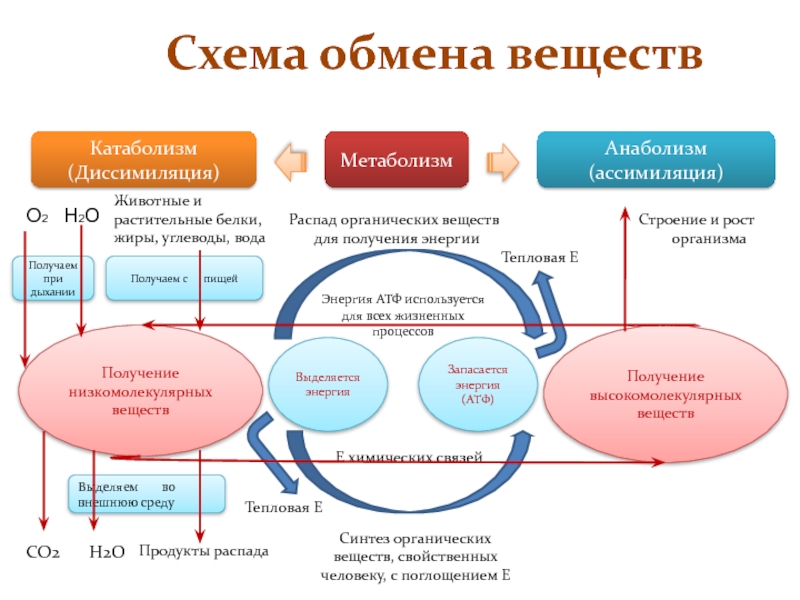
Обменные процессы в организме
Между организмом и окружающей его средой непрерывно происходят обменные процессы различных веществ и энергии.
Обмен веществ обычно идёт в два этапа:
первый этап ─ пластический обмен;
второй этап – энергетический обмен.
Пластический обмен начинается с поступления в организм воды и питательных веществ. В пищеварительном канале часть веществ расщепляется до более простых, они переходят во внутреннюю среду организма – кровь и лимфу.
С кровью вещества попадают в клетки, где происходят процессы их химических превращений. То есть из простых веществ синтезируются более сложные.
Главный смысл пластического обмена – это превращение поступивших веществ в вещества, необходимые организму.
Энергетический обмен
– второй этап обменных процессов в организме.
Потому что при расщеплении образовавшихся веществ выделяется энергия, необходимая для жизнедеятельности клеток, тканей и всего организма в целом.
В результате этих реакций выделяется не только энергия, но и образовываются такие вещества, как углекислый газ, аммиак и другие соединения фосфора, натрия, хлора, которые выводятся из организма.
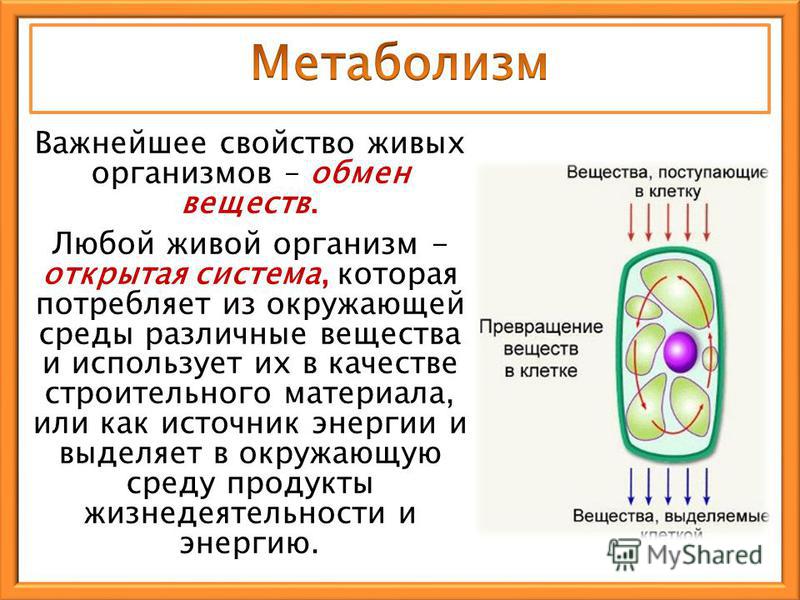
Обмен веществ — это набор химических реакций, которые возникают в живом организме для поддержания его жизнедеятельности. Эти процессы позволяют организмам расти, развиваться, размножаться, а также отвечать на воздействия окружающей среды.
Также можно сказать, что обмен веществ – это получение необходимых организму веществ из окружающей среды, преобразование их в организме и выведение продуктов распада, то есть ненужных веществ.
В результате обмена организм получает вещества для построения клеток и энергию для жизненных процессов.
Обменные процессы органических веществ (белков, жиров и углеводов), а также обменные процессы неорганических веществ (воды и минеральных солей)
Обмен белков в организме
С пищей в организм поступают «строительные материалы» ─ белки, которые содержатся в молочных, мясных, рыбных и других продуктах (их называют пищевыми белками). Поступив в желудок, они перевариваются, то есть расщепляются ферментом желудочного сока – пепсином.
Фермент — это белковая молекула, которая ускоряет химические реакции в организме.
Далее пищевые белки следуют в двенадцатиперстную кишку, где они расщепляются ферментом поджелудочной железы трипсином
Аминокислоты – это органические соединения, из которых состоит белок.
Аминокислоты через
кровеносные капилляры ворсинок кишечника поступают в печень. Там они
превращаются в жиры и углеводы, необходимые организму.
А в клетках аминокислоты строятся в белки тела.
Проще говоря, пищевые белки, попав в организм, расщепляются до аминокислот, а уже из аминокислот преобразовываются новые белки — белки тела, которые необходимы для определённых реакций.
Функции белков
Белки входят в состав ядер, цитоплазмы и мембран клеток.
Они являются ферментами.
Принимают участие в свёртывании крови и транспортировке кислорода.
Входят в состав кости.
При расщеплении белков выделяется энергия, необходимая организму.
Обмен жировЖиры содержатся в таких продуктах, как мясо, сало, жирные сорта рыбы, растительные масла, орехи и другие.
Попав в желудок, жиры расщепляются на глицерин и жирные кислоты.
В клетках кишечника из глицерина и жирных кислот синтезируется жир, характерный для организма.
Через лимфатическую
систему синтезированный жир направляется в жировое депо и клетки, где он
используется как запасное вещество и строительный материал.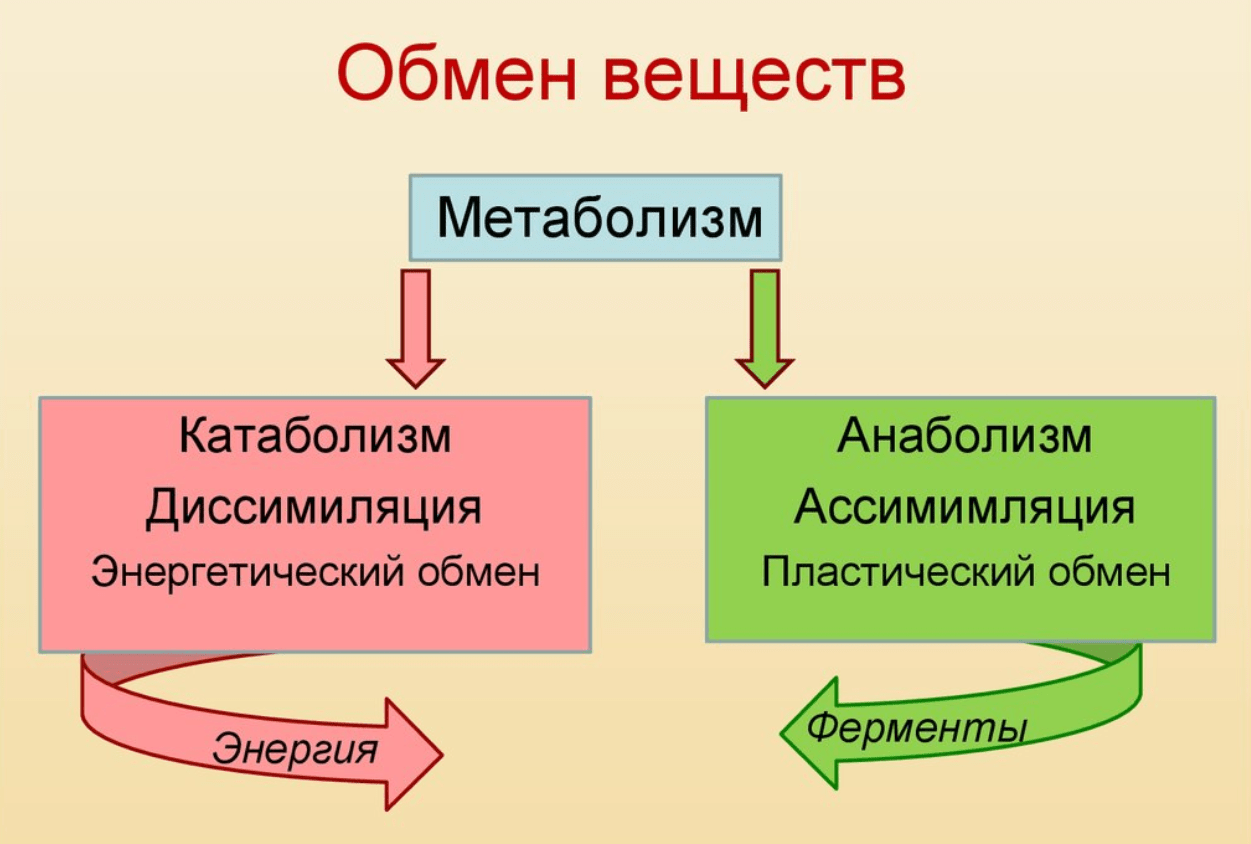
Функции жиров
Они входят в состав клеточных мембран.
В жирах растворяются некоторые витамины (жирорастворимые).
Из жиров образуются некоторые гормоны и биологически активные вещества.
Гормоны – это специальные химические посредники, которые регулируют работу организма и заставляют работать определённые клетки
Также жиры выполняют защитную функцию, предохраняя некоторые органы от сотрясения. Подкожный жир плохо проводит тепло, что способствует сохранению постоянной температуры тела.
Они являются богатым источником энергии. При расщеплении одного грамма жира освобождается в два раза больше энергии, чем при расщеплении одного грамма белков или углеводов.
При распаде жиров выделяется также вода, которая необходима для поддержания водного обмена.
Обмен углеводов
Углеводы содержатся во
многих растительных продуктах: в зёрнах злаков, сахаре, мёде, картофеле, ягодах
и фруктах.
Некоторые углеводы начинают распадаться в ротовой полости под действием фермента слюны – амилазы.
В двенадцатиперстной кишке под действием ферментов поджелудочной железы углеводы расщепляются до глюкозы и других простых углеводов. Глюкоза и простые углеводы всасываются в тонком кишечнике, попадают в кровь и направляются в печень.
Там они превращаются в гликоген и другие соединения, которые распределяются между клетками тела.
Гликоген – это основной запасной углевод животных.
Он образует энергетический резерв, который может быстро восполнить внезапный недостаток глюкозы в крови.
Уровень глюкозы в крови постоянный, он регулируется гормоном поджелудочной железы – инсулином.
Под действием инсулина избыток глюкозы в крови превращается в гликоген, запасы которого откладываются в печени и мышцах.
При недостатке гормона
инсулина возникает тяжёлое заболевание – сахарный диабет, при котором
избыточная глюкоза накапливается в крови.
А при недостатке глюкозы в крови гликоген превращается в глюкозу под действием гормона поджелудочной железы – глюкагона.
Функции углеводов в организме человека:
· энергетическая функция.
В качестве основного энергетического источника в организме используется свободная глюкоза или запасные углеводы в виде гликогена.
Головной мозг может нормально функционировать только в том случае, если к нему в качестве энергетического материала поступает глюкоза;
· транспортная функция. Углеводы участвуют в передаче наследственной информации, в составе нуклеиновых кислот, таких как ДНК и РНК.
Нуклеиновые кислоты ДНК и РНК присутствуют в клетках всех живых организмов и выполняют важнейшие функции по хранению, передаче и реализации наследственной информации;
·
защитная
фикция. Сложные углеводы входят в состав компонентов иммунной системы и
защищают от проникновения бактерий и вирусов, а также от механических
повреждений.
Процессы обмена веществ в организме взаимосвязаны. Белки могут превращаться в жиры и углеводы, а некоторые углеводы в жиры. Жиры могут стать источником углеводов, а недостаток углеводов может пополниться за счёт жиров и белков.
Однако, недостаток белков в организме не восполним, так как белки строятся из аминокислот, а аминокислоты не все образуются в организме.
Все обменные процессы органических веществ (белков, жиров и углеводов), а также их биохимические реакции проходят в водной среде.
Обмен неорганических веществ воды и минеральных солей
Основную часть таких жидкостей тела человека, как кровь, лимфа, пищеварительные соки составляет вода. Она содержится в ядрах клеток и цитоплазме.
Внутренняя среда человека
содержит до 90 % воды. Вода в организме либо химически связана с другими
соединениями, либо содержит в себе растворенные минеральные соли и органические
вещества.
Транспорт питательных веществ и кислорода, а также выход продуктов распада осуществляется благодаря наличию воды.
Таким образом, в организме поддерживается баланс между поступающей и выделяемой водой.
Чтобы не допустить обезвоживание, необходимо ежедневно восполнять потери воды. В среднем взрослому мужчине необходимо около 3 литров жидкости − это 15 стаканов объёмом 200 миллилитров. Таковы рекомендации института питания. Причём чистой питьевой воды необходимо пить около 8 стаканов, остальное количество приходится на суп, чай, компот и прочее.
Почему нужно пить именно воду?
Дело в том, что организму
необходимо совершать дополнительные усилия, чтобы выделить чистую воду, скажем,
из чая или сока. Потребляя простую воду, вы помогаете своему телу, избавляете
его от лишних усилий по добыванию чистой воды. Чай, кофе, молоко, компот и соки
организм воспринимает как пищу. В древности даже говорили не «попить молока», а
«поесть молока».
В древности даже говорили не «попить молока», а
«поесть молока».
Обменные процессы неорганических веществ – минеральных солей
Так, например, соли кальция необходимы в процессе свёртывания крови, калий и кальций участвуют в механизме мышечного сокращения.
Минеральные соли составляют основу межклеточного вещества костной ткани.
В состав минеральных солей входят различные элементы, их подразделяют на макро— и микроэлементы.
К макроэлементам относят: кальций, калий, натрий, фосфор, хлор и другие.
К микроэлементам относят: железо, кобальт, цинк, фтор, йод и другие.
Сохранение водно-солевого равновесия очень важно для организма.
Если в крови и тканевой
жидкости концентрация соли увеличивается, то вода начинает выходит из клеток, в
результате чего клетки обезвоживаются.
А если в крови и тканевой жидкости концентрация соли уменьшается, то вода, наоборот, поступает в клетки, в результате чего клетки начинают увеличиваться, а их работа нарушается.
Минералы содержатся во всех продуктах, поэтому все необходимые организму минеральные вещества можно получить, если употреблять разнообразную пищу.
В противном случае стоит использовать поливитаминные комплексы, в состав которых входят не только витамины, но и минеральные вещества.
Как лечить нарушение обмена веществ?
« Назад04.09.2019 00:00
Обмен веществ во многом определяет состояние нашего здоровья. Замедление обмена веществ ведет к постепенному увеличению веса, ухудшению состояния кожи, волос и общего самочувствия.
Ухудшение обмена веществ нередко связано с неправильным питанием, поэтому в первую очередь следует пересмотреть свой рацион. Все прекрасно знают о пользе клетчатки, которая очищает наш кишечник и выводит токсины. Особенно полезны в этом плане отруби, морковь и свекла.
Все прекрасно знают о пользе клетчатки, которая очищает наш кишечник и выводит токсины. Особенно полезны в этом плане отруби, морковь и свекла.
А вот большое количество пищи животного происхождения приводит к нарушениям обменных процессов и ожирению, потому что организм тяжелее и дольше справляется с такой пищей. Чтобы дать организму восстановиться, иногда надо проводить разгрузочные дни.
Как лечить нарушение обмена веществ народными методами?
1. Облепиховое масло.
