растения Западной Тропической Африки (2-е изд.), т. 1 , Королевский ботанический сад, Кью.
Google Scholar
Далзил, Дж. М. (1937). Полезные растения западной тропической Африки. В Huchinson, J., and Dalziel, J. M. (eds.), Приложение к флоре Западной Тропической Африки , Whitefriars Press, Лондон.
Google Scholar
Дойл, Т. В., и Брэднер, В. Т. (1980). Трихотеканы. В Кэссиди, Дж.М. и Дурос, Дж. Д. (ред.), Противораковые агенты на основе моделей натуральных продуктов , Academic Press, Нью-Йорк, Лондон, Торонто, Сидней, Сан-Франциско, стр. 43–72.
В., и Брэднер, В. Т. (1980). Трихотеканы. В Кэссиди, Дж.М. и Дурос, Дж. Д. (ред.), Противораковые агенты на основе моделей натуральных продуктов , Academic Press, Нью-Йорк, Лондон, Торонто, Сидней, Сан-Франциско, стр. 43–72.
Google Scholar
Гянджян И., Кубо И. и Фудзински П. (1983). Лактоны элеманолида, противозачаточные насекомым, из Vernonia amygdalina . Фитохимия 22: 2525–2526.
Google Scholar
Гаске, м., Бамба, Д., Бабаджамян, А., Балансард, Г., Тимон-Дэвид, П., и Мецгер, Дж. (1985). Действие амебицида и антигельминтных веществ вернолида и гидроксивернолида, изолятов дезоксирибонуклеиновой кислоты Vernonia colorata (Willd.) Drake. Eur. J. Med. Chem. 20: 111–115.
Google Scholar
Хеккер, Э. (1963). Über die Wirkstoffe des Croton Ols, I. Biologische Teste zur Quantitativen Messung der entzundlichen cocarcinogenen und toxischen Wirkung. Z. Krebforsch. 65: 325–333.
Google Scholar
Хаффман, М. А., и Сейфу, М. (1989). Наблюдения за болезнью и потреблением возможно лекарственного растения Vernonia amygdalina (Del.) Диким шимпанзе в национальном парке Махале-Маунтинс, Танзания. Приматы 30: 51–63.
Google Scholar
Кокваро, Дж. О. (1976). Лекарственные растения Восточной Африки , General Printers, Найроби.
Google Scholar
Купчан С. М., Хемингуэй Р. Дж., Карим А. и Вернер Д. (1969). Ингибиторы опухолей. XLVII. Вернодалин и верномигдин, два новых цитотоксических сесквитерпеновых лактона из Vernonia amygdalina Del. J. Org. Chem. 34: 3908–3911.
Google Scholar
Мишель, Р. И., и Даттон Р. У. (1967). Иммунизация диссоциированных культур клеток селезенки нормальных мышей. J. Exp. Med. 126: 423–442.
И., и Даттон Р. У. (1967). Иммунизация диссоциированных культур клеток селезенки нормальных мышей. J. Exp. Med. 126: 423–442.
Google Scholar
Мурамацу, М., Ониши, Т., Макино, С., Хаякумо, Ю. и Фуджи, С. (1965). Подавление триптической активности различными синтетическими ингибиторами. J. Biochem. 58: 214–226.
Google Scholar
Нисида, Т.(1990). Четверть века исследований в горах Махале. In Nishida, T. (ed.), The Shimpanzees of Mahale Mountains Sexual and Life-History Strategies , University of Tokyo Press, Tokyo, pp. 3–35.
Google Scholar
Охигаши, Х., Такамура, Х., Кошимидзу, К., Токуда, Х. и Ито, Ю. (1986). Поиск возможных противоопухолевых промоторов путем ингибирования активации вируса Эпштейна-Барра, вызванной 12-O-тетрадеканоилфорбол-13-ацетатом; Урсоловая кислота и олеаноловая кислота из противовоспалительного китайского лекарственного растения, Glechoma hederacea L. Cancer Lett. 30: 143–151.
Google Scholar
Окуяма, Х. (1990). Биохимия питания незаменимых жирных кислот. Кагаку — Сейбуцу 28: 175–181 (на японском).
Google Scholar
Филлипс-Конрой, Дж. Э. (1986). Павианы, диета и болезни: выбор пищи и шистосомоз. В Taub, D. M., and King, F. A. (eds.), Current Perspectives in Primate Social Dynamics , Van Nostrand Reinhold, New York, pp.287–304.
Google Scholar
Поттер, Н. Н. (1968). Наука о продуктах питания , AVI, Вестпорт, Коннектикут, стр. 398–419.
Google Scholar
Родрикес, М., Арегуллин, М., Нишида, Т., Уэхара, С., Рэнгэм, Р., Абрамовски, З., Финлейсон, А. , и Тауэрс, Г. Х. Н. (1985). Тиарубурин А, биоактивный компонент Aspilia (Asteraceae), потребляемый дикими шимпанзе. Experientia 41: 419–420.
, и Тауэрс, Г. Х. Н. (1985). Тиарубурин А, биоактивный компонент Aspilia (Asteraceae), потребляемый дикими шимпанзе. Experientia 41: 419–420.
Google Scholar
Родригес, Э., Тауэрс, Г. Х. Н. и Митчелл, Дж. К. (1976). Биологическая активность сесквитерпеновых лактонов (обзор). Фитохимия 15: 1573–1580.
Google Scholar
Шиба Т. (1976). Химия горького и острого вкуса. Kagaku Sosetsu 14: 129–156 (на японском языке).
Google Scholar
Такасаки, Х.и Хант К. (1987). Дальнейшее потребление лекарственных растений дикими шимпанзе? Afr. Stud. Monogr. 8: 125–128.
Google Scholar
Токуда, Х., Охигаши, Х., Кошимидзу, К., Ито, Ю. (1986). Ингибирующее действие урсоловой и олеаноловой кислоты на продвижение опухоли кожи 12-O-тетрадеканоилфорбол-13-ацетата. Cancer Lett. 33: 279–285.
Google Scholar
Ватт, Дж.М., и Брейер-Брандвейк, М. Г. (1962). Лекарственные и ядовитые растения Южной и Восточной Африки . Э. и С. Ливингстон, Эдинбург и Лондон, стр. 296.
Google Scholar
Ваттенберг, Л. В. (1985). Химиопрофилактика рака. Cancer Res. 45: 1–8.
Google Scholar
Wrangham, R. W., and Goodall, J. (1989). Шимпанзе используют лечебные листья.В Heltne, P. G., and Murquardt, L.A. (ред.), Understanding Chimpanzees , Harvard University Press, Cambridge, Mass., Стр. 22–37.
Google Scholar
Wrangham, R. W., and Nishida, T. (1983). Aspilia spp. листья: загадка пищевого поведения диких шимпанзе. Приматы 24: 276–282.
Google Scholar
Восстановление физиологической активности тромбоцитов у новорожденных телят с дефицитом железа
Завалишина С.Ю. Восстановление физиологической активности тромбоцитов у новорожденных телят с дефицитом железа. Biomed Pharmacol J 2017; 10 (2).
Рукопись получена: 11 марта 2017 г.
Рукопись принята: 3 апреля 2017 г.
Опубликовано онлайн: —
Проверка на плагиат: Да
Как Citeclose | История публикаций Закрыть Просмотры: (Посещали 352 раза, Посещали сегодня 1) Загрузки в формате PDF: 458
Светлана Юрьевна Завалишина
Всероссийский научно-исследовательский институт физиологии, биохимии и питания животных, Институт села, Боровск, Россия.
DOI: https://dx.doi.org/10.13005/bpj/1160
Аннотация
Новорожденные телята все еще часто встречаются с дефицитом железа. Это состояние в определенной степени негативно влияет на их рост и развитие из-за развития тромбоцитопатии. В связи с этим большое научное и практическое значение для ветеринарии и физиологии имеет поиск подходов к эффективной коррекции тромбоцитопатии новорожденных телят в условиях недостатка железа.Представляется перспективным оценить степень влияния на активность тромбоцитов новорожденных телят традиционно применяемого при дефиците железа ферроглюкина в сочетании со стимуляторами метаболизма (полисоном и крезацином). В ходе наших исследований было установлено, что у новорожденных телят с дефицитом железа также наблюдается снижение антиоксидантной защиты плазмы, усиление процессов перекисного окисления липидов, повышение гемостатической активности тромбоцитов. В результате применения комбинации ферроглюкина, полисона и крезацина новорожденным теленкам с дефицитом железа удалось добиться заметного повышения антиоксидантной защищенности плазмы крови, значительного снижения ею процессов перекисного окисления липидов при нормализации активности тромбоцитов.