Регуляцию обмена веществ хорошо восстанавливает облепиховое масло. Кроме этого оно имеет противоопухолевые свойства. Чтобы приготовить масло из ягод облепихи, их надо промыть и просушить. После этого ягоды надо подавить, насыпать в емкость и залить растительным маслом, чтобы оно немного покрыло ягоды. Оставьте настояться в темном месте одну неделю при периодическом перемешивании. Готовый настой отжать, процедить и оставить еще настояться до прозрачности. Затем масло слить в емкость с непрозрачным стеклом, держать в холодильнике. Перед употреблением облепиховое масло разводят растительным маслом в пропорции 1:5.
Перед употреблением облепиховое масло разводят растительным маслом в пропорции 1:5.
2. Йод.
Йод является обязательным условием для нормального протекания обменных процессов. Так как во многих районах нашей страны йода не хватает в продуктах, то следует добавлять в питание морепродукты и морскую капусту, хурму и редьку. А пищу присаливать йодированной солью.
3. Лечебное голодание.
Обмен веществ хорошо восстанавливается после проведения процедур очищения. Сюда относится в первую очередь лечебное голодание, во время которого организм вынужден начать переваривать собственные ткани. Однако в первую очередь он начнет использовать самое ненужное, куда входят отложения солей, лишний жир, кисты, жировики, чужеродные патологические новообразования. Такой процесс начинается от того, что усиливается ферментативная и энзимная активность лейкоцитов.
К примеру, голодание длительностью в 36 ч увеличивает их активность в 3 раза, чего уже достаточно для очищения организма. Более длительные голодовки приводят к полному обновлению и омоложению организма, а также к усилению обмена веществ. После лечебного голодания первый месяц наблюдается улучшение обмена веществ на 5-6 % и лучшее усвоение пищи. Это объясняется отдыхом во время голодания всех органов, восстановлением поврежденных структур и избавлением от шлаков.
Более длительные голодовки приводят к полному обновлению и омоложению организма, а также к усилению обмена веществ. После лечебного голодания первый месяц наблюдается улучшение обмена веществ на 5-6 % и лучшее усвоение пищи. Это объясняется отдыхом во время голодания всех органов, восстановлением поврежденных структур и избавлением от шлаков.
4. Рецепт тибетских лекарей.
Этот сбор улучшает обмен веществ, приводит к очищению организма и омоложению. Для него необходимо по 100 г травы зверобоя продырявленного, березовых почек, бессмертника песчаного, цветков ромашки аптечной.
Чтобы его приготовить надо все составляющие измельчить с помощью мясорубки. Затем отделить 1 ст. л. сбора и залить вечером пол-литром кипятка, настоять 20 минут и процедить. Пить 1 стакан настоя перед сном с добавлением 1 ч. л. меда. После выпитого настоя нельзя ничего пить и есть. А утром следует немного подогреть стакан оставшегося настоя и выпить натощак за полчаса до еды с 1 ч. л. меда. Пить отвар до окончания первоначальных компонентов. Курс можно повторять раз в 5 лет.
л. меда. Пить отвар до окончания первоначальных компонентов. Курс можно повторять раз в 5 лет.
Как видите, рекомендации по улучшению обмена веществ, несложные. Главное, как считает Здоровая жизнь, желать помочь своему организму быть здоровым.
Источник
Обменные процессы — Справочник химика 21
Ядерный магнитный резонанс (ЯМР) и электронный парамагнитный резонанс (ЭПР) — два метода радиоспектроскопии, позволяющие изучать структуру и динамику молекул, радикалов, ионов в конденсированных и газовой фазах вещества. Спектры ЯМР обладают высокой специфичностью и широко применяются для идентификации соединений, в структурно-аналитических целях, а также для изучения быстрых обменных процессов. Спектроскопия ЭПР — метод исследования парамагнитных частиц и центров, кинетики и механизмов процессов, происходящих с их участием. Особенно большой прогресс в развитии методов спектроскопии ЯМР и ЭПР, достигнутый в последние годы, связан с появлением импульсных фурье-спектрометров, двухмерной спектроскопии и техники множественного ядерного, электрон-ядерного и электрон-электрон-ного резонанса.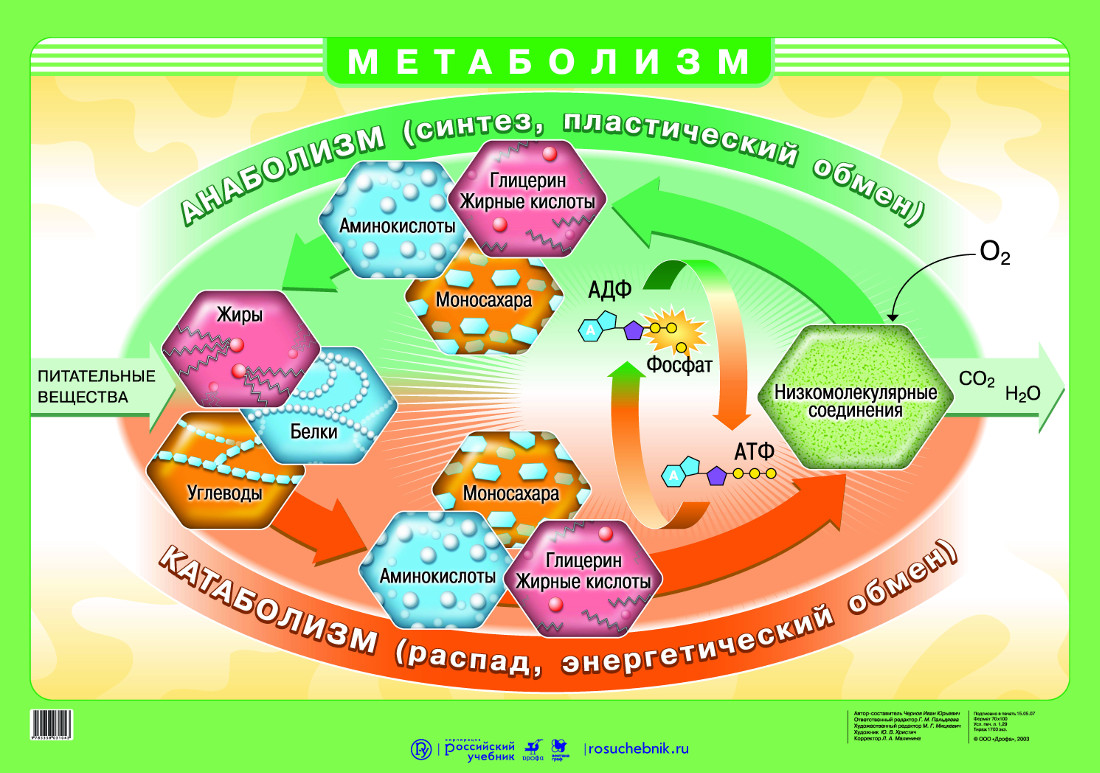 [c.5]
[c.5]
Количественные гидродинамические характеристики насадочных колонн ниже точки инверсии. К важнейшим параметрам гидродинамической структуры потоков в насадке ниже точки инверсии относятся перепад давления в насадке, отношение скорости газа (пара) к скорости в инверсионной точке, длительность пребывания потоков в аппарате, доля эффективно используемого объема системы, степень продольного перемешивания в колонне, характер и интенсивность обменных процессов в жидкой, газообразной (паровой) фазах и т. п. [c.394]
Н х, t) S — переменный объем п-й ячейки потока газа /сг, i, / 2 — коэффициенты обмена замкнутой цепи обменных процессов между ячейками. Частные случаи модели (7.140) не раз встречались в литературе. Так, если пренебречь распределенностью гидродинамических параметров по длине аппарата и во времени, а также обменом между проточными и застойными зонами в жидкости, то система уравнений (7. 140) примет вид, который исследовался в работе [49] [c.418]
140) примет вид, который исследовался в работе [49] [c.418]
При установившемся равновесии обменного процесса поверхность ионита и раствор приобретают электрические заряды противоположного знака, на границе раздела ионит — раствор возникает двойной электрический слой, которому соответствует скачок потенциала. Поскольку иониты обладают повышенной избирательной способностью по отношению к определенному виду ионов, находящихся в растворе, ионообменные электроды называются также ионоселективными. Стеклянный электрод является важнейшим среди этой группы электродов. Он представляет собой тонкую мембрану из специального стекла, в котором повышено содержание щелочных составляющих — соединений натрия, лития и др. Согласно теории Б. П. Никольского потенциалопределяющий процесс на границе раствор — стекло заключается в обмене между ионами щелочного металла, например Ма+, содержащимися в стекле, и ионами Н+, находящимися в растворе [c.484]
Эффективность полученных результатов после ступенчатой обработки смешанного стока объясняется тем, что после каждой ступени обработки в дисперсионной среде уменьшается концентрация сольвара и вследствие сдвига обменных процессов между коллоидной и растворенной частью стока и частью, находящейся в виде коагулированных агрегатов.
 [c.106]
[c.106]Поскольку механизм действия при мицеллярно-полимерном воздействии в нефтяных пластах в большой степени определяется сорбционными и обменными процессами, требуется глубокое изучение сложной физико-химической основы метода. Можно предвидеть, что при промышленном испытании и последующем внедрении этой технологии неизбежны большие трудности методического и технологического порядка. Эти трудности будут заключаться в количественной оценке механизма процессов и его учете при проектировании, в правильном подборе состава мицеллярных растворов для конкретных физико-геологических условий месторождений (минералогического состава пластовой воды и породы), определении оптимального размера оторочки мицеллярного раствора и оторочек полимерного раствора, технологии нагнетания этих растворов, методах прогноза и проектирования процесса, оценки эффективности и т. д. [c.167]
Специалистам-нефтяникам становится все более ясно, что сорбционные явления н обменные процессы в пластах играют существенную роль в механизме нефтегазоотдачи, особенно при примене-нни нетрадиционных методов воздействия на залежи. Задача их изучения, качественный и количественный учет при технологических решениях — актуальный вопрос в науке и практике нефтедобычи. [c.171]
Задача их изучения, качественный и количественный учет при технологических решениях — актуальный вопрос в науке и практике нефтедобычи. [c.171]
Существует определенная разница между естественной и искусственной сушкой сена. При луговой естественной сушке травы на сено происходит ряд ферментационных превращений, в том числе и под влиянием ультрафиолетового облучения, а при искусственной сушке горячим воздухом ничего подобного не наблюдается. Искусственная тепловая обработка способствует сохранению в сухом продукте протеина, витаминов и крахмала, поскольку в этом случае тормозятся любые обменные процессы, ведущие к сокращению запасов полезных веществ. Сено грану- [c.342]
Обработка табака и каучука-сырца. Два вида тропических сельскохозяйственных технических культур — табак и каучук — обязательно должны пройти дегидратацию. Каучук-сырец проходит многократную сушку. Табак во время сушки подвергается управляемой обработке, при которой в табачном листе происходят обменные процессы. В обоих случаях можно использовать горячий воздух, однако все более широкое распространение начинает находить процесс прямой сушки продуктами сгорания, что позволяет регулировать и значительно ускорять процесс сушки. [c.343]
В обоих случаях можно использовать горячий воздух, однако все более широкое распространение начинает находить процесс прямой сушки продуктами сгорания, что позволяет регулировать и значительно ускорять процесс сушки. [c.343]
Влияние гидроокиси натрия на показатели набухания бентонита и Рт (табл. 26) при малых концентрациях объясняется пептизацией, а при высоких — коагуляцией. Есть основания полагать, что при контакте с глинистыми минералами наряду с обменными процессами происходит частичное молекулярное поглощение щелочи — хемосорбционный процесс. [c.60]
Спектроскопия ЯМР широко и успешно применяется для исследования равновесных химических превращений и обменных процессов, при которых периодически меняется строение, а значит, электронное окружение магнитных ядер и спин-спиновое взаимодействие ядер, т. е. химические сдвиги б и константы /. К таким процессам относятся как внутримолекулярные превращения (заторможенное внутреннее вращение, инверсия пирамидальной системы связей у азота, инверсия циклов, таутомерия и т.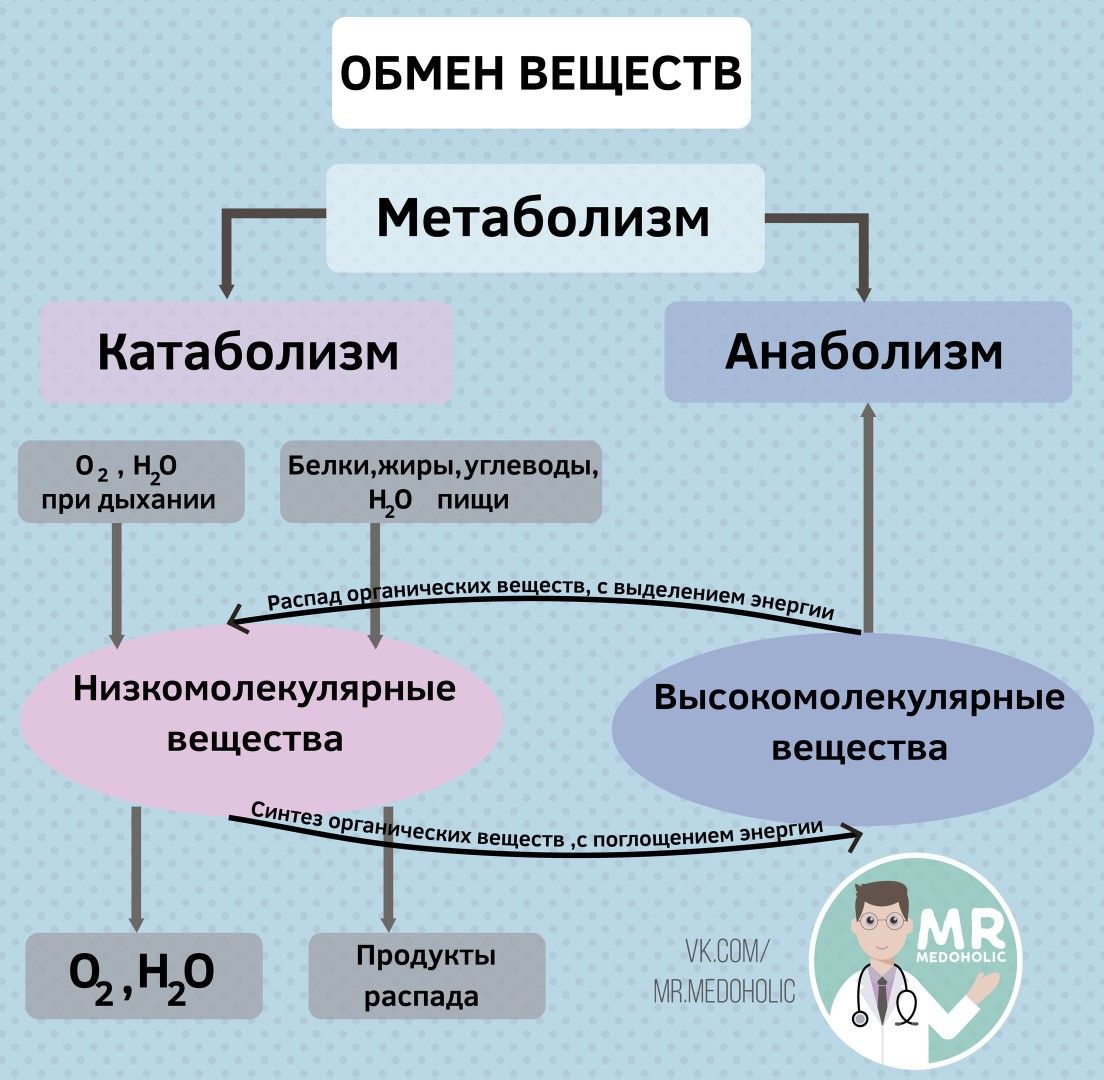 д.), так и межмо-лекулярные обменные и другие равновесные химические реакции (протонный обмен в водных растворах карбоновых кислот, аммиака, лигандный обмен, рекомбинация ионов, биохимические взаимодействия фермент — субстрат и т. д.). [c.40]
д.), так и межмо-лекулярные обменные и другие равновесные химические реакции (протонный обмен в водных растворах карбоновых кислот, аммиака, лигандный обмен, рекомбинация ионов, биохимические взаимодействия фермент — субстрат и т. д.). [c.40]
Предосторожности при приготовлении растворов. При приготовлении растворов нужно иметь в виду, что на стекле возможны обменные процессы, в результате которых очень малые количества элементов могут полностью исчезнуть из раствора, а вместо них появляются ионы натрия, кремниевая кислота и другие примеси, перешедшие из стекла. Такое же явление может происходить и при выпаривании очень разбавленных нейтральных растворов. Для длительного хранения следует приготовлять более концентрированные растворы. [c.21]
Применяется при нарушениях белкового обмена и как общий стимулятор обменных процессов (например, при заболеваниях печени) [c.252]
Явление самопроизвольного переноса растворителя через полупроницаемую перегородку называют осмосом. Он сопровождает многие биологические процессы и находит широкое применение в лабораторной практике. Благодаря ему происходят обменные процессы в растениях и животных организмах. А в производстве полимеров на использовании осмоса основан один из важнейших методов контроля (см. разд. VI.20). [c.208]
Он сопровождает многие биологические процессы и находит широкое применение в лабораторной практике. Благодаря ему происходят обменные процессы в растениях и животных организмах. А в производстве полимеров на использовании осмоса основан один из важнейших методов контроля (см. разд. VI.20). [c.208]
Применение методов спектроскопии ЭПР в химических исследованиях весьма разнообразно. Но грубо можно говорить о двух направлениях— одном, касающемся в основном структурных аспектов, и другом — динамики процессов. К первому относится изучение структуры органических, неорганических и комплексных радикалов и ион-радикалов, парамагнитных центров в твердых телах и т. д., а ко второму — изучение механизмов и кинетики химических реакций, обменных процессов и т. д. [c.68]
Формулой (11.9) обычно и пользуются для оценок УоЛ. АО по константам скоростей обменных процессов, найденным методом динамической спектроскопии ЯМР. [c.41]
[c.41]
В невырожденных системах, если содержание одной из форм менее 1% (хвразность энергий форм ДО (или, грубо, Д1 —разность глубины потенциальных ям) более — 15 кДж/моль, то чувствительность метода динамического ЯМР становится недостаточной для фиксирования обменного процесса. Оптимально его применение к изучению состояний при сравнимых заселенностях. [c.43]
Для сужения сигналов ЭПР на практике часто приходится прибегать к сильному охлаждению образцов жидким азотом или даже гелием, или водородом, что прежде всего позволяет увеличить время спин-решеточной релаксации. Это особенно бывает необходимо при изучении солей переходных металлов и редкоземельных элементов. Для снижения эффектов, вызываемых спин-спиновой релаксацией и обменными процессами, прибегают также к разбавлению образцов диамагнитными веществами и изоляции парамагнитных центров друг от друга в матрицах и при замораживании растворов. [c.66]
Зависимость, приведенная для коэффициента турбулентного обмена, аналогична зависимости для коэффициента молекулярной диффузии D= 3lav, где /о—длина пути свободного пробега молекулы, а и — средняя скорость молекулы. Если I не превосходит глубину фронта пламени в ламинарном потоке бн, то поверхность пламени должна остаться гладкой , однако, как оказалось, и в этом случае наличие турбулентности интенсифицирует обменные процессы. Величина 5н равна примерно 1 мм. Теория рассматривает поверхностное горение турбулентных объемов газа, когда 1объемное горение, когда [c.166]
Если I не превосходит глубину фронта пламени в ламинарном потоке бн, то поверхность пламени должна остаться гладкой , однако, как оказалось, и в этом случае наличие турбулентности интенсифицирует обменные процессы. Величина 5н равна примерно 1 мм. Теория рассматривает поверхностное горение турбулентных объемов газа, когда 1объемное горение, когда [c.166]
Обменные процессы в условиях МФК идут даже при конкурентном образовании карбенов из равных молярных количеств бромоформа и хлороформа например, реакция со стиролом в специфических условиях в присутствии системы гидроксид натрия/ТЭБА приводит к образованию 78% аддукта СВгг, 15% аддукта ССЬ и 7% аддукта Br l [384]. В отсут- [c.352]
Химические обменные процессы заметно изменяют ширину линий. Этот эффект также можно ослабить путем разведения. Если в обмене участвуют эквивалентные парамагнитные частицы, то линии уширяются у основания и становятся уже у центра. Если в обмене участвуют различные ионы, то отдельные линии сливаются и дают один сигнал, который может быть широким или узким в зависимости от скорости обмена. Такой эффект наблюдается для uS04-5h30, в элементарной ячейке которого имеются два различных центра меди [2]. [c.205]
Такой эффект наблюдается для uS04-5h30, в элементарной ячейке которого имеются два различных центра меди [2]. [c.205]
Из проведенного ранее обсуждения химических сдвигов ионизационных пиков РФС электронов оболочки можно сделать вывод, что для электронов оболочки всегда наблюдаются простые спектры, например, для каждого заметно различающегося окружения атома азота наблюдается один пик для Ь-электронов азота. К счастью, зто не всегда так [27]. Мы уже видели, что парамагнитные частицы, такие, как О2, вызывают обменные расщепления линий электронов оболочки. Такие же расщепления, обусловленные обменными процессами, обнаружены и в спектрах РФС парамагнитных комплексов ионов переходных металлов. Кларк и Адамс [60] сообщили о Зх-обменном расщеплении хрома величиной около 4,5 эВ в Сг(ЬГа)з и 3,1 эВ в Сг(Ь -С5Н5)2. Может возникнуть вопрос, должен ли анализ такого расщепления способствовать пониманию деталей контактных сдвигов Ферми в ЯМР, наблюдаемых для парамагнитных частиц. [c.353]
[c.353]
Учитывая, что одной из основных задач фундаментальных исследований проблемы увеличения нефтеотдачи пластов является поиск принципиально новых методов и химреагентов для извлечения нефти из недр, нами разработан новый метод извлечения остаточной нефти, основанный на принципе взаимодействйя комплексообразующих химреагентов с полярными нефтяными компонентами. Метод основан на воздействии химреагентов на металло-порфирины нефти, что приводит к разрушению асфальтосмолистых структур. Установлено, что при воздействии поли-функциональных реагентов на нефть на границе нефть — вода происходят обменные процессы между ассоциатами нефти и химическими добавками, что приводит к разрушению структуры, снижению вязкости нефти и к повышению нефтеотдачи пласта.Наиболее эффективными в этом плане являются азот-, фосфор- и кислородсодержащие реагенты, растворимые в воде. В работе представлены результаты комплексного изучения механизма взаимодействия относительно недорогих комплексооб разующих реагентов с нефтями различных месторождений, приводящие к изменению их физико-химических свойств. На основе исследований разработаны [c.4]
На основе исследований разработаны [c.4]
Непористые реакционно-диффузионные мембраны отличаются от прочих химической формой связи компонентов разделяемой смеси и исходного материала мембраны. Химические реакции приводят к образованию новых веществ, участвующих в транспорте целевого компонента. Массоперенос компонентов разделяемой газовой смеси определяется не только внешними параметрами и особенностями структуры матрицы, но и химическими реакциями, протекающими в мембране. В подобных системах за счет энергетического сопряжения процессов диффузии и химического превращения возможно ускорение или замедление мембранного переноса, в определенных условиях возникает активный транспорт, т. е. результирующий перенос компонента в направлении, противоположном движению под действием градиента химического потенциала этого компонента. В сильнонеравновесных мембранных системах могут формироваться структуры, в которых возникают принципиально иные механизмы переноса, например триггерный и осциллирующий режимы функционирования мембранной системы. Обменные процессы такого рода обнаружены в природных мембранах, но есть основания полагать, что синтетические реакционно-диффузионные мембраны в будущем станут основным типом разделительных систем, в частности, при извлечении токсичных примесей из промышленных газовых выбросов. [c.14]
Обменные процессы такого рода обнаружены в природных мембранах, но есть основания полагать, что синтетические реакционно-диффузионные мембраны в будущем станут основным типом разделительных систем, в частности, при извлечении токсичных примесей из промышленных газовых выбросов. [c.14]
Высказывавшиеся ьшою ранее [5381 представления о возможном об-» разевании нефтяных СС с помощью рассмотренных обменных процессов в свете современных данных, конечно, устарели.— Прим. ред. [c.75]
Математическую модель нестационарного процесса абсорбции в насадочном аппарате построим так, чтобы она отражала три основных фактора, наиболее важных в общем динавлическом поведении процесса 1) неравномерность распределения по времени пребывания элементное потока в аппарате, 2) распределенность в пространстве и времени основных гидродинамических параметров процесса удерживающей способности, расхода жидкости в колонне, перепада давления, 3) наличие полной замкнутой цепи обменных процессов в насадочном аппарате газовая фаза—проточная зона потока жидкости—застойная зона потока жидкости—газовая фаза с количественным выражением интенсивности обменных процессов всех звеньев замкнутой цепи. [c.415]
[c.415]
Мы видим, что в обоих случаях Д аксп > о. Аналогичный результат получается и для обменного процесса типа [c.146]
Таким образом, согласно теории активированного комплекса, при представлении константы скорости обменного процесса в аррениусовской форме экспериментальная эпергин активации увеличивается с повышением температуры. Однако это изменение энергии активации относительно невелико и в пределах погрешностей измерений обычно не обнаруживается. [c.146]
Приведенный механизм был доказан введением СОз радикалов. Прн 300 °С изотопный состав пропилена следующий 30% СдНе, 20 о СзНзО, 30% СзН4В2 и 20% СзНзОд. Это указывает на наличие обменных процессов. [c.195]
Разновидностью адсорбции является ионный обмен — процесс оазделения, основанный на способности некоторых твердых веществ (ионитов) обменивать свои подвижные ионы на ионы растворов электролитов. [c.382]
Биолог. Мне известно, что инсулин, используемый при лечении больных сахарным диабетом, не только ускоряет процессы утилизации глюкозы клетками, но и значительно интенсифицирует ряд других обменных процессов в организме, включая процессы синтеза белка в сердечной мьтще, а также в жировой ткани, печени и скелетных мьшщах [Руководство по физиологии, 1982], Если это так, то введение инсулина должно способствовать повьппению Параметра Подобия. Может бьггь, в этом и состоит причина наблюдаемого вами улучшения состояния инфарктных больных после назначения им инсулина [c.89]
Мне известно, что инсулин, используемый при лечении больных сахарным диабетом, не только ускоряет процессы утилизации глюкозы клетками, но и значительно интенсифицирует ряд других обменных процессов в организме, включая процессы синтеза белка в сердечной мьтще, а также в жировой ткани, печени и скелетных мьшщах [Руководство по физиологии, 1982], Если это так, то введение инсулина должно способствовать повьппению Параметра Подобия. Может бьггь, в этом и состоит причина наблюдаемого вами улучшения состояния инфарктных больных после назначения им инсулина [c.89]
Биолог. И вот на что еще я хочу обратить ваше внимание. Известно, что понижение средней интенсивности обменных процессов в организме приводит к уменьшению числа митохондрий в клетках, а значкг, и Н-параметра. Сами же митохондрии очень похожи на аэробные бактерии [Кемп, Арме, 1988], а к ним относится и холерный вибрион. Поэтому снижение //-параметра, или Живой Температуры Организма, уменьшает опасность возникновения холеры, брюшного тифа и других инфекционных болезней, но увеличивает опасность возникновения рака, сердечнососудистых и вирусных заболеваний. .. [c.119]
.. [c.119]
Представляется перспективным найденный новый метод из-илечения остаточной нефти, основанный на принципе взаимодействия комплексообразующих агентов с полярными нефтяными компонентами,В результате значительно возрастает эффективность нефтеизвлечения. Метод основан на химическом воздействии иа металлопорфирины нефти, что приводит к разрушению асфальто-смолистых структур. При воздействии химических реагентов на нефть на границе нефть — вода происходят обменные процессы между ассоциатами в нефти и химическими добавками. В связи с этим на практике становится возможным применение водных растворов химических реагентов, способных взаимодействовать с металлопорфиринами, переводя их из нефтяной фазы в водную.Наиболее эффективными в этом плане являются полифунк-циональные азот-, фосфор-, кислородсодержащие реагенты, растворимые I воде. [c.105]
Если время релаксации велико, то заселенность верхнего уровня будет возрастать, а интенсивность сигнала ЭПР падать из-за насыщения. При малом времени релаксации линия будет широкой из-за принципа неопределенности. Уширяют сигнал и нерелаксационные процессы, в частности тонкое и сверхтонкое спин-спиновое взаимодействие (см. выше), обменные процессы и др. Что касается обменных процессов, то принципы эффекта являются общими для спектроскопии ЭПР и ЯМР и обсуждались в гл. I, однако при рассмотрении спектров ЭПР должен учитываться не только обмен ядер, но и обмен электронов. [c.66]
При малом времени релаксации линия будет широкой из-за принципа неопределенности. Уширяют сигнал и нерелаксационные процессы, в частности тонкое и сверхтонкое спин-спиновое взаимодействие (см. выше), обменные процессы и др. Что касается обменных процессов, то принципы эффекта являются общими для спектроскопии ЭПР и ЯМР и обсуждались в гл. I, однако при рассмотрении спектров ЭПР должен учитываться не только обмен ядер, но и обмен электронов. [c.66]
Как всякий физический метод, мессбауэровская спектроскопия имеет свое характеристическое время. Если изучаемое соединение само меняется во времени, т. е. происходят, например, обменные процессы или переходы одной формы соединения в другую, то при этом могут, естественно, меняться как изомерный сдвиг, так и градиент электрического поля на ядре (квадрупольное расщепление) и внутренние магнитные поля. Поэтому важно, как соотносятся времена жизни разных форм образца (частота их перехода) и характеристическое время метода. Разность частот мес-сбаузровских пере.ходов источника и образца имеет порядок 10 с (порядок величины частотной характеристики изомерного сдвига см. выще). Для того чтобы можно было наблюдать отдельные сигналы и измерять изомерные сдвиги для двух переходящих друг в друга форм образца, частота их перехода не должна превышать 10 С». Таким образом, порядок характеристического времени метода мессбауэровской спектроскопии можно оценить как 10 с. [c.127]
Разность частот мес-сбаузровских пере.ходов источника и образца имеет порядок 10 с (порядок величины частотной характеристики изомерного сдвига см. выще). Для того чтобы можно было наблюдать отдельные сигналы и измерять изомерные сдвиги для двух переходящих друг в друга форм образца, частота их перехода не должна превышать 10 С». Таким образом, порядок характеристического времени метода мессбауэровской спектроскопии можно оценить как 10 с. [c.127]
В комплексе железа со смешанной валентностью, т. е. атомами Ре(И) и Ре(111), в спектре будут наблюдаться два сигнала, только если любой переход электрона между этими атомами будет происходить достаточно медленно. Например, в комплексном соединении [Ре Ре2 0(СНзС00)б(Н20)з] при температуре 290 К в мессбауэровском спектре наблюдается один усредненный синглетный сигнал (рис. У.9), указывающий на протекание быстрого обменного процесса (высокочастотный переход электрона). При понижении температуры этот сигнал постепенно расщепляется, причем наиболее четко структура сигналов, указывающих на наличие двух неэквивалентных атомов Ре, проявляется лишь при 17 К. Сделан вывод о переходе электрона в пределах фрагмента РезО, а энергия активации оценена в 470 см . [c.127]
Сделан вывод о переходе электрона в пределах фрагмента РезО, а энергия активации оценена в 470 см . [c.127]
Миокардиодистрофия
Определение
Миокардиодистрофия — невоспалительное поражение сердечной мышцы в результате нарушений ее метаболизма под влиянием внесердечных факторов.
Этиология и патогенез
Острые и хронические экзогенные интоксикации, в том числе алкогольная (см.),
эндокринные и обменные нарушения (тиреотоксикоз, микседема, синдром Кушинга, ожирение, сахарный диабет, авитаминозы, голодание),
анемии,
патологический климакс,
физические перенапряжения («солдатское сердце»),
инфекции (в том числе тонзиллярная),
физические агенты (радиация, невесомость, перегревание),
системные заболевания (коллагенозы, нейромышечная дистрофия и др.).
К миокардиодистрофиям относят также отложение в миокарде патологических продуктов обмена или нормальных метаболитов: амилоида, железа (при гемохроматозе), гликогена и др.Некоторые формы миокардиодистрофии нередко называют также миокардиопатиями (алкогольная, климактерическая, амилоидная).
Патогенез. Изменение течения биохимических процессов в миокарде с последующим нарушением микроструктур и сократительной функции мышечных волокон. Эти изменения до определенного предела обратимы.
 Некоторые формы миокардиодистрофии нередко называют также миокардиопатиями (алкогольная, климактерическая, амилоидная).