Ключевые слова
новорожденных телят; дефицит железа; система гемостаза; ферроглюкин; полисон; крезацин
Загрузите эту статью как:| Для цитирования этой статьи скопируйте следующее: Завалишина С. Ю. Восстановление физиологической активности тромбоцитов у новорожденных телят с дефицитом железа. Biomed Pharmacol J 2017; 10 (2). |
| Скопируйте следующее, чтобы процитировать этот URL: Завалишина С.Ю. Восстановление физиологической активности тромбоцитов у новорожденных телят с дефицитом железа. Biomed Pharmacol J 2017; 10 (2). Доступно по ссылке: http://biomedpharmajournal.org/?p=14568 |
Введение
Становится ясно, что фаза новорожденного крупного рогатого скота — очень важный этап развития для всего последующего онтогенеза. 1,2 Благополучие и плохие условия окружающей среды на этом этапе жизни могут серьезно повлиять на последующие фазы раннего онтогенеза, включая процессы реализации наследственной информации во время роста, развития и воспроизводства. 3,4 В то же время на многих российских фермах у новорожденных телят часто обнаруживаются различные отклонения, отрицательно влияющие на процессы обмена веществ и, наконец, на их рост и развитие. 5 В предыдущих исследованиях было установлено, что при отклонениях гомеостаза, особенно у молодого организма, может происходить быстрое повышение активности компонентов гемостаза, что может привести к нарушению микроциркуляции. 6,7 Наиболее многочисленные исследования в этой области проводились на людях. 8,9,10 По их результатам удалось составить представление о наличии возрастной динамики активности компонентов гемостаза, 11,12 наиболее уязвимых ее механизмов и возможности различных вариантов воздействия на организм с целью оптимизации процессов гемостаза. Ввиду большой социальной значимости тромбозов при кардиальной патологии 13,14 современные исследователи твердо удерживают внимание на аспектах гемостатических изменений, возникающих у данной категории пациентов. 15,16,17 Именно при изучении патогенеза гемостазиопатии при кардиальной патологии сформулировано представление о возможности ее коррекции не только обычными кардиологическими средствами 18,19,20 , но и показана возможность их минимизации с помощью традиционных приложений 21,22,23 , что действительно важно для биологических исследований.
Ввиду большой социальной значимости тромбозов при кардиальной патологии 13,14 современные исследователи твердо удерживают внимание на аспектах гемостатических изменений, возникающих у данной категории пациентов. 15,16,17 Именно при изучении патогенеза гемостазиопатии при кардиальной патологии сформулировано представление о возможности ее коррекции не только обычными кардиологическими средствами 18,19,20 , но и показана возможность их минимизации с помощью традиционных приложений 21,22,23 , что действительно важно для биологических исследований.
Поскольку состояние дефицита железа среди новорожденных телят достаточно распространено 1,5 и довольно часто сопровождается развитием нарушений в системе гемостаза 24 , существует большая практическая потребность в их быстром и эффективном устранении среди телят. на фермах.В то же время эффективные подходы, направленные на одновременное уменьшение признаков дефицита железа и гемостазиопатии, все еще остаются неудовлетворительными. 25
Поэтому исследования новорожденных телят с целью поиска подходов к ранней и эффективной коррекции гемостазиопатии на модели железодефицитного состояния сохраняют свое большое научное и практическое значение. Разработанные при данном состоянии варианты доказательств снижения нарушений гемостаза могут служить основой для последующего создания корректирующих комплексов, способных быть эффективными в области снижения гемостазиопатии новорожденных телят при многих заболеваниях.Большой интерес представляет оценка влияния на всю систему гемостаза комбинации традиционно применяемых при дефиците железа ферроглюкина 25 и ранее показавших их высокую биологическую активность и способность влиять на отдельные компоненты системы гемостаза, метаболически активные средства — полисон 26 и крезацин. 27
В связи с этим мы ставим перед собой следующую цель — найти доказательства коррекции активности тромбоцитов у новорожденных телят с дефицитом железа с помощью комбинации ферроглюкина, полисона и крезацина.
Материалы и методы
Работа выполнена с 37 новорожденными теленками с признаками эритрогенеза и снижением содержания железа в организме (сывороточное железо 13,1 ± 0,09 мкмоль / л, сидероциты 1,5 ± 0,05%, гемоглобин 98, 2 ± 0,25 г / л, эритроциты 4,2 ± 0,18 × 10 12 / л). Контрольную группу составили 29 здоровых новорожденных телят.
Состояние перекисного окисления липидов (ПОЛ) в плазме крови животных определяли по количеству в ней тиобарбитуровой кислоты — активных продуктов с помощью набора фирмы «Агат-Мед» (Россия) и ацилгидропероксидов с учет уровня антиоксидантной активности жидкой части крови. 28 Число тромбоцитов в крови телят определено их подсчетом в камере Горяева. Агрегацию тромбоцитов регистрировали визуальным микрометодом 29 с некоторыми индукторами: с АДФ (0,5 × 10 -4 М), с тромбином (0,125 ед / мл), с коллагеном (разведение 1: 2 основной суспензии). , с ристомицином (0,8 мг / мл), с адреналином (5 × 10 -6 М) в плазме со стандартизованным количеством тромбоцитов в ней (200 × 10 9 тр.).
Коррекция железодефицитного состояния новорожденных телят проводилась ферроглюкином внутримышечно, однократно из расчета 15 мг железа на 1 кг массы тела, полисоном 5 мг / кг утром в схеме жидкого кормления в течение 6 дней и крезацином. — ежедневно по 3 мг / кг в схеме жидкого кормления в течение 6 дней, начиная с одновременного применения ферроглюкина.Оценка состояния здоровых животных производилась дважды — при рождении и на 7 -е -е сутки жизни. Из-за отсутствия достоверных различий между результатами обоих исследований контрольные значения каждого показателя представлены одной цифрой — простым средним между ними. Обследование телят с дефицитом железа проводилось дважды — при рождении и на следующий день после окончания коррекции (7 -й -й день жизни). Статистическая обработка полученных данных проводилась по t-критерию Стьюдента.
Результаты
У обследованных новорожденных телят с дефицитом железа обнаружены характерные для данного состояния слабость, вялость, отсутствие интереса к окружающей среде, бледность риноскопа и слоев слизи. У этих животных отмечена повышенная активность ПОЛ в плазме (ацилгидропероксид 3,41 ± 0,022 D 233 /1 мл, активные продукты тиобарбитуровой кислоты 5,20 ± 0,027 мкмоль / л при снижении значения антиоксидантной активности жидкой части крови 22,2 ± 0,15%). Значения этих показателей в контроле составили 1,45 ± 0,010 D 233 /1 мл, 3,46 ± 0,012 мкмоль / л и 33,7 ± 0,15% соответственно.
Количество тромбоцитов в крови новорожденных телят соответствовало норме. Кроме того, у животных с дефицитом железа достоверно увеличилась агрегация тромбоцитов (таблица). Их самая ранняя агрегация тромбоцитов возникла в ответ на коллаген (19,2 ± 0,21 с), чуть позже — с АДФ и ристомицином, еще позже — в ответ на тромбин (36,5 ± 0,12 с). Последняя агрегация тромбоцитов у телят с дефицитом железа возникла под действием адреналина (67,9 ± 0,23 с).
Таблица 1: Параметры гемостаза у новорожденных телят с
| Считаем показатели | Телята с дефицитом железа, n = 37, M ± m | управление, | исход | после коррекции | n = 29, M ± m |
| агрегация тромбоцитов с АДФ, с | 26,0 ± 0,16 | 40,1 ± 0,12 | 40,2 ± 0,08 | р 1 <0,01 | р <0,01 |
| агрегация тромбоцитов с коллагеном, с | 19,2 ± 0,21 | 31,3 ± 0,08 | 31,4 ± 0,08 | р 1 <0,01 | р <0,01 |
| агрегация тромбоцитов с тромбином, с | 36,5 ± 0,12 | 54,2 ± 0,20 | 53,8 ± 0,07 | р 1 <0,01 | р <0,01 |
| агрегация тромбоцитов с кристалломицином, с | 21,0 ± 0,19 | 48,1 ± 0,14 | 48,0 ± 0,12 | р 1 <0,01 | р <0,01 |
| агрегация тромбоцитов с адреналином, с | 67,9 ± 0,23 | 97,4 ± 0,16 | 97,6 ± 0,06 | р 1 <0,01 | р <0,01 |
Условные обозначения: p — достоверность различий показателей между контролем и исходным состоянием телят с дефицитом железа, p 1 — достоверность динамики показателей у телят с дефицитом железа на фоне коррекции.
Проведенная коррекция состояния обеспечила у обследованных телят улучшение общего состояния и активности железодефицитных телят, повышение уровня сывороточного железа до контрольных значений (23,2 ± 0,21 мкмоль / л). На фоне комбинации ферроглюкина, полисона и крезацина у обследованных телят обнаружено выраженное снижение содержания в плазме ацилгидропероксидов (1,70 ± 0,014 D 233 /1 мл, p <0,05) и продуктов, действующих на тиобарбитуровую кислоту (3 , 87 ± 0,019 мкмоль / л, p <0,05) при повышении антиоксидантной активности до 28,6 ± 0,16% (p <0,05).
Проведение коррекции у животных, вначале имевших дефицит железа, сопровождалось неизменностью количества тромбоцитов в их крови и замедлением агрегации тромбоцитов до контрольного уровня. Кроме того, наиболее активно тромбоциты животных реагировали агрегацией на коллаген, АДФ и ристомицин, менее активно — на добавление в плазму тромбина и адреналина (таблица).