Некоторые формы миокардиодистрофии нередко называют также миокардиопатиями (алкогольная, климактерическая, амилоидная).Клиническая картина
Симптомы нередко маскируются признаками основного заболевания. Отмечается повышенная утомляемость, небольшая одышка при физических напряжениях, приглушение I тона сердца на верхушке, иногда умеренная тахикардия. При анемии часто выслушивается систолический шум над верхушкой и легочной артерией, при тиреотоксикозе нередки боли в области сердца, выраженная тахикардия, повышение систолического и пульсового давления, мерцательная аритмия. При микседеме — одышка, редкий пульс, гипотензия, глухость сердечных тонов, рано появляются признаки сердечной недостаточности. При прогрессирующем течении миокардиодистрофии любой этиологии развивается сердечная недостаточность, возникают нарушения сердечного ритма.
Диагностика
Электрокардиография, эхокардиография, ангиография, пункция миокарда.
Лечение
Устранение основного патологического процесса, вызвавшего миокардиодистрофию. Регулирование режима труда, предупреждение физических нагрузок, рациональное питание (коррекция обменных нарушений), санация хронических очагов инфекции. Назначают средства, способствующие нормализации обмена в миокарде.
Прогноз
Прогноз при адекватной терапии благоприятный.
Кедровая бочка
Кедровая бочка (фитосауна)-помогает избавиться от многих недугов, омолаживает организм. Сочетание пара, насыщенного растительными травами, который подается в кабину из сибирского кедра, благотворно действует на человека. Кедр считается древом — фармацевтом. Его древесина выделяет фитонциды, которые убивают и подавляют рост и развитие вредных микроорганизмов.
Эффект, получаемый от использования «кедровой бочки»:
- активация антистрессовых механизмов
- комплексная коррекция фигуры
- коррекция нарушений обмена веществ и эндокринной системы
- улучшение циркуляции крови
- восстановление и повышение половой функции
- профилактика и лечение простудных заболеваний
- профилактика преждевременного старения
- иммуномодулирующее действие
- лечение функциональных заболеваний нервной системы
- лечение заболеваний опорно – двигательного аппарата
- прекрасная миорелаксация перед сеансом массажа
Под воздействием горячего насыщенного целебными травами пара кожные поры открываются, происходит активное потоотделение. Обильно конденсирующиеся на коже капли пара, стекая по телу, смывают выделившиеся наружу вредные вещества, не давая им снова всосаться обратно. Таким образом, происходит очистка организма, — общая и на клеточном уровне, стимулируется работа всех органов и систем, нормализуется обмен веществ, очищается и омолаживается кожа.
Обильно конденсирующиеся на коже капли пара, стекая по телу, смывают выделившиеся наружу вредные вещества, не давая им снова всосаться обратно. Таким образом, происходит очистка организма, — общая и на клеточном уровне, стимулируется работа всех органов и систем, нормализуется обмен веществ, очищается и омолаживается кожа.
Под воздействием процедуры «Кедровая бочка» происходит:
- Избавление от синдрома «хронической усталости» в результате нормализации обменных процессов, улучшения работы всех органов и систем.
- Избавление от лишнего веса. Действует она как мини-сауна, Тело хорошо разогревается, улучшается кровоток, активизируются обменные процессы. Замедляется накопление и отложение жиров. Организм начинает постепенно потреблять накопленные запасы.
- Снятие стрессов. Успокоение и умиротворение наступает буквально с первого раза. Приятный насыщенный лечебными травами пар окутывает тело, конденсируется на коже, капельки стекают вниз, смывая пот и снимая стресс. Напряженные мышцы при этом приятно расслабляются. Вырабатывается устойчивость к стрессам.
- Профилактика сердечно-сосудистых заболеваний. Воздействие «Кедровой бочки» на сердечно-сосудистую систему проявляется в расширении и увеличении количества функционирующих капилляров, облегчении продвижения крови по артериям.
- Омолаживание кожи. Практически все функции кожи в «Кедровой бочке» активизируются. Потовые железы выделяют большое количество секрета, происходит шелушение эпидермиса, число патогенных микроорганизмов на коже уменьшается. При регулярном пользовании «Кедровой бочкой» проявляется выраженный омолаживающий косметический эффект, кожа становится упругой, гладкой, эластичной.
- «Кедровая бочка» восстанавливает нарушенный лимфоотток, улучшает лимфоциркуляцию. Это даёт выраженный антицеллюлитный эффект.
- «Кедровая бочка» хорошо помогает при мышечном переутомлении. Насыщенный целебными травами пар оказывает воздействие на окислительно-восстановительные процессы в мышцах. Приток крови насыщенной кислородом активизирует работу клеток мышечной ткани. Поэтому в «Кедровой бочке» снимается мышечная усталость, восстанавливается работоспособность мышц.
- Через очищенные раскрывшиеся поры в организм проникают натуральные лечебные вытяжки из трав и кедра, увеличивая количество различных необходимых человеку микроэлементов в крови. Вследствие этого нормализуется деятельность коры надпочечников, щитовидной железы, половых желез. «Кедровая бочка» способствует и увеличению потенции.
- Курс процедур в «Кедровой бочке» активно влияет на обмен веществ, усиливая минеральный, газовый и белковый обмен, способствует выведению из организма азотистых веществ, неорганического фосфора, натрия хлорида, мочевины, мочевой кислоты.
 Напряженные мышцы при этом приятно расслабляются. Вырабатывается устойчивость к стрессам.
Напряженные мышцы при этом приятно расслабляются. Вырабатывается устойчивость к стрессам. Приток крови насыщенной кислородом активизирует работу клеток мышечной ткани. Поэтому в «Кедровой бочке» снимается мышечная усталость, восстанавливается работоспособность мышц.
Приток крови насыщенной кислородом активизирует работу клеток мышечной ткани. Поэтому в «Кедровой бочке» снимается мышечная усталость, восстанавливается работоспособность мышц.
Противопоказания:
- Острые воспалительные заболевания.
- Хронические процессы в стадии обострения.
- Кровотечения.
- Злокачественные новообразования.
- Доброкачественные образования с тенденциями к росту.
- Активный туберкулез.
- Недостаточность кровообращения выше 1 степени.
- Тромбофлебит глубоких вен. Мерцательная аритмия.
- Сердечно-сосудистые заболевания 2-ой степени.
- Атеросклероз, протекающий с ярко выраженными поражениями кровеносных сосудов сердца и головного мозга.
- Гипертоническая болезнь 3-й степени.
- Вторая половина беременности.
- Мокнущая экзема.
- Цирроз печени.

Что такое витамины? – Коммерсантъ Красноярск
Витамины – это особые вещества, необходимые организму для полноценного функционирования. Они не обладают пищевой ценностью, но выполняют важнейшие функции – регулируют обменные процессы, катализируют биохимические реакции, помогают в усвоении многих минеральных компонентов. При дефиците витаминов развиваются различные обменные нарушения, которые приводят к сбоям в работе организма и формированию патологий. Дефицит возникает при нарушении их поступления в организм или проблемах с их всасыванием и усвоением.
Дефицит возникает при нарушении их поступления в организм или проблемах с их всасыванием и усвоением.
Существует много классификаций витаминов, однако самым основным является разделение их на растворимые в воде и растворимые в жирах. Эта особенность витаминов во многом определяет их метаболические превращения в организме и пути выведения. Кроме того, витамины, растворимые в воде, практически не накапливаются в теле, за исключением В12, который копится в тканях печени. В виду хорошей растворимости в воде, эти активно участвуют в метаболизме и выводятся с мочой, передозировка их практически не возникает, даже если они принимаются в несколько повышенных дозировках.
Классификация витаминов к водорастворимым относит группы В, аскорбиновую кислоту, Р, биотин (Н).
С жирорастворимыми витаминами все сложнее. Эти вещества не растворимы в воде, они растворяются в жирах, что позволяет им накапливаться в организме, создавая определенное депо. Однако, это же их свойство опасно тем, что прием повышенных доз (что вполне вероятно при неправильном, самостоятельном рассечете) может грозить передозировкой.
К группе жирорастворимых витаминов относят ретинол (А), К, токоферол (Е) и Д. Также зачастую к этой группе причисляют F – это целая группа особых ненасыщенных жирных кислот, необходимых для полноценной работы сердца и сосудов, красоты кожи и нормального самочувствия.
Среди всех витаминов этой группы, Д может частично синтезироваться в организме, а К практически полностью синтезируется за счет здоровой микробной флоры кишечника. Остальные должны регулярно поступать в организм, чтобы создавать запасы и расходоваться на нужды тела. Жирорастворимые витамины крайне важны для здоровья, они участвуют во многих жизненно важных процессах, обмене минералов, свертывании крови и функционировании органов чувств. Однако, данные витамины способны полноценно усваиваться в условиях присутствия жиров, в которых они растворяются и совместно с которыми всасываются и работают.
Обычно они содержатся в животных продуктах, где кроме них самих, имеются и жиры, помогающие усвоению. Если это растительные продукты, усвоение жирорастворимых витаминов из них будет активным при наличии жира (растительные масла, сливочное масло, сметана, сливки).
Естественно, что основным источником витаминов, как водо- так и жирорастворимых является пища. Источниками ретинола будут различные плоды, имеющие желтую или оранжевую окраску, масла (как растительные, так сливочное), а также животные продукты – яйца, молоко, мясо, печень, рыбий жир.
• Д способен синтезироваться в коже под действием ультрафиолета, а также поступает с пищей – богаты им икра, жирная рыба, мясо, желтки, печень и сливочное масло.
• Е содержится в растительных маслах, орехах, пророщенных злаках, семечках, зелени, жирных сливках, маргаринах.
• К производят полезные микробы кишечника, кроме того, он содержится в зеленых и белых овощах (капуста, шпинат, листовой салат, огородная зелень).
• F можно найти в растительных маслах первого отжима, морепродуктах, овсяной крупе, кукурузе, авокадо и миндале.
«Трансформация – это естественная реакция на изменения внешних факторов»
На сегодняшний день основная задача STEP LOGIC – максимально безболезненно провести изменение каналов коммуникации с заказчиками и настроить гибкое взаимодействие внутри рабочих команд. Удаленная и распределенная работа требует большей автономности, поэтому продолжаются процессы максимального внедрения принятия решений на основе data-driven-подхода. Об этом рассказывает заместитель генерального директора по техническому развитию STEP LOGIC Роман Остапчук.
Удаленная и распределенная работа требует большей автономности, поэтому продолжаются процессы максимального внедрения принятия решений на основе data-driven-подхода. Об этом рассказывает заместитель генерального директора по техническому развитию STEP LOGIC Роман Остапчук.
– Роман, какие изменения, происходящие внутри компаний вашего сектора, сейчас наиболее актуальны?
– Практически все проекты по внедрению ИТ-технологий — сложные цепочки взаимодействия разных организаций. Ранее в рабочих процессах мы активно использовали прямые коммуникации между людьми. Текущая ситуация заставляет нас перестраивать взаимодействие по направлениям, связанным с координацией и выполнением работ, прогнозированием и планированием, обучением.
В то же время пандемия дала необходимый толчок для внедрения более эффективных методов, не требующих непосредственного физического участия, а текущие ограничения позволяют активнее тиражировать такие практики.
– Какие изменения в планы по внутреннему развитию внесла именно пандемия?
– Пандемия внесла значительные коррективы в наши рабочие процессы. Компания ускорила внутренние проекты по автоматизации процессов, автономному принятию решений на основе объективных данных.
Серьезное влияние переход на удаленный режим работы оказал на структуру работы с персоналом, на процессы обучения и развития технологического портфеля.
Вопросы технической поддержки, продаж новых комплексных решений, равно как и качественная реализация проектов, невозможны без слаженной и оперативной работы множества сотрудников. Для поддержки таких команд мы создали омниканальную рабочую ИТ-платформу, интегрирующую все потоки входящих и исходящих данных и рабочие инструменты: WFS, BMP, почту, телефон и Instant Messaging.
– Что еще побуждает ИТ-компании меняться сейчас?
– ИТ-компании вынуждены адаптироваться к серьезным изменениям внешних факторов, таким как усиление конкуренции, диверсификация отраслей экономики, серьёзный тренд на импортозамещение, кадровый голод. Все это вынуждает игроков рынка искать и проверять новые гипотезы, модифицировать существующие рабочие практики, иногда возвращать в новом виде рабочие методы, от которых осознанно отказались ранее.
Все это вынуждает игроков рынка искать и проверять новые гипотезы, модифицировать существующие рабочие практики, иногда возвращать в новом виде рабочие методы, от которых осознанно отказались ранее.
– Какие нововведения, преобразования становятся сейчас условием сохранения конкурентоспособности и почему? Можно ли, вообще, оставаться конкурентоспособным, не трансформируясь?
– Трансформация — это естественная реакция на изменения внешних факторов. Если компания имеет уникальную бизнес-модель и положение на рынке, то она может оставаться конкурентоспособной достаточно длинный промежуток времени, не обращая внимание на изменения внешней среды. Однако подавляющее большинство ИТ-компаний находятся в условиях погружения в обменные процессы экономики и социума, требующих непрерывной адаптации.
В настоящий момент ключевым фактором давления в ИТ-секторе является непростая экономическая ситуация. Остро, на мой взгляд, стоит также кадровый вопрос.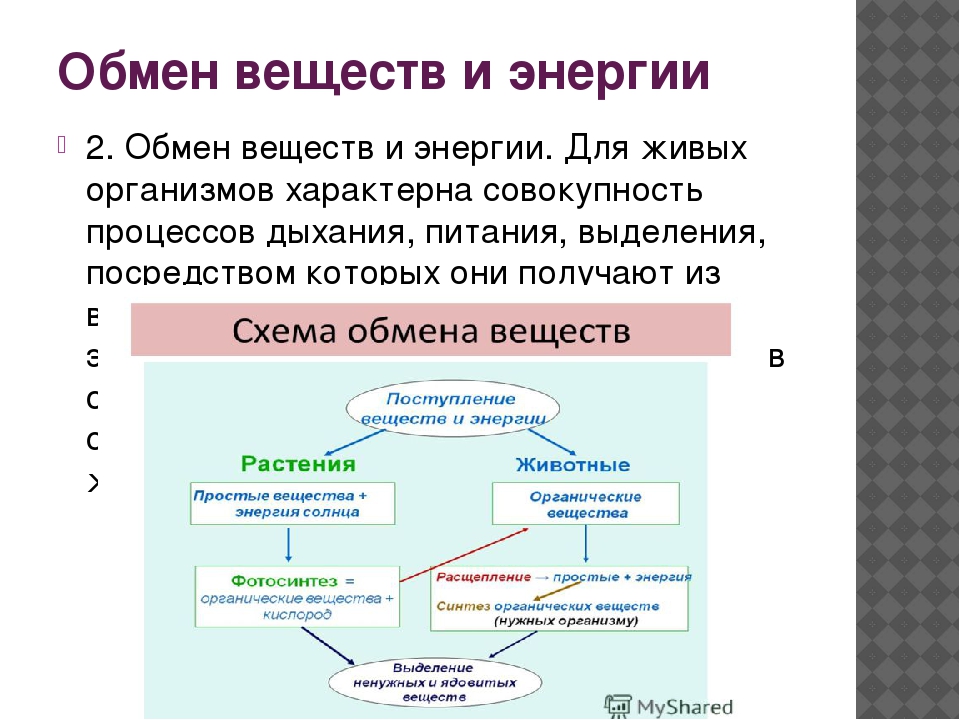 По подсчетам аналитиков, к 2024 году года дефицит квалифицированных ИТ-специалистов составит 300 тыс. человек в год.
По подсчетам аналитиков, к 2024 году года дефицит квалифицированных ИТ-специалистов составит 300 тыс. человек в год.
Ввиду вышеперечисленных проблем, наиболее актуальными для отрасли являются вопросы точного планирования, технико-экономического обоснования, управления рисками, а также эффективного и гибкого управления кадрами.
– А есть ли нечто, что можно отнести к «модным», имиджевым трендам?
– Надеюсь, что в это сложное время наиболее «модным» и «имиджевым» трендом для ИТ-компаний станет всесторонняя социальная поддержка сотрудников, помощь в адаптации к текущим рабочим условиям, обучение новым технологиям. Со своей стороны, STEP LOGIC стремится следовать наилучшим практикам в этой области, создавая комфортную среду для работы своих сотрудников и заказчиков.
– Какие изменения происходят внутри вашей компании?
– На сегодняшний день наша основная задача — максимально безболезненно провести изменение каналов коммуникации с нашими заказчиками и настроить гибкое взаимодействие внутри рабочих команд.
Удаленная и распределенная работа требует большей автономности, поэтому продолжаются процессы максимального внедрения принятия решений на основе data-driven-подхода.
– Расскажите подробнее о нескольких внутренних проектах.
– Удаленный характер совместной работы не позволяет больше делать ставку на личный обмен информацией в локальных командах. Сейчас мы дорабатываем существующие ИТ-инструменты компании для оперативного, удобного, конфиденциального и безопасного сбора всех критичных данных. Следующий шаг – увеличение доли автономно прорабатываемых задач и принимаемых решений.
Ключевым внутренним проектом этого периода является реинжиниринг сбора, анализа данных, принятия решений и автоматизирующих их информационных систем, который позволит нам поддерживать существующий высокий SLA исполнения рабочих задач.
– Меняет ли ваша компания в связи с проведением этих проектов свое позиционирование, и если да, то как?
– На мой взгляд, основная задача системной интеграции — концентрация разнообразного практического опыта и его передача другим организациям.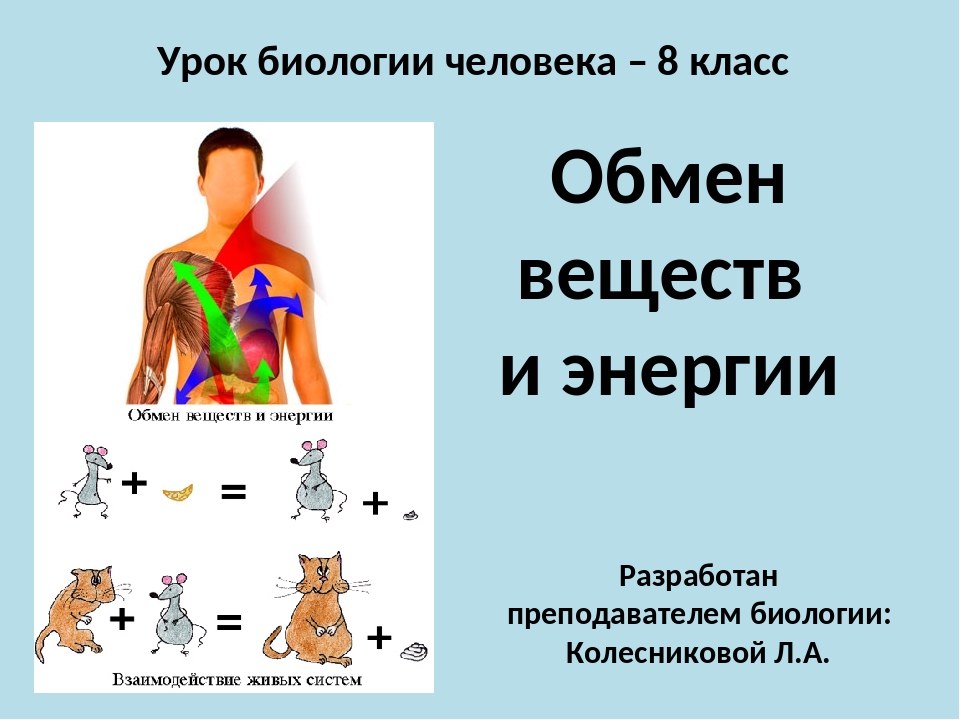 В наших планах дополнить существующее позиционирование STEP LOGIC как универсального ИТ-интегратора экспертизой в отраслевой специфике и инновационных решениях для конкретных сфер деятельности.
В наших планах дополнить существующее позиционирование STEP LOGIC как универсального ИТ-интегратора экспертизой в отраслевой специфике и инновационных решениях для конкретных сфер деятельности.
– Ваш прогноз насчет дальнейших преобразований в ИТ-компаниях отечественного рынка системной интеграции — что потребуется трансформировать в ближайшей перспективе, почему, для достижения каких целей?
– Сегодня системные интеграторы все чаще выступают в роли проводников перспективных решений, вырабатывая оптимальные ИТ-инструменты для эффективных инвестиций как в развитие новых экспертиз, так и в operational excellence для существующих направлений бизнеса. Системная интеграция делает большой вклад в развитие технологий. Его значимость не снизится даже в условиях увеличения количества производителей и качественном росте собственной экспертизы конечных заказчиков.
– Большое спасибо за беседу!
Источник: ict-online.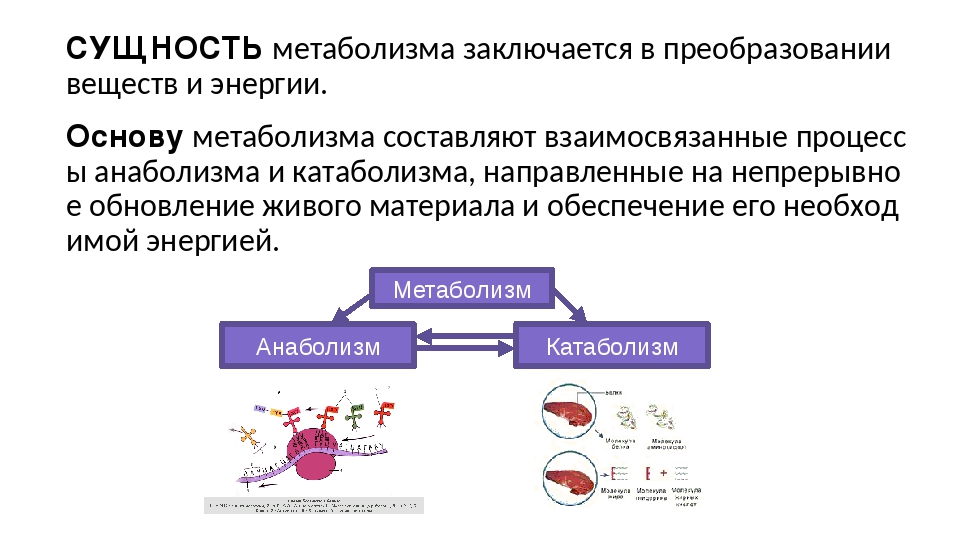 ru
ru
Самый быстрый словарь в мире: Vocabulary.com
метаболический процесс органические процессы, необходимые для жизни
адиабатический процесс любой процесс, который происходит без увеличения или уменьшения тепла
математический процесс (математика) расчет математическими методами
метаболический алкалоз, алкалоз, возникший в результате потери ионов водорода или чрезмерного потребления щелочных веществ
 85″>
85″>патологический процесс органический процесс, возникший вследствие болезни
нарушение обмена веществ нарушение или дефект обмена веществ
скорость метаболизма скорость обмена веществ
геологический процесс естественный процесс, в результате которого изменяются геологические особенности
психический процесс выполнение некоторой сложной познавательной деятельности
патологический процесс органический процесс, возникший вследствие болезни
биологический процесс процесс, происходящий в живых организмах
Шиловидный отросток идет от основания височной кости
телесный процесс органический процесс, происходящий в теле
крыловидный отросток два костных отростка, отходящих от тела клиновидной кости
естественный процесс Процесс, существующий в природе или производимый ею
геологический процесс естественный процесс, в результате которого изменяются геологические особенности
метаболизм необходимые для жизни органические процессы
фотожелатиновый процесс фотомеханический процесс печати, в котором используется стеклянная пластина с желатиновой поверхностью, на которой размещается воспроизводимое изображение; можно использовать с одним или несколькими цветами
Марковский процесс простой случайный процесс, в котором распределение будущих состояний зависит только от текущего состояния, а не от того, как оно перешло в текущее состояние
Химические реакции в метаболических процессах
Химические реакции в метаболических процессах
Чтобы произошла химическая реакция, реагирующие молекулы (или атомы) должны сначала столкнуться, а затем иметь достаточную энергию (энергию активации), чтобы вызвать образование новых связей.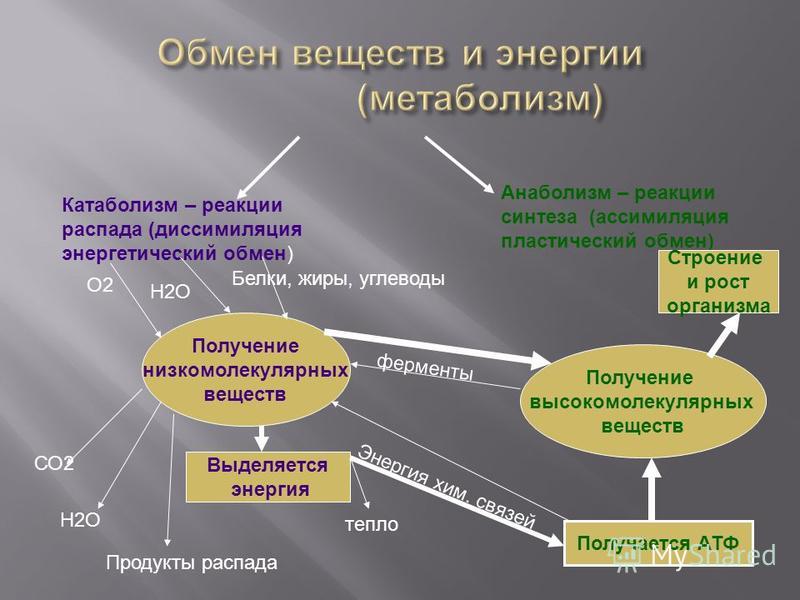 Хотя многие реакции могут происходить самопроизвольно, присутствие катализатора ускоряет скорость реакции, поскольку снижает энергию активации, необходимую для протекания реакции. Катализатор — это любое вещество, которое ускоряет реакцию, но само не претерпевает химических изменений. Поскольку катализатор не изменяется в результате реакции, его можно использовать снова и снова.
Хотя многие реакции могут происходить самопроизвольно, присутствие катализатора ускоряет скорость реакции, поскольку снижает энергию активации, необходимую для протекания реакции. Катализатор — это любое вещество, которое ускоряет реакцию, но само не претерпевает химических изменений. Поскольку катализатор не изменяется в результате реакции, его можно использовать снова и снова.
Химические реакции, происходящие в биологических системах, называются метаболизмом. Метаболизм включает расщепление веществ (катаболизм), образование новых продуктов (синтез или анаболизм) или передачу энергии от одного вещества к другому. Метаболические процессы имеют следующие общие характеристики:
- Ферменты действуют как катализаторы метаболических реакций. Ферменты — это белки, специфичные для определенных реакций. Стандартный суффикс для ферментов — «аза», поэтому легко определить ферменты, в которых используется это окончание (хотя у некоторых нет). Вещество, на которое действует фермент, называется субстратом. Например, фермент амилаза катализирует расщепление субстрата амилозы (крахмала) с образованием глюкозы. Модель с индуцированной подгонкой описывает, как работают ферменты. Внутри белка (фермента) есть активный сайт, с которым реагенты легко взаимодействуют из-за формы, полярности или других характеристик активного сайта. Взаимодействие реагентов (субстрата) и фермента заставляет фермент изменять форму.Новое положение помещает молекулы субстрата в положение, благоприятное для их реакции, и ускоряет образование продукта.
 Вещество, на которое действует фермент, называется субстратом. Например, фермент амилаза катализирует расщепление субстрата амилозы (крахмала) с образованием глюкозы. Модель с индуцированной подгонкой описывает, как работают ферменты. Внутри белка (фермента) есть активный сайт, с которым реагенты легко взаимодействуют из-за формы, полярности или других характеристик активного сайта. Взаимодействие реагентов (субстрата) и фермента заставляет фермент изменять форму.Новое положение помещает молекулы субстрата в положение, благоприятное для их реакции, и ускоряет образование продукта.
Вещество, на которое действует фермент, называется субстратом. Например, фермент амилаза катализирует расщепление субстрата амилозы (крахмала) с образованием глюкозы. Модель с индуцированной подгонкой описывает, как работают ферменты. Внутри белка (фермента) есть активный сайт, с которым реагенты легко взаимодействуют из-за формы, полярности или других характеристик активного сайта. Взаимодействие реагентов (субстрата) и фермента заставляет фермент изменять форму.Новое положение помещает молекулы субстрата в положение, благоприятное для их реакции, и ускоряет образование продукта.- Аденозинтрифосфат (АТФ) является обычным источником энергии активации метаболических реакций. На рисунке 1 волнистые линии между двумя последними фосфатными группами молекулы АТФ указывают на высокоэнергетические связи. Когда АТФ поставляет энергию в реакцию, обычно в реакцию доставляется энергия последней связи.В процессе передачи этой энергии последняя фосфатная связь разрывается, и молекула АТФ превращается в АДФ (аденозиндифосфат) и фосфатную группу (обозначенную P i ). Напротив, новые молекулы АТФ собираются путем фосфорилирования, когда АДФ объединяется с фосфатной группой с использованием энергии, полученной от какой-либо богатой энергией молекулы (например, глюкозы).
- Кофакторы — это небелковые молекулы, которые помогают ферментам. Холофермент — это союз кофактора и фермента (называемый апоферментом, когда он входит в состав холофермента).Если кофакторы являются органическими, они называются коферментами и обычно действуют, чтобы отдавать или принимать какой-либо компонент реакции, часто электроны. Некоторые витамины являются коферментами или компонентами коферментов. Неорганические кофакторы часто представляют собой ионы металлов, например Fe ++ .
 Напротив, новые молекулы АТФ собираются путем фосфорилирования, когда АДФ объединяется с фосфатной группой с использованием энергии, полученной от какой-либо богатой энергией молекулы (например, глюкозы).
Напротив, новые молекулы АТФ собираются путем фосфорилирования, когда АДФ объединяется с фосфатной группой с использованием энергии, полученной от какой-либо богатой энергией молекулы (например, глюкозы).Рис. 1. Высокоэнергетические связи аденозинтрифосфата (АТФ).
Синхронизация обменных процессов в растениях с метаболизмом крассулоидной кислоты | Журнал экспериментальной ботаники
Аннотация
У растений с метаболизмом крассулоидной кислоты разделение процессов карбоксилирования, опосредованных фосфо енол пируваткарбоксилазой (PEPC) и Rubisco, оптимизирует фотосинтетические характеристики и увеличение количества углерода в потенциально ограничивающей среде.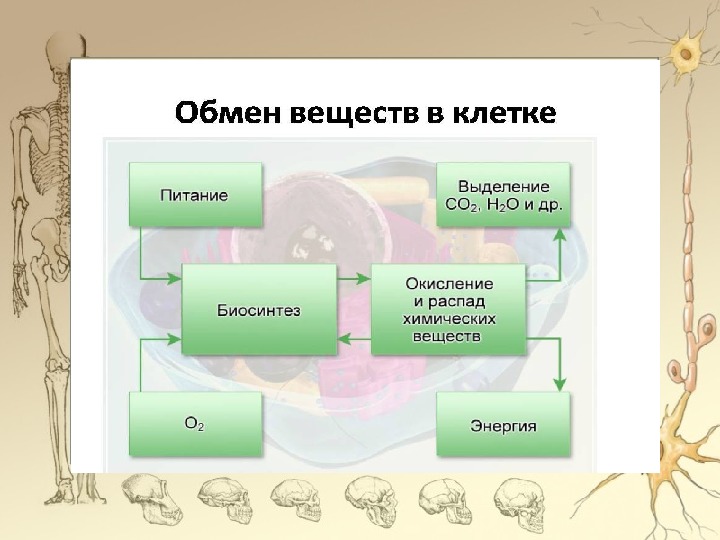 В этом обзоре рассматриваются механизмы, которые синхронизируют спрос и предложение на углерод при сохранении фотосинтетической пластичности в течение 24-часового цикла САМ. Циркадные часы играют центральную роль в управлении многими метаболическими, транспортными и физиологическими компонентами САМ. Уровень контроля, осуществляемого часами, может варьироваться от транскрипционной до посттрансляционной регуляции, в зависимости от рассматриваемых генов, белков и даже видов растений. Дополнительный уровень контроля обеспечивается метаболитами, в том числе органическими кислотами и углеводами, содержание которых в течение цикла диэфира существенно меняется.Обсуждаются механизмы, ответственные за определение содержания метаболитов, а также требования к передаче сигналов для координации потоков углерода. Эволюционные последствия рассматриваются с точки зрения того, как циркадный и метаболический контроль цикла САМ мог быть получен из растений C 3 .