Обсуждение
Реализация генетически определенных процессов роста и развития живых организмов происходит при постоянном воздействии на организм множества факторов внешней и внутренней среды. 4,30 Физиологические особенности их влияния в большей степени выражаются оптимумом кровоснабжения живых существ 31,32 , особенно в части деятельности компонентов системы гемостаза. 33,34 Кроме того, любые нарушения в организме сопровождаются отрицательной динамикой гематологических показателей 9,35 , включая параметры системы гемостаза. 34,36 Выясняется, что в основе развития гемостазиопатии у обследованных новорожденных телят не только дефицит железа, но и обнаруженное в ходе исследования снижение антиоксидантной защиты плазмы, что, как показали предыдущие работы, вызывает в ней активацию ПОЛ. .Повышение перекисного окисления в плазме повреждает структуры тромбоцитов и сосудов и влияет на их функции. 37,38 У новорожденных телят с дефицитом железа. Ускорение агрегации тромбоцитов указывает на повышение чувствительности их рецепторов к стимулирующим воздействиям извне. 39 Кроме того, активное развитие агрегации тромбоцитов в ответ на ристомицин у телят с дефицитом железа следует рассматривать как следствие повышения их чувствительности к фактору Виллибранда. 10 Кроме того, происходящее у этих животных ускорение агрегации тромбоцитов косвенно говорит об усилении в их тромбоцитах обменных процессов арахидоновой кислоты с образованием избыточного тромбоксана А 2 . 40
Ускорение агрегации тромбоцитов указывает на повышение чувствительности их рецепторов к стимулирующим воздействиям извне. 39 Кроме того, активное развитие агрегации тромбоцитов в ответ на ристомицин у телят с дефицитом железа следует рассматривать как следствие повышения их чувствительности к фактору Виллибранда. 10 Кроме того, происходящее у этих животных ускорение агрегации тромбоцитов косвенно говорит об усилении в их тромбоцитах обменных процессов арахидоновой кислоты с образованием избыточного тромбоксана А 2 . 40
Применение комбинации ферроглюкина, полисона и крезацина заставило новорожденных телят с железодефицитным состоянием почувствовать насыщение своего организма железом, положительную динамику эритроцитов и общие показатели состояния животных. Оказанное воздействие на организм обследованных телят сопровождалось снижением интенсивности процессов их ПОЛ в плазме, что ослабляло ее повреждающее влияние на эндотелий и тромбоциты печени.Обнаруженная нормализация агрегации тромбоцитов телят с железодефицитным состоянием после приема комбинации ферроглюкина, полизона и крезацина во многом является следствием положительного воздействия комбинации этих средств на механизмы функционирования ПОЛ внутренних тромбоцитов, рецепторных и пострецепторных тромбоцитов. 10 Развивающееся в этих условиях увеличение времени агрегации тромбоцитов в ответ на ристомицин указывало на снижение в крови этих телят кофактора адгезии — фактора Виллибранда. 20
Заключение
Для новорожденных телят с дефицитом железа характерно снижение антиоксидантной защиты плазмы крови, усиление в ней процессов ПОЛ, повышение гемостатической активности тромбоцитов. Применение комбинации ферроглюкина, полисона и крезацина у новорожденных телят с дефицитом железа позволяет реально усилить антиоксидантную защиту плазмы, ослабить в ней активность ПОЛ, нормализовать активность тромбоцитов.
Благодарности
Автор благодарит Курский институт социального воспитания (филиал) Российского государственного социального университета и Всероссийский научно-исследовательский институт физиологии, биохимии и питания животных «Институт села» за предоставленные лабораторное оборудование и реактивы.
Конфликт интересов
Конфликт интересов не декларируется.
Список литературы
- Явуз Э., Тодоров Н., Ганчев Г. и Неделков К. Влияние кормления различными молочными программами на рост, здоровье и развитие молочных телят. Болгарский сельскохозяйственный журнал. Наука. 2015; 21: 384-93.
- Кутафина Н.В., Медведев И.Н. Динамика физиологических показателей телят в раннем онтогенезе. Зоотехния. 2015; 3: 25-7.
- Амелина И. В., Медведев И. Н. Транскрипционная активность ядрышковых организующих областей хромосом у населения Курской области. Вестник экспериментальной биологии и медицины. 2009; 147 (6): 730-32.
CrossRef - Амелина И. В., Медведев И. Н. Оценка зависимости интенсивности мутагенеза от активности хромосомных участков ядрышка-организатора у коренного населения Курской области. Вестник экспериментальной биологии и медицины. 2008; 145 (1): 68-71.
CrossRef - Белова Т. А. и Медведев И. Н. Онтогенетическая динамика микрореологических свойств эритроцитов и тромбоцитов у телят разного физиологического статуса. Курск. 2011; 268.
- Глаголева Т.И. Функциональные и биохимические особенности организма и показателей крови крупного рогатого скота в онтогенезе. Ветеринария, зоотехния и биотехнология. 2015; 3: 53-66.
- Медведев И. Н., Лапшина Е. В., Ю. С. З. Активность тромбоцитарного гемостаза у детей с деформациями позвоночника. Вестник экспериментальной биологии и медицины. 2010; 149 (5): 645-46.
CrossRef - Шитикова А.С. Тромбоцитопатия врожденная и приобретенная. Санкт-Петербург. 2008; 320.
- Симоненко В. Б., Медведев И. Н., Мезенцева Н. И., Толмачев В. В. Антиагрегационная активность сосудистой стенки у больных артериальной гипертензией с метаболическим синдромом. Клиническая медицина. 2007; 85 (7): 28-30.
- Симоненко В.Б., Медведев И.Н., Толмачев В.В. Сравнительная оценка влияния сульфгидрильных и фосфатных ингибиторов АПФ на агрегацию тромбоцитов у пациентов, страдающих артериальной гипертензией с метаболическим синдромом. Клиническая медицина. 2007; 85 (4): 24.
- Кутафина Н.В., Медведев И.Н. Агрегация тромбоцитов у клинически здоровых лиц второго возраста, проживающих в Курской области. Успехи геронтологии. 2015; 5 (4): 267-70.
CrossRef - Медведев И.Н., Громнацкий Н. И. Коррекция тромбоцитарного гемостаза и снижение биологического возраста при метаболическом синдроме. Клиническая медицина. 2005; 83 (8): 54-7.
- Симоненко В. Б., Медведев Н., Кумова Т. А. Патогенетические аспекты артериальной гипертензии при метаболическом синдроме. Военно-медицинский журнал. 2010; 331 (9): 41-4.
- Симоненко В. Б., Медведев И. Н., Толмачев В. В. Патогенетические аспекты артериальной гипертензии при метаболическом синдроме. Клиническая медицина. 2011; 89 (1): 49-51.
- Громнацкий Н.И., Медведев И.Н. Немедикаментозная коррекция нарушения гемостаза тромбоцитов у больных АГ с метаболическим синдромом. Клиническая медицина. 2003; 81 (4): 31-4.
- Медведев И. Н., Громнацкий Н. И., Голиков Б. М., Аль-Зураики Э. М., Ли В. И. Влияние лизиноприла на агрегацию тромбоцитов у пациентов с артериальной гипертензией с метаболическим синдромом. Кардиология. 2004; 44 (10): 57-9.
- Симоненко В.Б., Медведев И. Н., Толмачев В. В. Динамика активности первичного гемостаза у пациентов с артериальной гипертензией и метаболическим синдромом, получавших кандесартан // Клиническая медицина. 2011; 89 (3): 35-8.
- Медведев И. Н., Громнацкий Н. И. Влияние амлодипина на внутрисосудистую активность тромбоцитов у пациентов с артериальной гипертензией и метаболическим синдромом. Клиническая медицина. 2005; 83 (2): 37-9.
- Медведев И. Н., Громнацкий Н. И. Влияние небиволола на агрегацию тромбоцитов у больных артериальной гипертензией с метаболическим синдромом. Клиническая медицина. 2005; 83 (3): 31-3.
- Медведев И. Н., Кумова Т. А. Снижение агрегации тромбоцитов у пациентов с артериальной гипертензией и метаболическим синдромом, принимающих лозартан. Российский кардиологический журнал. 2008; 1: 40-2.
- Медведев И. Н., Громнацкий Н. И., Волобуев И. В., Осипова В. М., Дементьев В. И., Стороженко М. В. Тромбоцитарный гемостаз у больных АГ с метаболическим синдромом и его коррекция ловастатином. Клиническая медицина. 2004; 82 (10): 37-41.
- Медведев И. Н., Громнацкий Н. И. Влияние гипокалорийной диеты на реологию тромбоцитов у пациентов с метаболическим синдромом. Клиническая медицина. 2006; 84 (3): 49-52.
- Медведев И. Н., Савченко А. П. Коррекция активности тромбоцитов регулярными физическими упражнениями у молодых людей с повышенным нормальным артериальным давлением. Российский кардиологический журнал. 2010; 2 (82): 35-40.
- Глаголева Т.И. Способность к агрегации эритроцитов, тромбоцитов и лейкоцитов у новорожденных телят. Ветеринарный врач. 2015; 3: 49-53.
- Глаголева Т. И., Ю. С. З., Медведев И. Н. Сосудистый контроль агрегации тромбоцитов у новорожденных телят с дефицитом железа, получавших ферроглюкин. Современные высокие технологии. 2011; 3: 93.
- Хусаинов В.Р., Фенченко Н.Г. Полизон Влияние на рост свиней и качество их продукции. Сибирский вестник сельскохозяйственных наук. 2005; 2: 89-93.
- Гурьянов А. М., Петуненков С.В., Борин А.В., Макаров И.I. Эффективность кормления телят и кормовых добавок Natuphos krezatcina в составе кормов. Зоотехния. 2007; 10: 10-11.
- Чевари С., Эндиал Т. и Стренджер Дж. Определение антиоксидантных параметров крови и их диагностическое значение у пожилых людей. Лабораторная работа. 1991; 10: 9-13.
- Медведев И. Н., Савченко А. П., Ю. С. З., Краснова Е. Г., Кумова Т. А., Гамолина О. В., Скорятина И. А., Фадеева Т. С. Методические подходы к изучению реологических свойств крови в различных состояниях. Российский кардиологический журнал. 2009; 5: 42-5.
- Медведев И. Н., Амелина И. В. Связь морфологических фенотипических характеристик человека с активностью хромосомных ядрышковых регионов-организаторов в межфазном ядре клетки у коренного населения Курской области. Морфология. 2012; 142 (4): 87-91.
- Медведев И. Н., Брюховецкий А. Г. Применение верошпирона и степень агрегации тромбоцитов при артериальной гипертензии с абдоминальным ожирением. Клиническая медицина. 2014: 92 (3): 50-53.
- Медведев И. Н., Скорятина И. А. Агрегация эритроцитов у пациентов с артериальной гипертензией и дислипидемией, получавших правастатин. Клиническая медицина. 2014; 92 (11): 34-38.