В этом обзоре рассматриваются механизмы, которые синхронизируют спрос и предложение на углерод при сохранении фотосинтетической пластичности в течение 24-часового цикла САМ. Циркадные часы играют центральную роль в управлении многими метаболическими, транспортными и физиологическими компонентами САМ. Уровень контроля, осуществляемого часами, может варьироваться от транскрипционной до посттрансляционной регуляции, в зависимости от рассматриваемых генов, белков и даже видов растений. Дополнительный уровень контроля обеспечивается метаболитами, в том числе органическими кислотами и углеводами, содержание которых в течение цикла диэфира существенно меняется.Обсуждаются механизмы, ответственные за определение содержания метаболитов, а также требования к передаче сигналов для координации потоков углерода. Эволюционные последствия рассматриваются с точки зрения того, как циркадный и метаболический контроль цикла САМ мог быть получен из растений C 3 .
Поступило 28 октября 2003 г . ; Принята к печати 19 января 2004 г.
; Принята к печати 19 января 2004 г.
Введение
Метаболизм крассулоидной кислоты — это особый способ фотосинтетической ассимиляции углерода, который развился в ответ на исключительные условия окружающей среды.На сегодняшний день известно, что примерно 7% видов растений, состоящих из 33 семейств и 328 родов, обладают способностью к CAM (Winter and Smith, 1996, a ). Такое таксономическое разнообразие отражается в диапазоне сред обитания, предпочитаемых CAM-растениями, который варьируется от засушливых пустынь, тропических лесов до водных экосистем. CAM представляет собой концентрирующий механизм CO 2 , в котором используется фермент фосфо енол пируваткарбоксилаза (PEPC) для улавливания дыхательного и атмосферного CO 2 в ночное время.Физиологическое значение CAM заключается в сохранении углерода и воды в растениях, растущих в среде, которая ограничивает доступность одного или обоих ресурсов на периодической или долгосрочной основе. В то время как ферментный аппарат, необходимый для CAM, присутствует у всех высших растений, эволюция пути потребовала изменения регуляторной способности ключевых ферментов и переносчиков, чтобы поддерживать временное разделение процессов карбоксилирования, которые являются центральными для CAM. Цель этого обзора — рассмотреть механизмы, которые могут синхронизировать спрос и предложение на углерод в течение 24-часового цикла CAM.Молекулярные подходы и новые геномные ресурсы предоставляют беспрецедентный потенциал для использования цикла DIEL CAM для выяснения компонентов контроля циркадных ритмов и метаболитов, которые оптимизируют фотосинтетические характеристики в потенциально ограниченных и экстремальных условиях.
В то время как ферментный аппарат, необходимый для CAM, присутствует у всех высших растений, эволюция пути потребовала изменения регуляторной способности ключевых ферментов и переносчиков, чтобы поддерживать временное разделение процессов карбоксилирования, которые являются центральными для CAM. Цель этого обзора — рассмотреть механизмы, которые могут синхронизировать спрос и предложение на углерод в течение 24-часового цикла CAM.Молекулярные подходы и новые геномные ресурсы предоставляют беспрецедентный потенциал для использования цикла DIEL CAM для выяснения компонентов контроля циркадных ритмов и метаболитов, которые оптимизируют фотосинтетические характеристики в потенциально ограниченных и экстремальных условиях.
По сути, CAM может экспрессироваться на фоне фотосинтеза C 3 посредством развертывания ночного карбоксилирования и последующих процессов декарбоксилирования в дневное время (Fig. 1a). Ночью, когда скорость эвапотранспирации низкая, атмосферный CO 2 и / или респираторный CO 2 фиксируются в цитозоле ферментом фосфо енол пируваткарбоксилазой (PEPC). Субстрат 3-C, фосфо енол пируват (PEP), образуется в результате гликолитического расщепления углеводов, образовавшихся в течение предыдущего дня. Конечный продукт 4-C, яблочная кислота, хранится в большой центральной вакуоли. В течение дня малат выходит из вакуоли, и декарбоксилирование может происходить за счет однократного или комбинированного действия трех карбоксилаз (в зависимости от вида растений): НАДФ-яблочный фермент (НАДФ-МЭ), НАД-МЕ и фосфо- енол пируваткарбоксикиназа ( PEPCK). Помимо продуктов 3-C PEP или пирувата, CO 2 выделяется при высоком внутреннем парциальном давлении ( p CO 2 ), которого часто бывает достаточно для закрытия устьиц и, таким образом, экономии воды.Высокое значение p CO 2 , генерируемое декарбоксилированием, также подавляет фотодыхание. Восстановление углеводов посредством глюконеогенеза требует высоких энергетических затрат на этот путь, но обеспечивает производство субстрата для последующего ночного карбоксилирования и разделения для роста. Углеводы, которые обеспечивают субстрат для ночных реакций, транспортируются либо в хлоропласт и хранятся в виде крахмала, либо транспортируются в вакуоль и хранятся в виде сахарозы и / или гексозы, в зависимости от вида (Christopher and Holtum, 1996).Что касается чистого потока углерода, результатом реакций, показанных на рис. 1а, являются существенные низкие и возвратно-поступательные колебания содержания малата и углеводов (рис. 1b), которые могут составлять до 20% от сухой массы листа.
Субстрат 3-C, фосфо енол пируват (PEP), образуется в результате гликолитического расщепления углеводов, образовавшихся в течение предыдущего дня. Конечный продукт 4-C, яблочная кислота, хранится в большой центральной вакуоли. В течение дня малат выходит из вакуоли, и декарбоксилирование может происходить за счет однократного или комбинированного действия трех карбоксилаз (в зависимости от вида растений): НАДФ-яблочный фермент (НАДФ-МЭ), НАД-МЕ и фосфо- енол пируваткарбоксикиназа ( PEPCK). Помимо продуктов 3-C PEP или пирувата, CO 2 выделяется при высоком внутреннем парциальном давлении ( p CO 2 ), которого часто бывает достаточно для закрытия устьиц и, таким образом, экономии воды.Высокое значение p CO 2 , генерируемое декарбоксилированием, также подавляет фотодыхание. Восстановление углеводов посредством глюконеогенеза требует высоких энергетических затрат на этот путь, но обеспечивает производство субстрата для последующего ночного карбоксилирования и разделения для роста. Углеводы, которые обеспечивают субстрат для ночных реакций, транспортируются либо в хлоропласт и хранятся в виде крахмала, либо транспортируются в вакуоль и хранятся в виде сахарозы и / или гексозы, в зависимости от вида (Christopher and Holtum, 1996).Что касается чистого потока углерода, результатом реакций, показанных на рис. 1а, являются существенные низкие и возвратно-поступательные колебания содержания малата и углеводов (рис. 1b), которые могут составлять до 20% от сухой массы листа.
Несмотря на энергетические затраты, связанные с CAM, во многих случаях потенциал высокой производительности не снижается. Агрономически важные виды САМ, включая ананас ( Ananas comosus ) и некоторые виды агав, могут показывать продуктивность, сопоставимую с продуктивностью сахарного тростника (Bartholomew and Kadzimin, 1977; Nobel, 1996).Такие желательные атрибуты являются следствием пластичности, с помощью которой CAM может быть задействован или отключен в ответ на периодические или долгосрочные (сезонные) возмущения окружающей среды. Факторы развития и окружающей среды (например, доступность воды, интенсивность света) сильно влияют на долю CO 2 , поглощаемую ночью через PEPC и непосредственно в течение дня через Rubisco на заводах CAM (Cushman and Borland, 2002; Dodd et al . , 2002). Рост и продуктивность большинства CAM-растений максимальны, когда преобладает прямая дневная фиксация CO 2 через Rubisco (фаза IV газообмена).Как определяющая черта САМ, фотосинтетическая пластичность должна быть достигнута при одновременном поддержании синхронизации между процессами карбоксилирования, декарбоксилирования и транспорта, показанными на рис. 1а, чтобы минимизировать бесполезные циклы круговорота углерода в течение цикла диль.
Циркадные часы устанавливают основные фазы CAM
Хорошо задокументированные ритмы газообмена у видов рода Kalanchoë , выращенных в непрерывной темноте и без CO 2 в воздухе, показали, что эндогенные циркадные часы играют кардинальную роль в установлении и синхронизации разделенных во времени метаболических процессов. компоненты САМ (Wilkins, 1992; Lüttge, 2000).Циркадный контроль потока углерода через PEPC обычно рассматривается как ключевой компонент, лежащий в основе разделения процессов карбоксилирования день / ночь, которые определяют CAM (Nimmo, 2000). PEPC активируется ночью посредством фосфорилирования остатка серина рядом с N-концом белка, что делает фермент более чувствительным к PEP и положительным эффекторам, глюкозе-6-P и триозе-P, и менее чувствительным к аллостерическому ингибитору, малату. (Рис.2; Nimmo et al ., 1986, 1987; Chollet et al ., 1996). Временной интервал, в течение которого PEPC остается активным, на что указывают образцы ночного накопления малата, таким образом, отражается в явных изменениях состояния фосфорилирования фермента, на что указывает [малат], необходимый для 50% -ного ингибирования фермента (т. Е. K i малат). Более того, степень фосфорилирования PEPC является основным определяющим фактором количества CO 2 , поглощенного и сохраненного в виде малата в течение ночи у разных видов САМ, где экстрагируемая активность PEPC варьировалась не более чем на 10% (рис.3).
Состояние фосфорилирования PEPC определяется наличием или отсутствием выделенной Ca 2+ -независимой протеинкиназы Ser / Thr, которая, в свою очередь, регулируется на уровне экспрессии генов с помощью циркадного осциллятора (рис. 2; Картер и др. ., 1991; Hartwell и др. ., 1996, 1999; Тайби и др. ., 2000). Теоретически контроль активности киназы PEPC (PPCK) с помощью эндогенных часов должен позволять предвидеть фотопериод и обеспечивать быструю инактивацию PEPC в начале дня, тем самым избегая бесполезного цикла синтеза малата и декарбоксилирования.Однако экологическое преимущество такого циркадного контроля поглощения CO 2 менее очевидно, учитывая присущую САМ-растениям пластичность для модуляции потока углерода в ответ на изменение условий окружающей среды. Полевые измерения мгновенной дискриминации изотопов углерода у полуэпифитных душителей из рода Clusia показали, что PEPc может оставаться активным в течение 4-5 часов после рассвета (Borland et al ., 1993; Roberts et al . , 1997). На рисунке 3 показано, что время инактивации PEPC может варьироваться между разными видами CAM, даже если они выращиваются в идентичных условиях окружающей среды.
Метаболиты обеспечивают дополнительный уровень контроля над циклом Diel CAM
Экспериментальные наблюдения теперь показывают, что экспрессия и активность PPCK могут быть изменены метаболическим статусом листа. В листьях Kalanchoë daigremontiana , которые не могли накапливать малат из-за помещения в атмосферу с N 2 в течение части или всей ночи, после переноса наблюдались резкие сдвиги в амплитуде и продолжительности поглощения CO 2 PEPC. к окружающему воздуху (рис.4а; Borland и др. ., 1999). Стимуляция поглощения CO 2 через PEPC в ночное время и в начале дня объясняется изменениями в величине активности PPCK (рис. 4b), которая контролируется на уровне экспрессии генов (Hartwell et al ., 1999; Borland и др. ., 1999). Наблюдения согласуются с точкой зрения, что цитоплазматический малат (или родственный метаболит) вызывает ингибирование по обратной связи экспрессии гена PPCK. Таким образом, у растений, у которых накапливается малат, активность и экспрессия PPCK выше, чем в контроле, и происходит задержка подавления PPCK у растений с пониженным содержанием малата (рис.4; Borland и др. ., 1999). Следовательно, было высказано предположение, что циркадный контроль экспрессии гена PPCK в САМ является вторичным ответом на циркадные изменения в транспорте малата через тонопластную мембрану вакуоли (Nimmo, 2000). Независимые доказательства в пользу того, что мембрана тонопласта является «главным переключателем» для циркадной регуляции CAM, была получена на основе компьютерного моделирования ритмов CAM, основанного на осмотических соображениях оборота малата и механизма напряжения / релаксации тонопласта (Lüttge, 2000, 2002 a). ).Поскольку сдвиги в дневных / ночных изменениях содержания малата являются отличительной чертой пластичности CAM, контроль метаболитов фосфорилирования PEPC обеспечит эффективное средство точной настройки поглощения CO 2 в течение дневного / ночного цикла в соответствии с изменениями условий окружающей среды. . Такая интеграция циркадных сигналов и сигналов окружающей среды может обеспечить основу для синхронизации и пластичности метаболизма, присущих САМ. Однако такая гипотеза вызывает дополнительные вопросы с точки зрения того, сколько метаболических компонентов цикла САМ напрямую связано с циркадным осциллятором и сколько зависит от дневных / ночных потоков метаболитов через тонопласт?
Какие компоненты CAM регулируются часами?
Гены, контролируемые часами
Анализы примерно 40 генов в индуцибельном CAM-растении, Mesembryanthemum crisinum , показали ритмические изменения в содержании транскриптов более 30 выбранных генов с пиками экспрессии на различных фазах в течение всего цикла diel (Boxall et al ., 2001, 2002). Более того, большее количество генов обнаруживает ритмические изменения в изобилии транскриптов у растений, экспрессирующих CAM, по сравнению с растениями в состоянии C 3 (Boxall et al ., 2001, 2002). Одним из способов, которым часы могут синхронизировать метаболические компоненты САМ, было бы фазирование транскрипции определенных генов в определенное время в дневном / ночном цикле, обеспечивая, таким образом, наибольшее количество соответствующих ферментов и транспортеров, когда это необходимо. Обнаружено, что гены управляются часами в CAM-выполнении M.crystalinum кодируют ферменты, участвующие в фотосинтезе, гликолизе, ночном поглощении CO 2 , декарбоксилировании, метаболизме сахарозы и крахмала (Dodd et al ., 2003), транспорте метаболитов хлоропластов и вакуолярной АТФазе (Boxall et al ., 2001, 2002). Таким образом, основные метаболические компоненты САМ, как показано на рис. 1а, по-видимому, подчиняются элементу контроля, осуществляемому циркадными часами. Текущий анализ генных чипов M. crystalinum , содержащих 8400 генов, несомненно, даст захватывающее представление о том, какие гены попадают под контроль часов при индуцировании САМ (Дж. Кушман, личное сообщение).
Хотя есть соблазн предположить, что ключевым событием в эволюции CAM было соединение большего количества / разных генов с центральным осциллятором, требуется осторожность при интерпретации функционального значения ритмических изменений в изобилии транскриптов. В растении C 3 , Arabidopsis thaliana , эксперименты на микроматрицах показали, что по крайней мере 6% генов экспрессируются ритмично (Harmer et al ., 2000; Schaffer et al ., 2001). Однако более поздние подходы, использующие захват in vivo энхансера, показали, что 36% генома Arabidopsis потенциально находится под транскрипционным контролем циркадных часов (Michael and McClung, 2003).Расхождение между этими отчетами может быть связано с различиями в стабильности транскриптов между генами. Таким образом, чтобы колебания транскрипции приводили к колебаниям в содержании мРНК, необходимо, чтобы транскрипт был достаточно нестабильным, чтобы переключаться в пределах циркадного цикла. Индукция CAM в M. crystalinum сопровождается увеличением стабильности транскриптов CAM-специфической изоформы PEPC, но снижением стабильности транскриптов, кодирующих небольшую субъединицу Rubisco (Cushman et al ., 1990; ДеРоше и Бонерт, 1993). Таким образом, регулирование сложного метаболического пути, такого как CAM, с помощью циркадных часов, вероятно, включает несколько уровней контроля, от транскрипции до посттрансляционной модификации белка, воздействуя на любое количество ферментов или транспортеров.
Пост-транскрипционный контроль по часам
Чтобы определить физиологическое значение циркадных колебаний в изобилии транскриптов, необходимо также оценить уровни количества / активности соответствующих ферментов.Например, хотя сообщается, что активность PPCK регулируется на уровне экспрессии гена (Hartwell et al. ., 1999; Taybi et al. ., 2000), обилие транскриптов CAM-специфической изоформы PEPC также показывает суточные значения. и циркадные колебания в M. crystalinum (пик к концу фотопериода), хотя белок PEPC и экстрагируемая активность остаются относительно постоянными (J Hartwell, личное сообщение; Boxall et al ., 2001). Повышенное количество дневных транскриптов PEPC также было отмечено у CAM-выполняющих видов Clusia , но не у видов C 3 C.multiflora (Т Тайби, неопубликованные наблюдения). У всех этих видов CAM одна важная роль часов может заключаться в контроле оборота белка PEPC и обновлении уровня фермента каждую ночь. Дальнейший уровень контроля циркадных ритмов обеспечивается ритмическими сдвигами в экспрессии и активности PPCK, которые, в свою очередь, активируют белок PEPC ночью. По мере того, как становится доступным больше данных о ритмической экспрессии генов из сравнений микрочипов C 3 и CAM-выполняющих M.crystalinum , будет важно связать такие исследования с обычными биохимическими измерениями количества и активности соответствующих ферментов и переносчиков. Такая информация поможет определить, требует ли экспрессия CAM не только изменения комплемента генов, связанных с часами, но также требует изменения уровня контроля (транскрипционного или посттранскрипционного), который осуществляется Часы.