- Медведев И. Н., Скорятина И. А. Правастатин в коррекции антиагрегантного контроля сосудистой стенки над клетками крови у больных артериальной гипертензией и дислипидемией. Сердечно-сосудистая терапия и профилактика. 2014; 13 (6): 18-22.
- Медведев И. Н., Скорятина И. А. Динамика гемостаза тромбоцитов у пациентов с артериальной гипертензией и дислипидемией, принимающих симвастатин. Российский кардиологический журнал. 2010; 1 (81): 54-8.
- Медведев И. Н., Скориятина И. А. Динамика микрореологических свойств эритроцитов у больных артериальной гипертензией и дислипидемией, получавших аторвастатин. Клиническая медицина. 2012; 90 (6): 42-5.
- Симоненко В.Б., Медведев И. Н., Толмачев В. В. Влияние ирбесартана на функцию гемокоагуляционного компонента гемостаза у больных артериальной гипертензией при метаболическом синдроме. Клиническая медицина. 2010; 88 (6): 27-30.
- Медведев И. Н., Скориятина И. А. Влияние ловастатина на адгезивную и агрегационную функцию тромбоцитов у больных артериальной гипертензией и дислипидемией. Клиническая медицина. 2010; 88 (2): 38-40.
- Медведев И.Н., Скорятина И. А. Влияние флувастатина на агрегацию клеток крови у больных артериальной гипертензией и дислипидемией. Сердечно-сосудистая терапия и профилактика. 2013; 12 (2): 18-24.
CrossRef - Burnier L., Fontana P., Kwak B.R и Angelillo-Scherrer A. Микрочастицы клеточного происхождения в гемостазе и сосудистой медицине. Haemost. 2009; 101: 439-51.
CrossRef - Симоненко В. Б., Медведев И. Н., Гамолина О. В. Активность первичного гемостаза у пациентов с артериальной гипертензией и нарушением толерантности к глюкозе, получавших трандолаприл. Клиническая медицина. 2011; 89 (2): 29-31.
 Санкт-Петербург. 2008; 320.
Санкт-Петербург. 2008; 320. 2011; 89 (3): 35-8.
2011; 89 (3): 35-8. Зоотехния. 2007; 10: 10-11.
Зоотехния. 2007; 10: 10-11. Клиническая медицина. 2010; 88 (6): 27-30.
Клиническая медицина. 2010; 88 (6): 27-30.Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie.Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г. ,
браузер автоматически забудет файл cookie.Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
 ,
браузер автоматически забудет файл cookie.Чтобы исправить это, установите правильное время и дату на своем компьютере.
,
браузер автоматически забудет файл cookie.Чтобы исправить это, установите правильное время и дату на своем компьютере.Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Физиологическая активность подавляет синаптическую функцию за счет воздействия на прайминг пузырьков
Abstract
Нейроны задействуют компенсирующие гомеостатические синаптические изменения для поддержания своей общей скорости активации. Мы исследовали индукцию и проявление стойкой пресинаптической адаптации. Мы исследовали влияние умеренного повышения уровня внеклеточного калия на увеличение пиков пирамидных нейронов гиппокампа в физиологическом диапазоне. После нескольких дней умеренной деполяризации высвобождение глутамата адаптировалось, о чем свидетельствует увеличенное несоответствие между количеством активных, FM1-43-позитивных, глутаматергических синапсов и общим количеством синапсов, определяемым окрашиванием антител везикулярного переносчика глутамата-1. Удивительно, но адаптация глутаматных окончаний была полной; Размер пула рециркулирующих везикул в оставшихся активных синапсах не был существенно изменен адаптацией. Тетродотоксин (ТТХ), но не блокада постсинаптических рецепторов, адаптация, вызванная обратной деполяризацией, и ТТХ, добавленный к нормальной инкубационной среде, увеличивали количество активных синапсов, предполагая, что нормальная пиковая активность поддерживает устойчивый процент неактивных окончаний. Хроническая умеренная деполяризация подавляла EPSC и уменьшала размер легко высвобождаемого пула везикул (RRP).Несколько часов электростимуляции с частотой 10 Гц также снизили размер RRP, подтверждая, что один спайк вызывает адаптацию, а сильная стимуляция вызывает более быструю пресинаптическую адаптацию. Несмотря на важность изменения RRP для адаптации, ультраструктурные эксперименты не выявили изменений в количестве пристыкованных или общих синаптических пузырьков. Более того, альфа-латротоксин индуцировал высвобождение везикул в адаптированных синапсах, что согласуется с идеей, что адаптация является результатом изменений примирования везикул.Эти результаты показывают, что глутаматергическая нейротрансмиссия постоянно адаптируется к изменениям электрической активности в широком физиологическом диапазоне.
Удивительно, но адаптация глутаматных окончаний была полной; Размер пула рециркулирующих везикул в оставшихся активных синапсах не был существенно изменен адаптацией. Тетродотоксин (ТТХ), но не блокада постсинаптических рецепторов, адаптация, вызванная обратной деполяризацией, и ТТХ, добавленный к нормальной инкубационной среде, увеличивали количество активных синапсов, предполагая, что нормальная пиковая активность поддерживает устойчивый процент неактивных окончаний. Хроническая умеренная деполяризация подавляла EPSC и уменьшала размер легко высвобождаемого пула везикул (RRP).Несколько часов электростимуляции с частотой 10 Гц также снизили размер RRP, подтверждая, что один спайк вызывает адаптацию, а сильная стимуляция вызывает более быструю пресинаптическую адаптацию. Несмотря на важность изменения RRP для адаптации, ультраструктурные эксперименты не выявили изменений в количестве пристыкованных или общих синаптических пузырьков. Более того, альфа-латротоксин индуцировал высвобождение везикул в адаптированных синапсах, что согласуется с идеей, что адаптация является результатом изменений примирования везикул.Эти результаты показывают, что глутаматергическая нейротрансмиссия постоянно адаптируется к изменениям электрической активности в широком физиологическом диапазоне.
Введение
Податливость — это давно изучаемое интригующее свойство синапсов. Долговременную синаптическую пластичность, пластичность, значительно превышающую индуцирующий стимул, можно разделить на две широкие формы: геббовскую и гомеостатическую (Burrone and Murthy, 2003). Геббовская депрессия и усиление синаптической функции характеризуются синаптической специфичностью и ассоциативностью.Напротив, гомеостатическая синаптическая пластичность характеризуется глобальным синаптическим изменением в ответ на изменения общего уровня активности клетки. Индукция гомеостатических синаптических изменений не обязательно включает коррелированную или антикоррелированную активность пресинаптических и постсинаптических партнеров. Поскольку он имеет противоположную полярность индуцирующему стимулу, гомеостаз может быть важным адаптивным ответом, чтобы удерживать механизмы положительной обратной связи, такие как долгосрочное потенцирование, от насыщения синаптического аппарата (Turrigiano and Nelson, 2004).Синаптический гомеостаз также может помочь предотвратить чрезмерное возбуждение и эксайтотоксичность, когда баланс между возбуждением и торможением нарушен (Moulder et al., 2004). Конечно, это могут быть искусственные различия, и могут существовать другие, гибридные формы синаптической пластичности. Если это так, то соответствующие индуцирующие стимулы и механизмы экспрессии таких форм неясны.
Поскольку он имеет противоположную полярность индуцирующему стимулу, гомеостаз может быть важным адаптивным ответом, чтобы удерживать механизмы положительной обратной связи, такие как долгосрочное потенцирование, от насыщения синаптического аппарата (Turrigiano and Nelson, 2004).Синаптический гомеостаз также может помочь предотвратить чрезмерное возбуждение и эксайтотоксичность, когда баланс между возбуждением и торможением нарушен (Moulder et al., 2004). Конечно, это могут быть искусственные различия, и могут существовать другие, гибридные формы синаптической пластичности. Если это так, то соответствующие индуцирующие стимулы и механизмы экспрессии таких форм неясны.
Недавно мы сообщили, что глутаматные синапсы претерпевают стойкую пресинаптическую депрессию, когда им сопутствует сильная кратковременная патофизиологическая деполяризация (Moulder et al., 2003, 2004). Таким образом, ответ кажется гомеостатическим или адаптивным, но вызывает ли физиологическая активность эту форму пресинаптической пластичности, неизвестно. Пресинаптическое изменение к сильной деполяризации отражается в депрессивных EPSC, вызванных в ответ на гипертоническую сахарозу, Ca 2+ -независимый стимулятор секреции, который, как считается, запускает избирательный экзоцитоз легко высвобождаемого пула везикул (RRP) (Rosenmund and Stevens, 1996). Поэтому мы называем синаптическое изменение «стойкой депрессией RRP».В настоящей работе мы показываем, что физиологическая активность активирует эту синаптическую адаптацию в двух направлениях, и исследуем механизмы экспрессии этой пресинаптической депрессии. Мы заключаем, что стойкая депрессия RRP возникает из-за пристыкованных пузырьков, которые неспособны подвергнуться «примированию», биохимическому переходу к готовности к высвобождению пузырьков (Sudhof, 2004). Некоторые синаптические мутанты, как известно, вызывают дефицит прайминга (Augustin et al., 1999), но долговременная синаптическая модуляция с помощью этого механизма ранее не описывалась.Интересно, что эта форма синаптической пластичности в первую очередь влияет на пресинаптические терминалы бинарным образом, при этом многие терминалы функционируют нормально, а подмножество полностью отрезано от способности экзоцитозировать везикулы до физиологических (и даже нефизиологических) стимулов. Таким образом, пластичность имеет общие черты как гомеостатической пластичности (адаптивной, неассоциативной), так и пластичности Хебба (специфичной для синапсов, а не глобальной).
Результаты
Повышение и снижение физиологической активности вызывают пресинаптическую инактивацию
Ранее мы продемонстрировали, что сильная тоническая деполяризация, например, достигаемая во время патофизиологических событий, приводит к стойкой пресинаптической адаптации в глутаматных синапсах (Moulder et al., 2003, 2004). Эта форма адаптации необычна по сравнению с другими формами долгосрочных пресинаптических и постсинаптических изменений, поскольку она включает бинарную потерю функции на глутаматных окончаниях как для Ca 2+ -зависимых, так и для Ca 2+ -независимых стимуляторов секреции (Moulder et al. др., 2004). Неизвестно, возникает ли эта форма адаптации в ответ на физиологическую активность. Чтобы определить это, мы переместили активность нейронов в обоих направлениях от исходного уровня в диссоциированных культурах постнатальных нейронов гиппокампа.
Во-первых, мы предположили, что если стойкая адаптация RRP происходит при нормальной активности, мы должны наблюдать увеличение процента функциональных синапсов с блокадой активности. Мы провоцировали клетки комбинированными блокаторами ТТХ и ионотропных глутаматных рецепторов в течение 6–10 дней для подавления электрической активности (500 нм TTX плюс 25 мкм d-APV плюс 1 мкм NBQX). Мы проанализировали стойкую адаптацию RRP путем тестирования несоответствия сигнатуры между поглощением FM1-43 и иммунореактивностью vGluT-1 (Moulder et al., 2004), анализ, позволяющий избежать неоднозначности, возникающей при анализе пресинаптической функции с помощью электрофизиологических анализов на основе постсинаптических рецепторов. Мы стимулировали поглощение FM1-43FX (фиксируемая форма FM1-43) с помощью сильной деполяризации (45 мм K + в течение 2 мин) с последующей фиксацией и мечения антителом vGluT-1 для идентификации всех глутаматергических пресинаптических элементов. На конфокальных изображениях, на которых мы исследовали интенсивность мечения FM1-43FX vGluT-1-положительной точки, мы обнаружили, что в необработанных культурах 20-30% vGluT-1-положительных окончаний были лишены поглощения FM1-43FX (рис.1 А , С ). Аналогичные проценты наблюдались и другими (Rosenmund et al., 2002; Altrock et al., 2003). В культурах с блокированной активностью мы обнаружили увеличение процента функциональных пресинаптических элементов (рис. 1 B , C ). Это противоположная полярность изменения, которое мы ранее наблюдали при кратковременной сильной деполяризации. Результат убедительно свидетельствует о том, что доля неактивных пресинаптических элементов, наблюдаемая в нашем (Moulder et al., 2004) и других (Rosenmund et al., 2002; Altrock et al., 2003) предыдущие исследования представляют класс пресинаптических элементов, которые функционально молчаливы из-за активности базальных нейронов.
Фигура 1.Неактивные синапсы уменьшаются из-за хронического бездействия. A , Контрольные синапсы, помеченные FM1-43 для идентификации активных синапсов и post hoc иммунофлуоресценцией vGluT-1 для идентификации глутаматергических синапсов. Стрелки указывают на неактивные глутаматные синапсы (vGluT-1 положительный, FM1-43 отрицательный). B , Аналогичные панели из культуры, обработанной блокаторами ТТХ и глутаматных рецепторов. Масштабная линейка 5 мкм. C , Резюме процента активных синапсов из полей, полученных в слепом анализе. FM, FM1-43. ∗ p <0,001. D , Резюме количественного анализа синаптической области, как определено с помощью иммунореактивности vGluT-1 или с помощью мечения FM1-43. После хронического бездействия наблюдалось значительное увеличение размера точки FM1-43.∗ p <0,001. Для обоих типов: C и D , n = 6. Столбики ошибок представляют SEM.
Помимо изменения процентного содержания функциональных синапсов, культуры с блокированной активностью демонстрировали небольшое, но значительное увеличение площади точек FM1-43 (рис. 1 D ). Это согласуется с предыдущей работой, предполагающей изменение размера синапсов при блоке активности (Murthy et al., 2001). Средняя интенсивность FM1-43 puncta также увеличилась с 1691 ± 43 (контроль) до 1769 ± 50 (TTX) ( p <0.002).
Чтобы проверить, может ли пресинаптическая функция еще больше подавляться по сравнению с исходным уровнем за счет повышения активности, мы титровали внеклеточную концентрацию K + ([K + ] o ), чтобы получить увеличение частоты спонтанных спайков. Мы обнаружили, что увеличение на 6 мм [K + ] o (от базальных 4 мм) увеличивало частоту спайков с 0,51 ± 0,26 до 1,69 ± 0,53 Гц (рис. 2), все еще в пределах нормы. спонтанная пиковая активность для пирамидных клеток in situ (Gao et al., 1998, 1999). Хронически, к концу 6–9-дневного лечения мы обнаружили, что мембранный потенциал покоя находился всего на 10,7 ± 0,6 мВ от порога спайка по сравнению с 16,7 ± 1,3 мВ в контрольных нейронах ( n = 8 для каждого; p <0,001), что подтверждает повышенную возбудимость обработанных клеток в течение периода воздействия.
Фигура 2.Увеличение на 6 мм [K + ] o резко увеличивает выбросы и хронически приближает нейроны к пороговому значению.Характерный след записи токовых клещей во время острого наложения дополнительных 6, 8 и 10 мм [K + ] o . Обратите внимание на ступенчатую деполяризацию мембранного потенциала покоя и увеличение пиков с 6 мм [K + ] o .
После короткой 16-часовой инкубации с 6 мм [K + ] o мы не индуцировали заметной синаптической адаптации (данные не показаны). Это временной интервал, в течение которого 30 мм [K + ] o вызывают сильную пресинаптическую адаптацию (Moulder et al., 2004). Однако мы обнаружили, что хроническое (6–9 дней) воздействие легкой деполяризации вызывает высоконадежное снижение процента vGluT-1-положительных точек с соответствующей флуоресценцией FM1-43 (рис. 3 A – C ). Кроме того, мы обнаружили, что блокада синаптических рецепторов глутамата (25 мкм d-APV плюс 1 мкм NBQX) не смогла изменить снижение функциональных пресинаптических элементов (рис. 3 C ). Этот результат согласуется с идеей, что постсинаптическая рецепторная активность не требуется для этой формы синаптической пластичности.С другой стороны, ТТХ 500 нм предотвращал вызванное деполяризацией уменьшение процентного содержания функциональных окончаний (рис. 3 C ), демонстрируя, что поведение пиков, а не изменение мембранного потенциала покоя, было важным для эффекта увеличения [K + ] o . Уменьшение функциональных пресинаптических сайтов было первичным изменением, обнаруженным в этих экспериментах по совместному мечению FM1-43 / vGluT-1, поскольку не было измерено статистически значимых различий в площади или интенсивности точек.Для точек vGluT-1 средняя площадь составила 93,4 ± 10,2 (контроль) и 76,4 ± 3,9 (K + ) ( p <0,380), а средняя интенсивность составила 1841 ± 574 (контроль) и 1636 ± 424 (K ). + ) ( p <0,508 и n = 6 экспериментов). Для точек FM1-43 средняя площадь составила 67,2 ± 14,6 (контроль) и 48,7 ± 9,8 (K + ) ( p <0,143), а средняя интенсивность составила 1588 ± 397 (контроль) и 1420 ± 390 (K ). + ) ( p <0,063 и n = 6 экспериментов).В целом, эти эксперименты показывают, что уменьшение и увеличение пиковой активности в физиологическом диапазоне изменяет процент функциональных пресинаптических элементов в глутаматных синапсах без явного изменения количества функционально доступных пузырьков, остающихся на активных окончаниях.
Рисунок 3.Неактивные синапсы увеличиваются при хроническом воздействии на слегка повышенные [K + ] o . A , Контрольные синапсы, помеченные FM1-43 для идентификации активных синапсов и post hoc иммунофлуоресценцией vGluT-1 для идентификации глутаматергических синапсов.Стрелки указывают на неактивные глутаматные синапсы (vGluT-1 положительный, FM1-43 отрицательный). B , Аналогичные панели из культуры, обработанной дополнительными 6 мм [K + ] o . Масштабная линейка 5 мкм. C , Резюме процента активных синапсов из полей, полученных в слепом анализе ( n = 3–12). ∗ p <0,001 и ∗∗ p <0,002 по сравнению с контролем. FM, FM1-43. D , Кумулятивные гистограммы интегральных интенсивностей FM1-43 точек для> 200 терминалов (определяемых иммунореактивностью vGluT-1) в каждой группе из одного эксперимента.Сплошная линия представляет точки из контрольных культур, а пунктирная линия представляет точки из культур, обработанных K + . E , F , Гистограммы значений интегральной интенсивности (площадь × интенсивность) для отдельных точек FM1-43 из контроля ( E ) или K + -обработанных ( F ) культур. E и F содержат группированные данные с тех же терминалов, представленные кумулятивными графиками в D .Размер корзины составлял 10 000 условных единиц (AU). Обратите внимание на две популяции синапсов. Гистограммы не вычитаются из фона, учитывая ненулевой начальный пик, представляющий неактивные терминалы. Обратите внимание на изменение оси y между E и F . Планки погрешностей представляют SEM.