Физиологические процессы, контролируемые часами
Дополнительный подход к решению вопроса о том, какие компоненты пути САМ регулируются часами, заключался в использовании явных циркадных ритмов газообмена в листьях, обнаруженных у некоторых видов, для изучения более высоких уровней метаболической организации, которые подпадают под контроль циркадного осциллятора.Недавний анализ стехиометрии поглощения CO 2 и накопления малата в M. crystalinum и K. daigremontiana во время свободных ритмов газообмена показывает, что карбоксилирование C 3 вносит основной вклад в генерацию ритмических Поглощение CO 2 при непрерывном освещении (Dodd et al ., 2002, 2003; Wyka and Lüttge, 2003). При длительных периодах непрерывного света ослабление активности РЕРС оказывается более быстрым, чем ослабление явного ритма чистого поглощения CO 2 .Это указывает на то, что пока эндогенный контроль фосфорилирования PEPC не является устойчивым при постоянном освещении, другой и, возможно, отдельный осциллятор, связанный с C 3 , поддерживает ритмы в чистом поглощении CO 2 (Wyka and Lüttge, 2003). В настоящее время неясно, может ли эта ритмическая активность в карбоксилировании C 3 быть напрямую связана с циркадным контролем активации / активности Rubisco или с устьичной проводимостью. Известно, что оба эти аспекта метаболизма подлежат, по крайней мере, некоторому элементу циркадного контроля у C 3 spp (Liu et al ., 1996; Уэбб, 2003). Концепция, согласно которой общий контроль явных ритмов захвата CO 2 в CAM-растениях обеспечивается посредством колебаний устьичной апертуры, обсуждалась в другом месте (Lüttge, 2002, a ; Wyka and Lüttge, 2003). В рамках этой структуры циркадные осцилляторы, которые непосредственно контролируют PEPC и Rubisco, могут составлять другие компоненты сети, которые стабилизируют и координируют поглощение CO 2 в ответ на петли обратной связи метаболитов и сигналы окружающей среды (Dodd et al ., 2003). Такие соображения согласуются с идеей, что устойчивые ритмы газообмена листа не обязательно контролируются одним центральным часовым механизмом, который может работать на уровне транскрипции генов. Скорее, свободные колебания в чистом поглощении CO 2 , по-видимому, управляются сложными регуляторными сетями, интегрированными во времени и пространстве (Lüttge, 2000, 2002, a ; Rascher et al., ., 2001).
Контроль метаболитов CAM
Ряд генов обнаруживают ритмические изменения в содержании транскриптов только после того, как CAM индуцируется в M.crystalinum (Boxall et al ., 2001, 2002), и возможно, что многие из этих ритмов могут возникать как последствие цикла метаболитов, связанного с CAM. Проблема контроля метаболитов также актуальна для понимания того, как отдельные осцилляторы, как предложено выше, могут быть интегрированы в пространстве и времени для координации обмена CO 2 в различных условиях окружающей среды. Существенное взаимное изменение органических кислот и углеводов происходит в течение 24-часового цикла САМ (рис.1b), и известно, что обе категории метаболитов влияют на экспрессию разнообразного диапазона генов (Smeekens, 2000; Stitt et al ., 2002).
Обнаружение и сигнализация органических кислот
Недавние доказательства в поддержку индуцированного метаболитами цикла существующих генов C 3 предоставлены сравнениями экспрессии киназы PEPC (PPCK) в диапазоне видов Clusia , которые показывают заметные различия в способности к CAM. Способность к CAM в Clusia , по-видимому, определяется количеством белка PEPC (Borland et al ., 1998), который, в свою очередь, регулируется на уровне обилия транскриптов PEPC (Taybi et al ., 2004). Однако ген, кодирующий PPCK, белок, ответственный за активацию PEPC, экспрессируется на сопоставимых уровнях в листьях как C 3 , так и CAM-выполняющих видов Clusia , и дневные / ночные изменения экспрессии PPCK, по-видимому, являются следствие диэтиленового цикла органических кислот и растворимых сахаров (рис. 5; Taybi et al ., 2004). Более того, отдельные изменения в изобилии транскриптов PPCK, по-видимому, контролируются посредством понижающей регуляции гена в течение дня, а не повышающей регуляции в течение ночи (рис.5). Эти результаты контрастируют с результатами, полученными с M. crystalinum , в котором транскрипты PPCK присутствуют в небольшом количестве в листьях C 3 и где индукция CAM сопровождается повышающей регуляцией количества транскриптов PPCK в ночное время (Taybi et al. al ., 2000). Дневной отток малата (или некоторых других метаболитов) из вакуоли может быть первичным сигналом для подавления экспрессии PPCK во время фотопериода у видов Clusia после индукции CAM.Последующее декарбоксилирование органических кислот будет ключевым фактором, влияющим на увеличение экспрессии PPCK на следующую ночь. Интересно отметить, что видов Clusia могут накапливать высокие концентрации как яблочной, так и лимонной кислот, а фоновые концентрации обеих органических кислот могут быть относительно высокими у растений, осуществляющих фотосинтез C 3 (Borland et al ., 1996). Опять же, это контрастирует с ситуацией в M. crystalinum , где содержание как малата, так и цитрата низкое в режиме C 3 .Таким образом, в Clusia может показаться, что дневная мобилизация органических кислот является ключевым фактором, регулирующим как экспрессию CAM, так и PEPC-киназы (Borland et al ., 1996; Roberts et al ., 1997. ). Заметная фенотипическая пластичность растений, таких как C. minor для быстрого и обратимого переключения между фотосинтезом C 3 и CAM, может, таким образом, объясняться транспортными и / или ферментативными процессами, которые регулируют распределение метаболитов, особенно органических кислот, между вакуолями. и цитозоль.
Шаги транспорта, которые опосредуют приток и отток яблочной и лимонной кислот через мембрану тонопласта, будут центральным компонентом в восприятии органических кислот и передаче сигналов в CAM-растениях. Ночное накопление малата включает стехиометрический транспорт 2H + пермалата с транспортом H + , управляемым тонопластом H + ‐ATPase и, дополнительно, с помощью H + ‐PP i ase (Smith et al. ., 1996). Циркадные колебания количества транскриптов субъединицы С V-АТФазы были описаны у M.crystalinum (Rockel et al ., 1997), но неизвестно, подчиняется ли активность V-АТФазы циркадной регуляции. Хотя фактические переносчики органических кислот еще не идентифицированы на молекулярном уровне, кажется вероятным, что на тонопласте растений САМ существуют отдельные системы притока и оттока (Smith et al ., 1996). Приток малата через тонопласт, по-видимому, опосредуется зависимым от напряжения анион-селективным каналом (Hafke et al ., 2003). Этот канал реагирует на pH цитоплазмы способом, который согласуется с ночным поглощением малат-аниона и помогает минимизировать бесполезное возвращение малата обратно в вакуоль во время дневной фазы декисления. Карбоксилатный переносчик тонопласта, который, по-видимому, способен транспортировать как малат, так и цитрат, был недавно клонирован из A. thaliana (Emmerlich et al ., 2003). Более того, количество транскриптов этого переносчика можно регулировать в ответ на концентрацию малата (Emmerlich et al ., 2003). CAM-ортолог транспортера карбоксилата Arabidopsis может потенциально действовать как ключевой сенсор для передачи сигналов, опосредованной органическими кислотами, в течение цикла diel CAM, регулируя поток через тонопласт в ответ на концентрацию малата.
Обнаружение углеводов и сигнализация
Доступность углеводов является основным ограничивающим фактором для поглощения темного CO 2 CAM-растениями (Borland and Dodd, 2002). В листьях M. crystalinum , которые были обеднены углеводами на 50% в результате воздействия воздуха без CO 2 в течение 24 часов, последующее чистое темное поглощение CO 2 окружающим воздухом было снижено на 50% по сравнению с контролем. несмотря на заметное увеличение экспрессии PEPC-киназы (Dodd et al ., 2003). Доказательства ощущения дефицита углеводов (и, возможно, органических кислот) у растений, ограниченного в ночном поглощении CO 2 , также указываются сдвигами в последующем дневном чистом поглощении CO 2 и распределении углеводов, которые могут компенсировать предыдущий ночной дефицит в накоплении углерода (Roberts et al ., 1997; Borland and Griffiths, 1997; Dodd et al ., 2002). Таким образом, углеводный статус, по-видимому, является ключевым элементом в синхронизации метаболических потоков в течение цикла DIEL CAM в соответствии с изменениями в условиях окружающей среды.
Углеводный статус определяется взаимодействием циркадных ритмов и контроля метаболитов, которые регулируют обмен и субклеточное разделение на протяжении всего цикла. Наиболее изученные модели CAM (например, M. crystalinum , K. daigremontiana ) производят преимущественно крахмал в хлоропласте в качестве временного резерва углерода для поддержки ночного карбоксилирования и синтеза малата. У растений C 3 ранее предполагалось, что суточное накопление крахмала находится под циркадным контролем (Li et al ., 1992; Geiger и др. ., 1995). Для CAM-выполняющих M. crystalinum был предложен циркадный контроль над основным регулятором скорости стадии биосинтеза крахмала, катализируемым АДФ-глюкозопирофосфорилазой (Boxall et al ., 2001, 2002; J Hartwell, личное сообщение). Ключевая роль переносчиков хлоропластов в регуляции углеводного обмена в течение цикла диэлита указывается на открытие, что хлоропласты CAM-выполняющих листьев M. crystalinum уникальны тем, что содержат три класса транслокаторов фосфата (i.е. PEP, триоза-P и транслокаторы глюкозы-6-P), в дополнение к переносчику глюкозы в хлоропластах (Häusler et al ., 2000). Более того, динамическая диэль и циркадная регуляция количества транскриптов и активности этих транслокаторов хлоропластов очевидна, когда индуцируется CAM (Häusler et al. ., 2000; Boxall et al. ., 2001, 2002). Переносчики хлоропластов PEP, триоза-P, глюкозы-6-P и глюкозы могут играть ключевую роль в определении уровней углеводов и связанных с ними метаболитов и, следовательно, в модулировании транспортной активности таким образом, чтобы поддерживать обменные метаболические потоки во время цикла CAM (Häusler и др. ., 2000).
Ритмические изменения в содержании транскриптов нескольких генов, кодирующих ферменты, участвующие в деградации крахмала, также были зарегистрированы у M. crystalinum , причем пик содержания транскриптов приходился на конец (субъективного) фотопериода (Boxall et al ., 2001, 2002; Додд и др. ., 2003). Циркадный контроль деградации крахмала в M. crystalinum может обеспечить средства прогнозирования потребностей в субстрате для ночного карбоксилирования, обеспечивая сохранение адекватных запасов углеводов в течение фотопериода.Однако также вероятно, что существуют механизмы для «определения» доступности углеводов в течение ночи, поскольку переход от C 3 к CAM приводит к устойчивому «измерению» запасов углеводов, которые поддерживают доступность субстрата в течение темного периода ( Dodd et al ., 2003). В настоящее время неясно, определяется ли это очевидное восприятие уровней углеводов притяжением от PEPC к субстрату 3-C или прямым изменением скорости деградации углеводов.Однако недавние результаты, полученные с CAM-дефицитным мутантом M. crystalinum , показывают, что активность ферментов, расщепляющих крахмал, может модулироваться в соответствии с доступностью субстрата. В мутантной линии, выделенной группой Джона Кушмана, неспособность накопить малат в течение ночи, по-видимому, коррелирует с низким содержанием крахмала (Branco et al ., 2003). У этих CAM и крахмалодефицитных растений активность и флуктуации хлоропластной фосфорилазы крахмала также снижены по сравнению с диким типом (A. Borland и J Cushman, неопубликованные наблюдения).Точно так же мутант демонстрирует более низкую активность и пониженные отклонения диэлемента ряда глюкан-гидролаз, что указывает на то, что активность по разложению крахмала может регулироваться в соответствии с доступностью субстрата (A. Borland и J Cushman, неопубликованные наблюдения). Таким образом, деградация крахмала в M. crystalinum , по-видимому, подлежит циркадному контролю на уровне экспрессии генов и контролю метаболитов на уровне ферментативной активности.
Во многих растениях CAM, включая виды сельскохозяйственных культур Ananas comosus (ананас), Aloe vera и Agave , вакуолярные растворимые сахара являются преобладающей формой углеводов, накапливаемых в течение дня для поддержки цикла CAM (зима and Smith, 1996, b ).Дневные / ночные потоки сахаров через тонопластную мембрану ананаса могут быть значительными (до 20% сухой биомассы листа) и являются основным детерминантом экспрессии САМ. Предполагаемый вакуолярный переносчик сахарозы был описан в везикулах тонопласта A. comosus , который демонстрирует кинетические характеристики унипортера сахарозы и который, по-видимому, способен способствовать значительному притоку сахаров в вакуоль (McRae et al ., 2002). Исследования плазматической мембраны растений C 3 показали, что экспрессия транскриптов транспорта сахара может быть очень чувствительной к метаболическим сигналам и сигналам окружающей среды, а генные продукты могут участвовать в таких функциях, как восприятие сахара и передача сигналов в дополнение к транспорту ( Уильямс и др. ., 2000). Таким образом, поток сахаров через тонопласт может представлять собой стратегическую контрольную точку для интеграции циркадного и метаболического контроля цикла diel у видов САМ, которые накапливают растворимые сахара.
CO
2 обнаружение и сигнализацияБольшие, сильно вакуолизированные и плотно упакованные клетки листьев многих растений САМ представляют собой существенное препятствие для диффузии CO 2 как внутрь, так и из листа. Это потенциально может привести к 450-кратному падению p CO 2 в течение нескольких часов с момента максимального декарбоксилирования в фазе III до времени, когда происходит прямое поглощение атмосферного CO 2 во время фазы IV. (Максвелл и др. ., 1997). Таким образом, в течение цикла DIEL CAM сдвиги в режимах карбоксилирования C 3 и C 4 приводят к колебаниям внутренней концентрации CO 2 , которая может находиться в диапазоне от 0,011% до 5%. Увеличение p CO 2 считается основным внутренним фактором для контроля закрытия устьиц в фазе III (Bohn et al ., 2001). Ощущение p CO 2 во время перехода от фазы III к фазе IV также указывается на метаболическом уровне увеличением состояния активации Rubisco, который, как предполагалось, поддерживает снижение уровня CO 2 во время фазы IV (Максвелл и др. ., 1999: Максвелл, 2002; Griffiths et al ., 2002). Lüttge (2002, b ) также рассматривал роль p CO 2 в качестве сигнала для координации скорости потребления CO 2 Рубиско и ремобилизации яблочной кислоты из вакуоли во время фазы III CAM. Таким образом, зондирование и передача сигналов p CO 2 могут представлять собой еще один неотъемлемый компонент механизмов, которые синхронизируют потоки углерода в течение цикла diel в CAM-растениях.