Для дальнейшего рассмотрения очевидной бимодальности пресинаптического изменения функции, мы исследовали большое количество терминалов в серии из трех экспериментов, чтобы мы могли построить гистограммы интенсивности окрашивания.Данные одного из этих экспериментов проиллюстрированы на рисунке 3. Контрольные терминалы демонстрировали распределение интенсивности окрашивания, которое было явно бимодальным, что демонстрируется как на гистограммах совокупной вероятности (фиг. 3 D , сплошная линия), так и на обычных гистограммах (фиг. 3 ). E ). Первый пик распределения не превышал фоновые уровни флуоресценции, полученные из клеточных областей, прилегающих к vGluT-1-положительной точке (данные не показаны). Поэтому мы интерпретируем этот пик как неактивные терминалы.Второй пик интегрированного распределения интенсивности был шире, предположительно отражая диапазон количества пузырьков, составляющих рециклирующий пул в различных синапсах гиппокампа (Harata et al., 2001). Когда мы исследовали терминалы в деполяризованных культурах (фиг.3 D , пунктирная линия, F ), мы обнаружили, что основным отличием от контроля было изменение относительной амплитуды двух пиков (фиг.3 E ). , F ), без изменения средних значений двух пиков: 6389 ± 368 единиц флуоресценции (контроль, первый пик), 6291 ± 230 единиц флуоресценции (K + , первый пик; p <0.814), 49 911 ± 2571 единиц флуоресценции (контроль, второй пик) и 55 362 ± 1259 единиц флуоресценции (K + , второй пик; p <0,166). Аналогичные результаты были получены с гистограммами из двух других экспериментов, в которых было исследовано> 200 точек FM1-43 для каждого условия (данные не показаны). Эти результаты согласуются с гипотезой о том, что стойкое синаптическое изменение представляет собой прежде всего бинарное изменение функции в подмножестве глутаматных синапсов. Результаты совпадают с нашими выводами из независимых экспериментов с меньшим количеством терминалов и предыдущих экспериментов с очень сильной стимуляцией (Moulder et al., 2004).
Мы проверили, изменилась ли постсинаптическая функция при длительном лечении. Мы обнаружили, что увеличение [K + ] o на 6 мм не повлияло на процент точек GluR2, относящихся к vGluT-1 (рис. 4 A – C ), что свидетельствует об отсутствии увеличения количества точек GluR2. постсинаптически безмолвные синапсы. Размер (фиг. 4 D ) и средняя интенсивность (данные не показаны) точки GluR2 также не изменились, что согласуется с идеей, что плотность постсинаптических рецепторов не изменилась.В отдельных экспериментах мы также не наблюдали изменения размера или средней интенсивности точки GluR1 (данные не показаны).
Рисунок 4.Хроническое воздействие слегка повышенных [K + ] o не изменяет постсинаптические рецепторы глутамата. A , Контрольные синапсы, коиммуноокрашенные для vGluT-1 и GluR2. B , Аналогичные панели из культуры, обработанной дополнительными 6 мм [K + ] o .Масштабная линейка 5 мкм. C , Сводная информация о процентном содержании vGluT-1-положительных сайтов, меченных GluR2. D , Сводная информация о площади GluR2-положительных точек. Для C и D статистически значимых различий не наблюдалось. n = 3 для каждого. E , Типичные следы после экзогенного применения 50 мкМ каиновой кислоты из контрольного нейрона и нейрона, обработанного K + .Емкость контрольной ячейки составляла 22,7 пФ, а ячейки, обработанной K + , — 24,3 пФ. Планки погрешностей представляют SEM.
Электрофизиологические тесты адаптации к легкой деполяризации
В дополнение к экспериментам по визуализации, которые предполагали пресинаптические изменения без доказательств сопутствующих постсинаптических изменений, мы выполнили электрофизиологические тесты локуса изменения. После 6–9 дней пребывания в условиях умеренной деполяризации мы исследовали электрофизиологические реакции на экзогенное применение антагониста рецепторов AMPA / KA каиновой кислоты.Мы не обнаружили изменений плотности тока (рис.4 E ), со средним значением -53,2 ± 7,0 пА / пФ в контрольных нейронах и -45,4 ± 3,7 пА / пФ в нейронах, обработанных K + ( n = 15; p <0,34). Следовательно, основные изменения функции в ответ на изменения активности в наших условиях являются пресинаптическими.
Учитывая, что незрелые пресинаптические элементы могут существовать без постсинаптического партнера (Ahmari et al., 2000) и что эти незрелые профили могут вносить вклад в несовпадающее окрашивание FM1-43 / vGluT-1, мы проверили, являются ли функционально инактивированные терминалы (рис.3) влияют на сигнализацию, выполняя электрофизиологию. Мы обнаружили, что аутаптические ЭПСК, вызванные потенциалом действия, подавлялись через 6–9 дней лечения с увеличением на 6 мм [K + ] o с -6143 ± 1266 пА (контроль) до -1780 ± 512 пА (). n = 9 для каждого; p <0,006) (Рис.5 A , D ). И эксперименты FM1-43, и эти физиологические эксперименты проводили с Ca 2+ -зависимым высвобождением медиатора. Чтобы проверить, вызывают ли Ca 2+ -независимые стимуляторы секреции также подавленное высвобождение, мы использовали гипертоническую сахарозу (0.5 м), что вызывает экзоцитоз RRP (Rosenmund and Stevens, 1996). Ответы сахарозы были снижены с -980,9 ± 202,4 пКл (контроль) до -436,7 ± 78,1 пКл ( n = 9 для каждого; p <0,018), аналогично ответам потенциала действия (рис. 5 B , D ), что указывает на то, что стадии адаптации пресинаптической функции к активности не связаны с притоком Ca 2+ . Чтобы проверить, является ли уменьшение размера RRP результатом изменения скорости восстановления пула везикул, мы индуцировали высвобождение глутамата с помощью парных нанесений сахарозы 3.С интервалом 5 с для измерения относительного восстановления доступной сахарозе RRP (Altrock et al., 2003). Размер RRP восстановился до 43,8 ± 2,5 и 40,0 ± 3,0% в контроле и нейронах, обработанных K + , соответственно ( p <0,331 и n = 11 для каждого), что указывает на то, что хроническая легкая деполяризация не влияла на кинетика заправки РРП.
Рисунок 5.Хроническая умеренная деполяризация подавляет глутаматергическую синаптическую передачу. A , Типичные электрически вызванные аутаптические EPSC из одиночных нейронов, поддерживаемых в контрольной среде, или после хронического лечения с дополнительными 6 мм [K + ] o . B , Типичные вызванные сахарозой токи от тех же нейронов, что и в A . C , Образцы кривых, показывающие mEPSC, вызванные нанесением 0,3 мкл сахарозы и записанные в контрольных нейронах или нейронах, обработанных K + . D , Краткое изложение влияния хронической легкой деполяризации на амплитуду EPSC, размер RRP, частоту и амплитуду mEPSC. Все значения нормализованы к контрольным значениям из того же эксперимента.Исходные значения для каждого представлены в разделе «Результаты». ∗ p <0,02. Планки погрешностей представляют SEM.
Мы также оценивали мЭПСК после хронической легкой деполяризации. mEPSCs были вызваны с помощью 0,3 м сахарозы (Rosenmund and Stevens, 1996; Moulder et al., 2004). В 15 нейронах, исследованных для каждой экспериментальной группы, мы обнаружили, что частота (23,3 ± 1,5 Гц для контроля; 11,3 ± 1,2 Гц для K + ; p <0,001), но не амплитуда (20,7 ± 1,2 пА для контроля ; 21.0 ± 1,1 Гц для K + ; p <0,88), мЭПСК уменьшалась на 6–9 дней лечения с увеличением на 6 мм [K + ] o (Рис. 5 C , D ). Это согласуется с классической идеей, что изменения частоты mEPSC представляют собой пресинаптический локус, тогда как изменения индивидуального размера mEPSC представляют собой постсинаптический локус. Время нарастания mEPSC 10–90% (1,08 ± 0,07 мс для контроля и 1,24 ± 0,07 мс для K + ; p <0,103) и время спада 10–90% (2.21 ± 0,08 мс для контроля и 2,15 ± 0,08 мс для K + ; p <0,622) также не изменились после хронической легкой деполяризации.
Поскольку хроническая умеренная деполяризация, по-видимому, изменяет пресинаптическую передачу сигналов глутамата, мы хотели проверить, что только усиление пикового воздействия может вызвать такую же депрессию. Чтобы ответить на этот вопрос, мы электрически стимулировали нейроны с частотой 10 Гц в течение 16 часов, используя полевую стимуляцию. После этой обработки регистрировали вызванные сахарозой токи (рис.6 А ). Как и в случае хронической легкой деполяризации K + (рис. 5), хронический всплеск, хотя и в более коротком временном масштабе, уменьшал размер RRP (рис. 6 A , B ). Вызванные сахарозой mEPSCs также обнаруживают пониженную частоту (Fig. 6 B ), что снова согласуется с идеей, что пресинаптические изменения лежат в основе этого дефицита передачи сигналов. После стимуляции 10 Гц амплитуда mEPSC (19,4 ± 0,4 пА для контроля и 19,0 ± 0,7 пА для стимулированной), время нарастания 10–90% (1,38 ± 0,01).14 мс для контроля и 1,35 ± 0,13 мс для стимулированного; p <0,900) и время спада 10–90% (2,37 ± 0,11 мс для контроля и 2,57 ± 0,10 мс для стимулированного; p <0,468) не изменились. Для сравнения, 16 часов мягкой деполяризации (добавлено 6 мм K + ) не привели к изменению амплитуды вызванной ЭПСК (88 ± 15% от контроля; n = 10; p <0,75).
Рисунок 6.Продолжительная электрическая стимуляция подавляет глутаматергическую передачу сигналов. A , Типичные вызванные сахарозой токи от нестимулированного нейрона (слева) и от нейрона, подвергнутого хронической стимуляции полем 10 Гц (справа). B , Краткое изложение влияния хронической стимуляции на общий заряд токов, вызванных сахарозой ( n = 13; * p <0,02) и частоту (частоту) mEPSCs, вызванных 0,3 м сахарозой приложение ( n = 9; ∗∗ p <0,001). Планки погрешностей представляют SEM.
Тесты на изменение стыковки и прайминга пузырьков
Как визуализация, так и электрофизиологические результаты согласуются с пресинаптическим изменением, сильно участвующим в синаптической адаптации к длительным умеренным изменениям физиологического всплеска. Результаты предполагают устранение пузырьков в RRP путем функциональной инактивации пресинаптических окончаний. RRP ассоциирован с дополнением купированных пузырьков в пресинаптических активных зонах (Schikorski and Stevens, 2001).Следовательно, одна из гипотез, объясняющая подавление везикул, готовых к высвобождению, состоит в том, что клетки, подвергшиеся воздействию повышенных уровней активности, имеют меньше или не имеют пристыкованных везикул. Мы исследовали этот ультраструктурный вопрос, подвергая нейроны дополнительному 6 мм [K + ] o в течение 6–9 дней, затем вызывая аутаптические постсинаптические ответы (Рис. 7 C ), с последующей электронной микроскопией тех же клеток. (Рис.7 A , B ). Мы исследовали 88 случайных пресинаптических окончаний из контрольных культур и 98 терминалов из деполяризованных культур, но не обнаружили различий в количестве стыкованных (5.57 ± 0,59 для контроля; 6,04 ± 0,43 для K + ; p <0,555) или полные везикулы (55,73 ± 0,63 для контроля; 67,59 ± 7,87 для K + ; p <0,208) в терминалах из этих двух групп. Процент от общего числа стыкованных везикул также остался неизменным (данные не показаны).
Рисунок 7.Хроническая умеренная деполяризация не изменяет ультраструктуру синапсов, а скорее изменяет прайминг синаптических пузырьков. A , B , Электронные микрофотографии репрезентативных синапсов от контрольного нейрона ( A ) или от нейрона, обработанного K + ( B ). C , EPSC, записанные от контрольного нейрона, синапс которого показан в A (слева), и от обработанного K + нейрона, синапс которого показан в B (справа). D – E , Гистограммы количества пристыкованных пузырьков в синапсах из контроля ( D ) и нейронов, обработанных K + ( E ). Для D и E 88 синапсов (контроль) и 98 синапсов (K + ) были исследованы в трех экспериментах. F , Типичная текущая кривая mEPSCs, индуцированная приложением α-LTX. Переходные токи опосредуются рецепторами AMPA, потому что они блокируются 1 мкм NBQX. G , Краткое изложение эффекта хронической легкой стимуляции на частоту мЭПСК, индуцированных применением α-LTX. Статистически значимой разницы не наблюдалось (темные кружки). Однако хроническая умеренная стимуляция подавляла вызванные сахарозой токи (светлые кружки), зарегистрированные от тех же нейронов до применения α-LTX ( n = 12).Планки погрешностей представляют SEM.
Мы рассмотрели несколько возможных источников ошибки типа II (неспособность обнаружить разницу между контрольными и обработанными синапсами). Во-первых, световая микроскопия (сравнение FM1-43 / vGluT-1) предполагает, что всего лишь 20% терминалей могут испытывать полную потерю закрепленных везикул (Fig. 3 C ). Анализ мощности, выполненный для определения того, должна ли наша выборка выявлять потерю пузырьков в 20% синапсов, продемонстрировал мощность> 99.9%, что указывает на маловероятность ошибки типа II. Мы также рассмотрели возможность того, что, хотя функциональные эффекты проявляются по принципу «все или ничего» (Fig. 3 D – F ), морфологические изменения количества пристыкованных пузырьков могут быть более тонкими. В качестве еще одного теста на постепенное изменение количества везикул, пристыкованных к одному синапсу, мы выполнили анализ χ 2 , чтобы проверить, было ли в обработанных синапсах больше синапсов с менее чем тремя (менее половины среднего от контроля) пристыкованными пузырьками.Мы не обнаружили разницы между контрольной и экспериментальной группами в количестве синапсов, содержащих менее трех везикул ( p <0,125). Гистограммы количества стыкованных пузырьков на синапс показывают, что хроническая умеренная деполяризация не увеличивает количество синапсов с небольшим количеством стыкованных пузырьков (Рис. 7 D , E ).
Наконец, данные, показанные на рисунке 7, используют определение закрепленных везикул как тех везикул, которые соответствуют постсинаптической плотности и в пределах одного диаметра везикулы от плазматической мембраны (Pozzo-Miller et al., 1999; Мерфи и др., 2000). Если мы использовали более строгое определение стыковки, считая только пузырьки, контактирующие с плазматической мембраной (Harris and Sultan, 1995; Collin et al., 2001), количество стыкованных пузырьков уменьшилось (4,84 ± 0,52 для контроля; 4,76 ± 0,30 для K + ), но все еще не было статистически значимой разницы между контролем и нейронами, обработанными K + ( p <0,895). Следовательно, изменение морфологически купированных пузырьков не объясняет стойкую адаптацию RRP.
Генетическая делеция некоторых компонентов синаптического аппарата, таких как munc13–1 в глутаматных синапсах, связана с уменьшением общего пула доступных в клетках пузырьков и функциональных синаптических окончаний, но без ультраструктурных изменений (Augustin et al ., 1999; Розенмунд и др., 2002). Отличительной чертой синаптического фенотипа является то, что, хотя ни потенциалы действия, ни гипертоническая сахароза не могут вызвать высвобождение медиатора, α-LTX, активный компонент яда паука черной вдовы, способен вызывать высвобождение спящих пузырьков (Augustin et al., 1999). Интерпретация обычно состоит в том, что везикулы у этих мутантов останавливаются в морфологически закрепленном, но биохимически незрелом состоянии и, таким образом, недостаточно примированы для высвобождения (Augustin et al., 1999). Поскольку адаптированные к активности везикулы, по-видимому, все еще морфологически состыкованы (Fig. 7), мы можем предсказать, что эти везикулы могут быть задействованы для слияния с помощью α-LTX. Такой результат может указывать на изменение прайминга пузырьков как механизма адаптации.
Чтобы проверить, влияет ли физиологическая активность на процент функциональных пресинаптических окончаний за счет подавления прайминга везикул, мы провоцировали аутаптические клетки сахарозой, чтобы подтвердить стойкую адаптацию RRP, затем с помощью α-LTX, чтобы исследовать пристыкованные, но непраймированные пузырьки (рис.7 F ). Хотя сахарозные ответы показали характерную депрессию RRP, частота mEPSC, вызванных α-LTX, не отличалась статистически в деполяризованных и контрольных нейронах (фиг. 7 G ) ( p <0,451). Эти результаты дополнительно исключают недостаточность постсинаптической функции как фактора, способствующего гомеостатической депрессии, и в значительной степени указывают на недостаточность прайминга как механизма выражения пресинаптических изменений.
Обсуждение
Гомеостатическая пластичность — важное средство, с помощью которого нейроны могут регулировать свой общий уровень активности в условиях изменения входной силы.Ранее мы показали, что патофизиологические повышения в [K + ] o деполяризуют нейроны гиппокампа и впоследствии подавляют глутаматергическую передачу сигналов, предположительно действуя как форма гомеостатической регуляции (Moulder et al., 2003, 2004). Локусом этой депрессии был пресинаптический везикулярный RRP (Moulder et al., 2004). В настоящем исследовании мы обнаружили, что физиологическая активность изменяла нейротрансмиссию глутамата в течение нескольких дней. Опять же, первичной мишенью зависимой от активности модуляции была пресинаптическая, это указывает на то, что размер RRP (и / или доступность) является ключевым фактором в пластических изменениях синаптической функции.Более того, ответ на устойчивую активность приводит к очевидной регуляции пресинаптической функции по принципу «все или ничего», необычному механизму, который нарушает каноническую идею о том, что синаптический гомеостаз регулирует синаптическое усиление во всех синаптических контактах (Turrigiano et al., 1998).
Экспрессия синаптической адаптации
Несколько доказательств соответствовали нашему выводу о том, что хроническая умеренная деполяризация (например, более короткие и сильные деполяризации) (Moulder et al., 2004) подавляли глутаматергическую передачу сигналов, главным образом влияя на пресинаптическую функцию. Мы не обнаружили изменений в ответах на экзогенные агонисты AMPA / KA, не обнаружили изменений в амплитуде mEPSC и не обнаружили изменений в интенсивности маркировки или размере субъединиц постсинаптического рецептора. Вместо этого либо подавление, либо усиление пиков от исходного уровня модулировало процент FM1-43-позитивных синапсов, обеспечивая убедительные доказательства пресинаптической адаптации. В частности, несоответствие между функционально активными пресинаптическими окончаниями и наличием маркеров везикул указывает на уменьшение количества активных пресинаптических сайтов и изменение размера общего клеточного RRP.
Тот факт, что глутаматные синапсы были ультраструктурно нормальными с точки зрения общего числа и числа пристыкованных везикул (Рис. 7 A – E ), по-видимому, противоречит результатам FM1-43 и электрофизиологии. Однако наблюдение, что α-LTX, но не гипертоническая сахароза, индуцировало высвобождение синаптических пузырьков в этих местах (фиг. 7 G ), согласовывает результаты двух экспериментов. Нейроны гиппокампа мышей, генетически удаленных для Munc13-1, белка, необходимого для перехода между стыковкой синаптических везикул и праймированием, демонстрируют такой же избирательный ответ на α-LTX (Augustin et al., 1999). Таким образом, мы заключаем, что прайминг, вероятно, является целью синаптических адаптаций к хроническим изменениям активности. Интересно, что нейроны мышей, генетически удаленных для др. Пресинаптических белков, включая фагот компонента цитоматрикса, также обнаруживают сходный фенотип (Altrock et al., 2003). Поскольку праймирование является критическим этапом в цикле синаптических пузырьков, идея о том, что электрическая активность может модулировать прайминг, уже была высказана (Dulubova et al., 2005), а роль прайминга была показана для краткосрочной синаптической пластичности (Junge et al. al., 2004). Однако, насколько нам известно, текущая работа представляет собой первое сообщение о вовлечении прайминга в более долгосрочные адаптивные изменения синаптической функции.
После легкой деполяризации процент инактивированных окончаний, обнаруженных сравнениями FM1-43 / vGluT-1, казался менее заметным, чем изменения в электрофизиологически определенной RRP (сравните фиг. 3 C , 5 D ). В основе этого количественного расхождения могут лежать несколько факторов. Возможно, что постепенные изменения размера RRP на отдельных терминалах могут способствовать электрофизиологическим результатам, но не могут быть обнаружены анализом интенсивности FM1-43.Это кажется маловероятным, потому что не было последовательного изменения интегральной интенсивности активных синапсов после легкой деполяризации (рис. 3 E , F ), и мы смогли обнаружить постепенные изменения интенсивности, вызванные TTX. Также возможно, что синаптическая адаптация включает подавление vGluT-1 (De Gois et al., 2005), что может подавлять EPSC, но не окрашивание FM1-43. Это маловероятно, потому что амплитуда mEPSC не изменилась, и мы не обнаружили изменений в интенсивности окрашивания vGluT-1 в деполяризованных культурах.В электрофизиологических исследованиях может быть необнаруженный постсинаптический вклад, хотя некоторые результаты (см. Ниже) делают это маловероятным. Наконец, могут наблюдаться эффекты длительной деполяризации на общее количество образованных синаптических контактов, эффект, который может быть обнаружен несколькими электрофизиологическими анализами, но не нашим методом визуализации. Однако эта гипотеза не может объяснить сходство частот mEPSC, вызванных α-LTX, в контрольной и деполяризованной культурах (рис. 7 G ).Таким образом, мы предполагаем, что небольшие вклады нескольких из этих механизмов должны добавить, чтобы объяснить количественное несоответствие.
Требования к индукции
Наши первоначальные эксперименты были выполнены с использованием продолжительного воздействия дополнительных 6 мм [K + ] o . Эта обработка приблизила мембранный потенциал нейронов к пороговому значению и увеличила выбросы. Учитывая, что даже незначительные изменения в пресинаптическом мембранном потенциале покоя могут оказывать сильное воздействие на [Ca 2+ ] и (Awatramani et al., 2005), мы стремились определить роль пиков в депрессии нейротрансмиссии глутамата. Один только TTX увеличивал процент FM1-43-положительных активных терминалов и предотвращал снижение процента активных терминалов, вызванное длительной умеренной деполяризацией. Кроме того, стимуляция поля для непосредственного индуцирования пиков также подавляла физиологически определенный RRP. Вместе эти результаты демонстрируют, что физиологической пиковой активности достаточно, чтобы вызвать пресинаптическую адаптацию.
Интересно, что стимуляция поля с частотой 10 Гц вызывала синаптическую адаптацию быстрее (в течение 16 часов), чем легкая деполяризация. Эти результаты согласуются с общей картиной, в которой сильная патофизиологическая деполяризация (60 мм [K + ] o ) вызывает пресинаптическую адаптацию менее чем за 1 час (Moulder et al., 2004), но более низкие уровни деполяризации (30 мм K + ) требуется больше времени, чтобы вызвать детектируемую адаптацию (Moulder et al., 2004). Следовательно, стойкая депрессия RRP может быть вызвана широким спектром стимулов со скоростью, зависящей от силы стимула.
Сравнение с предыдущими результатами
Наши текущие результаты сосредоточены на пресинаптическом гомеостатическом изменении как после блокады хронической активности (рис. 1), так и после хронической активации активности. Как подробно описано выше, несколько линий экспериментов соответствовали идее, что это пресинаптическое изменение является основным локусом гомеостатической регуляции в нашей системе. Однако в этой области существуют разногласия относительно того, является ли в основном пресинаптическим (Bacci et al., 2001; Ши и др., 2001; Moulder et al., 2004; Де Гойс и др., 2005; Wang et al., 2005) или постсинаптическими (O’Brien et al., 1998; Turrigiano et al., 1998; Leslie et al., 2001; Ehlers, 2003; Ju et al., 2004; Perez-Otano and Ehlers, 2005; Richards et al., 2005; Wierenga et al., 2005) механизмы или даже оба (Luthi et al., 2001; Murthy et al., 2001; Alder et al., 2005; Thiagarajan et al., 2005) , регулируют гомеостаз. Непонятно, почему разные исследования по этому вопросу привели к разным результатам. Возможные причины включают методологию (например,g., витальные флуоресцентные красители, электрофизиология), нейрональная система, стадия развития (Burrone et al., 2002; Kirov et al., 2004) и продолжительность лечения. Последнее может представлять особый интерес, потому что в случае синаптической пластичности, вызванной нейротрофическим фактором мозга, ранние изменения, как сообщается, влияют на пресинаптический аппарат, тогда как поздние изменения влияют на постсинаптические компоненты (Alder et al., 2005).
Другой возможный фактор, способствующий этому, заключается в том, исследуется ли гомеостаз в ответ на повышенную активность или на блокаду активности.Кроме того, способы модуляции активности также могут различаться (например, лечение ТТХ по сравнению с блокадой глутаматных рецепторов). Наши текущие результаты не исключают возможного постсинаптического вклада в гомеостаз после блокады активности. Вместо этого они утверждают, что пресинаптические изменения, особенно функциональные RRP, действительно участвуют в последующем усилении синаптической функции (Fig. 1). Это может показаться контрастирующим с недавней работой, показывающей отсутствие изменений в поглощении FM1-43 после блокады активности (Wierenga et al., 2005). Однако разница, которую мы демонстрируем на рисунке 1, заключается в процентном соотношении активных (FM1-43-положительных) синапсов, которые, возможно, не были обнаружены в предыдущей работе.
Хотя о специфической активации синаптического GluR1 сообщалось при хронической блокаде активности (Ju et al., 2004; Thiagarajan et al., 2005), мы не наблюдали каких-либо различий в локализации GluR2 или GluR1 при хронической умеренной деполяризации. В соответствии с этим, мы не обнаружили никаких различий в кинетике mEPSC ни при обработке K + , ни при полевой стимуляции (рис.5, 6), что может свидетельствовать об изменении состава субъединиц синаптических рецепторов. Следовательно, оказывается, что модуляция трафика GluR1 может быть уникальной для гомеостаза после блокады активности.
В итоге мы обнаружили, что даже небольшое повышение [K + ] o , которое приводит к усилению пиков, может вызвать пресинаптические изменения при длительном воздействии. О подобном пресинаптическом эффекте недавно сообщалось in vivo при хронической стимуляции нервно-мышечного соединения (Belair et al., 2005), указывая на то, что эти механизмы, вероятно, важны для многих форм синаптической пластичности. Будет очень интересно определить, при каких условиях праймирование синаптических пузырьков принимает на себя основную роль в гомеостатической модуляции, и определить молекулярные пути, участвующие в индукции и экспрессии этой формы синаптической пластичности.
% PDF-1.2 % 304 0 объект > эндобдж xref 304 120 0000000016 00000 н. 0000002752 00000 н. 0000002852 00000 н. 0000003280 00000 н. 0000003559 00000 н. 0000003965 00000 н. 0000004088 00000 н. 0000004338 00000 п. 0000004472 00000 н. 0000004766 00000 н. 0000005885 00000 н. 0000005906 00000 н. 0000006027 00000 н. 0000006048 00000 н. 0000006172 00000 н. 0000006287 00000 н. 0000006308 00000 н. 0000006432 00000 н. 0000006453 00000 п. 0000006577 00000 н. 0000006598 00000 н. 0000006722 00000 н. 0000007837 00000 п. 0000008123 00000 н. 0000008144 00000 н. 0000008268 00000 н. 0000008554 00000 н. 0000008575 00000 н. 0000008697 00000 п. 0000008718 00000 н. 0000008739 00000 н. 0000008863 00000 н. 0000008884 00000 н. 0000009008 00000 н. 0000009029 00000 н. 0000009154 00000 н. 0000009175 00000 н. 0000009300 00000 н. 0000009321 00000 п. 0000009446 00000 н. 0000009467 00000 н. 0000009592 00000 н. 0000009613 00000 н. 0000009738 00000 п. 0000009759 00000 п. 0000009884 00000 н. 0000009905 00000 н. 0000010028 00000 п. 0000010049 00000 п. 0000010173 00000 п. 0000010194 00000 п. 0000010318 00000 п. 0000010339 00000 п. 0000010464 00000 п. 0000010485 00000 п. 0000010610 00000 п. 0000010631 00000 п. 0000010756 00000 п. 0000010777 00000 п. 0000010902 00000 п. 0000010923 00000 п. 0000011047 00000 п. 0000011068 00000 п. 0000011192 00000 п. 0000011213 00000 п. 0000011337 00000 п. 0000011358 00000 п. 0000011481 00000 п. 0000011503 00000 п. 0000011699 00000 п. 0000011722 00000 п. 0000017329 00000 п. 0000017352 00000 п. 0000018735 00000 п. 0000018758 00000 п. 0000022441 00000 п. 0000022464 00000 п. 0000024344 00000 п. 0000024367 00000 п. 0000027267 00000 н. 0000027290 00000 н. 0000030259 00000 п. 0000030282 00000 п. 0000033605 00000 п. 0000033628 00000 п. 0000036285 00000 п. 0000036308 00000 п. 0000039176 00000 п. 0000039199 00000 п.