Интеграция сигналов окружающей среды и метаболизма с циркадными часами
Механизм, лежащий в основе циркадных часов у всех организмов, основан на отложенных отрицательных петлях, регулирующих гены, участвующие в ядре осциллятора (Dunlap, 1999; Harmer et al ., 2001). Эти петли включают отрицательные регуляторы, которые подавляют свою собственную экспрессию, блокируя положительно действующие элементы (Harmer et al ., 2001; Young and Kay, 2001). Различные уровни контроля, включая посттранскрипционную регуляцию и связанные с ней взаимосвязанные петли обратной связи, обеспечивают механизмы, обеспечивающие стабильность и надежность часов в изменяющихся условиях, а также сбрасывают фазу часов с помощью сигналов окружающей среды, особенно световых (Shearman et al ., 2000). В Arabidopsis, циркадные часы включают набор генов, некоторые из которых являются хорошо охарактеризованными факторами транскрипции (например, регулятор псевдоответа TOC1: Время экспрессии CAB ) и факторы из семейства Myb ( CCA1 ). : Циркадные часы, связанные и LHY1 : Поздний удлиненный гипокотиль ). Эти факторы регулируют выражение друг друга в петле отрицательной обратной связи, что составляет примерно 24-часовой период.Отсутствие экспрессии TOC1 вызывает аритмию, тогда как гены CCA1 и LHY1 , которые имеют перекрывающиеся функции, участвуют в управлении периодом часов. Мутанты, лишенные экспрессии CCA1 и LHY1 , неспособны поддерживать устойчивые колебания ни в постоянном свете, ни в темноте (Alabadi et al ., 2002). Циркадная экспрессия этих часовых генов контролирует нисходящую экспрессию множества генов, участвующих во многих жизненно важных процессах, включая фотосинтетический метаболизм, движение листьев и цветение.В настоящее время ведется работа по идентификации генов-гомологов TOC1 , CCA1 и LHY1 в индуцибельном CAM-растении M. crystalinum (J Hartwell, личное сообщение). Полученные результаты помогут выяснить, как часы-контроль метаболических процессов задействованы при индукции CAM. Более того, исследования таких часовых генов могут выявить дополнительную роль часов в регуляции долгосрочной индукции CAM. У таких видов, как K. blossfeldiana и M.crystalinum, , долгосрочный контроль экспрессии CAM может быть коррелирован с изменением длины фотопериода (Brulfert et al. ., 1975; Edwards et al. ., 1996). Хотя этот переход развития к САМ может быть ускорен различными факторами окружающей среды (например, засухой, соленостью, ярким освещением), ощущая изменение продолжительности фотопериода, циркадные часы могут устанавливать метаболические и физиологические корректировки в ожидании изменений. в условиях окружающей среды (Тайби и др. ., 1995, 2002; Тайби и Кушман, 1999). Как у K. blofeldiana , так и у M. crystalinum фотопериодическая индукция САМ всегда сопровождается цветением и производством семян. В будущем будет интересно установить, контролируются ли долгосрочные (сезонные) ответы, такие как индукция CAM и цветение, а также краткосрочные (diel) ответы с помощью одних и тех же часов или наборов часовых генов.
Очевидно, что внешние (например, свет, температура) и внутренние факторы (метаболиты, гормоны) могут влиять как на амплитуду, так и на период основных ритмов, генерируемых часами в CAM-растениях.Такая модуляция необходима для оптимизации использования ресурсов в конкретном временном масштабе и для обеспечения координации развития CAM с сезонными изменениями. В настоящее время неясно, действуют ли эти внешние и внутренние факторы непосредственно на гены часов или действуют ниже по течению, увлекая метаболический выход часов. Однако мембрана тонопласта остается сильным кандидатом на интеграцию метаболического и циркадного контроля пути diel CAM. Lüttge (2000, 2002 a ) предположил, что биофизическое напряжение / расслабление тонопласта представляет собой основной осциллятор или кардиостимулятор для CAM.Другая возможная модель для гипотезы осциллятора тонопласта включает синтез пептида, который встроен в мембрану, где он имеет отрицательную обратную связь по собственному синтезу (Lüttge et al ., 2002 a ). Прямой циркадный контроль пептида, связанного с транспортом малата, пептида, который, в свою очередь, регулирует свою собственную экспрессию, может быть одним из возможных механизмов, с помощью которого тонопласт может интегрировать циркадные и метаболические сигналы для управления циклом САМ.
Эволюционные последствия и выводы
Два фундаментальных уровня контроля контролируют метаболические компоненты САМ. Управление с помощью циркадных часов устанавливает основные фазы CAM и обеспечивает соответствующую синхронизацию метаболических и транспортных процессов путем фазирования транскрипции определенных генов в определенное время в цикле день / ночь. Этот циркадный контроль перекрывается контролем метаболитов, действующим как на транскрипционном, так и на посттранскрипционном уровнях, что способствует фотосинтетической пластичности, унося выходной сигнал часов на колебания в окружающей среде.Возвратно-поступательные пулы органических кислот и углеводов играют центральную роль в метаболическом контроле САМ. Локализованные на мембране переносчики и сенсоры органических кислот, сахаров и промежуточных продуктов метаболизма, таким образом, занимают стратегические контрольные точки для интеграции циркадных и метаболических сигналов, которые синхронизируют и модулируют фазы диэль в ответ на окружающую среду. В частности, тонопласт играет ключевую роль в организации метаболизма в течение цикла день / ночь, контролируя поглощение и высвобождение органических кислот в цитозоль.Действительно, у некоторых видов CAM (например, Clusia ) дикие колебания количества транскриптов генов, кодирующих ключевые ферменты CAM (например, киназу PEPC), по-видимому, являются последствием круговорота метаболитов в тонопласте (рис.5).
Интегрированный механистический обзор цикла САМ, описанный выше, поднимает вопрос о том, как функции метаболического контроля САМ эволюционировали из фотосинтеза C 3 . Если CAM эволюционировал посредством серии дополнительных модификаций существующих функций, важно учитывать атрибуты CAM-предшественников, которые представляют предрасположенность к развитию как циркадных, так и метаболических компонентов этого пути.Считается, что начальный шаг в эволюции CAM включает в себя ночное поглощение респираторного CO 2 с помощью PEPC, особенности, которой, вероятно, способствовала анатомия суккулентных и крупных вакуолизированных листьев, типичная для видов CAM (Griffiths, 1989; Sage, 2002). ). Суккулентность, возможно, была ранее существовавшей чертой, которая улучшила гидравлический транспорт и емкость в местообитаниях с высокой потребностью в транспирации (Sage, 2002). Плотно упакованные клетки суккулентных листьев минимизировали бы утечку респираторного CO 2 из листа, в то время как увеличенная вакуоль была необходима для хранения фиксированного CO 2 в виде органических кислот в течение ночи.Так как вакуоль обычно занимает c . 97% объема клеток и цитоплазмы c . 1% в листьях растений САМ, цитоплазма могла легко быть затоплена вакуолярным содержимым, тем самым нарушая метаболический гомеостаз. Т.о., транспорт через тонопласт должен строго регулироваться, указывая тем самым, что ранним событием с точки зрения метаболического контроля CAM могло быть увеличение регуляторной способности транспортеров тонопласта. Это могло быть достигнуто за счет диверсификации набора транспортеров тонопластов (например,грамм. отдельные переносчики для притока и оттока органических кислот) вместе с изменением регуляторных свойств переносчиков. На молекулярном уровне такие качественные и количественные изменения в транспортерах тонопластов могли развиваться посредством дупликации генов с последующей специфической регуляцией одного или нескольких изогенов, как обычно предполагалось для нескольких основных метаболических компонентов CAM (Cushman and Bohnert, 1999). Важность вакуоли в поддержании клеточного гомеостаза может также сделать тонопласт ключевым (если не основным) компонентом циркадного осциллятора, который контролирует САМ.
При рассмотрении эволюционного происхождения циркадного контроля CAM также уместно рассмотреть, обладали ли предки CAM уже часы с всепроникающим контролем над метаболизмом. В то время как экспрессия циркадного гена в прокариотическом организме, Synechococcus , по-видимому, является универсальным феноменом (Liu et al. ., 1995), у высшего растения A. thaliana ~ 36% генома потенциально находится под транскрипционным контролем со стороны циркадные часы (Michael and McClung, 2003).Однако из генов Arabidopsis , которые, как известно, проявляют циркадные колебания в содержании транскриптов, большая часть (почти 70%) также напрямую реагирует на стресс окружающей среды (т.е. низкая температура, соль и засуха; Kreps et al ., 2002). Это наблюдение натолкнуло на предположение, что ритмическая экспрессия таких связанных со стрессом генов в ожидании предсказуемых изменений окружающей среды может подготовить растение к сопротивлению стрессу или наилучшим образом использовать потенциально ограничивающий ресурс (Eriksson and Millar, 2003).Поскольку обычно считается, что САМ возник в экстремальных условиях (Raven and Spicer, 1996), можно предположить, что предшественники САМ уже обладали часами с всепроникающим контролем над метаболизмом как средством поддержания гомеостаза в потенциально ограничивающих средах. Последующая эволюция пути могла происходить либо путем связывания большего количества / различных компонентов метаболического выхода с центральным осциллятором и / или посредством изменений в путях (и, возможно, механизмах), которые соединяют центральный осциллятор (ы) с метаболическим выходом.Будущие подходы к пониманию фотосинтетических характеристик растений CAM потребуют применения постгеномных технологий, включая транскриптомику, протеомику и метаболомику, чтобы идентифицировать гены и белки, которые контролируются часами, и определять роль метаболитов (например, органических кислот и сахаров) для синхронизации потоков остаточного углерода. Технология генных чипов в настоящее время применяется для анализа относительного вклада часов и контроля метаболитов у мутантов M.кристаллинум (Дж. Кушман, личное сообщение). Дальнейший прогресс требует разработки подходящей и поддающейся контролю системы трансформации для проверки гипотез о циркадных и метаболических сигналах, которые контролируют как экспрессию, так и синхронизацию метаболических компонентов, составляющих САМ.
Благодарности
Мы благодарны многим коллегам, которые внесли свой вклад в высказанные выше идеи, особенно Джону Кушману, Джеймсу Хартвеллу, Хью Ниммо и Эндрю Смиту.Наши исследования поддерживаются Советом по исследованиям окружающей среды Великобритании.
Рис. 1. Основные метаболические компоненты цикла САМ. Схематическое изображение (а), иллюстрирующее дневное (светлый фон) и ночное (темный фон) разделение основных метаболических и транспортных процессов для углерода. Распределение углеводов между ростом и хранением (будь то в виде растворимых сахаров в вакуоли или в виде крахмала в хлоропласте) является важным моментом контроля цикла диэфира.Конечными результатами этого простого разделения метаболизма (b) являются взаимно меняющиеся уровни органических кислот (в основном малата) и углеводов, которые могут составлять до 20% от сухой массы листа. Сплошная полоса на оси x указывает на темный период, а данные по метаболитам были собраны для Clusia fluminensis (Borland, неопубликованные результаты).
Рис. 1. Основные метаболические компоненты цикла САМ. Схематическое изображение (а), иллюстрирующее дневное (светлый фон) и ночное (темный фон) разделение основных метаболических и транспортных процессов для углерода.Распределение углеводов между ростом и хранением (будь то в виде растворимых сахаров в вакуоли или в виде крахмала в хлоропласте) является важным моментом контроля цикла диэфира. Конечными результатами этого простого разделения метаболизма (b) являются взаимно меняющиеся уровни органических кислот (в основном малата) и углеводов, которые могут составлять до 20% от сухой массы листа. Сплошная полоса на оси x указывает на темный период, а данные по метаболитам были собраны для Clusia fluminensis (Borland, неопубликованные результаты).
Рис. 2. Регулирование фосфо енол пируваткарбоксилазы в САМ растениях. Ген Ppc может быть быстро индуцирован факторами окружающей среды (водный стресс, солевой стресс и т. Д.) Или в качестве долгосрочной реакции на изменение фотопериода посредством контроля циркадным осциллятором. Ppc Транскрипты также колеблются на циркадной основе, но это изменение не приводит к изменениям количества белка PEPC в течение 24-часового цикла.Diel-регуляция PEPC контролируется посредством ночного фосфорилирования, катализируемого специальной киназой (PEPC-kinase). Ген Ppck может быть индуцирован солевым стрессом у некоторых видов ( Mesembryanthemum crystalinum, сорго) и контролируется циркадным осциллятором и / или метаболитами (органическими кислотами и сахарами). Транскрипты Ppck накапливаются в основном в течение ночи в CAM-растениях, что приводит к ночному накоплению PEPC-киназы и фосфорилированию PEPC. Фосфорилированный PEPC более активен, более чувствителен к активации глюкозо-6-фосфатом и триозофосфатом и менее чувствителен к ингибированию малатом.
Рис. 2. Регулирование фосфо енол пируваткарбоксилазы в САМ растениях. Ген Ppc может быть быстро индуцирован факторами окружающей среды (водный стресс, солевой стресс и т. Д.) Или в качестве долгосрочной реакции на изменение фотопериода посредством контроля циркадным осциллятором. Ppc Транскрипты также колеблются на циркадной основе, но это изменение не приводит к изменениям количества белка PEPC в течение 24-часового цикла. Diel-регуляция PEPC контролируется посредством ночного фосфорилирования, катализируемого специальной киназой (PEPC-kinase).Ген Ppck может быть индуцирован солевым стрессом у некоторых видов ( Mesembryanthemum crystalinum, сорго) и контролируется циркадным осциллятором и / или метаболитами (органическими кислотами и сахарами). Транскрипты Ppck накапливаются в основном в течение ночи в CAM-растениях, что приводит к ночному накоплению PEPC-киназы и фосфорилированию PEPC. Фосфорилированный PEPC более активен, более чувствителен к активации глюкозо-6-фосфатом и триозофосфатом и менее чувствителен к ингибированию малатом.
Рис. 3. Регуляция Diel и очевидный статус фосфорилирования PEPC у контрастирующих видов CAM. Чувствительность PEPC к малату (a) и содержание малата в листьях (b) в листьях Kalanchoë daigremontiana (закрашенные кружки), Clusia minor (белые кружки) и Tillandsia usneoides (незакрашенные ромбики), выращенных в идентичных контролируемых условиях с сплошная полоса на оси x , указывающая период темноты.Графики составлены на основе данных, представленных Borland и Griffiths (1997) и Haslam и др. . (2002).
Рис. 3. Регуляция Diel и очевидный статус фосфорилирования PEPC у контрастирующих видов CAM. Чувствительность PEPC к малату (a) и содержание малата в листьях (b) в листьях Kalanchoë daigremontiana (закрашенные кружки), Clusia minor (белые кружки) и Tillandsia usneoides (незакрашенные ромбики), выращенных в идентичных контролируемых условиях с сплошная полоса на оси x , указывающая период темноты.Графики составлены на основе данных, представленных Borland и Griffiths (1997) и Haslam и др. . (2002).
Рис. 4. Метаболитный контроль фосфорилирования РЕРС и суммарное поглощение CO 2 . Диаграммы Diel чистого поглощения CO 2 (а) листьями Kalanchoë daigremontiana при окружающем воздухе (контроль) или при поддержании в атмосфере N 2 в течение первых 6 часов (половина N 2 ) или в течение всего периода. 12 часов (полный N 2 ) темного периода (обозначено сплошной полосой на оси x ).Приложение в N 2 предотвращает накопление малата и последующее высвобождение из N 2 приводит к удвоению чистого поглощения CO 2 через PEPC по сравнению с контролями. Увеличение чистого поглощения CO 2 в листьях, обработанных N 2 , сопровождается увеличением активности (b) и экспрессии (данные не показаны) киназы PEPC, белка, ответственного за фосфорилирование PEPC. Данные были повторно взяты из Borland et al . (1999).
Рис.4. Метаболитный контроль фосфорилирования РЕРС и чистого поглощения CO 2 . Диаграммы Diel чистого поглощения CO 2 (а) листьями Kalanchoë daigremontiana при окружающем воздухе (контроль) или при поддержании в атмосфере N 2 в течение первых 6 часов (половина N 2 ) или в течение всего периода. 12 часов (полный N 2 ) темного периода (обозначено сплошной полосой на оси x ). Приложение в N 2 предотвращает накопление малата и последующее высвобождение из N 2 приводит к удвоению чистого поглощения CO 2 через PEPC по сравнению с контролями.Увеличение чистого поглощения CO 2 в листьях, обработанных N 2 , сопровождается увеличением активности (b) и экспрессии (данные не показаны) киназы PEPC, белка, ответственного за фосфорилирование PEPC. Данные были повторно взяты из Borland et al . (1999).
Рис. 5. Экспрессия Diel киназы РЕРС у видов Clusia . Дневные (3 часа и 6 часов) и ночные (15 часов и 18 часов) стабильные уровни транскриптов Ppck1 и Ubq1 (контроль) в листьях Clusia multiflora (C 3 видов), C. .aripoensis (C 3 ), C. minor (CAM) и C. rosea (CAM), выращенных в идентичных контролируемых условиях.
Рис. 5. Экспрессия Diel киназы PEPC у видов Clusia . Дневные (3 часа и 6 часов) и ночные (15 часов и 18 часов) стабильные уровни транскриптов Ppck1 и Ubq1 (контроль) в листьях Clusia multiflora (C 3 видов), C. . aripoensis (С 3 ), С.minor (CAM) и C. rosea (CAM), выращенные в идентичных контролируемых условиях.
Список литературы
Алабади Д , Яновский М.Ю., Мас П, Хармер С.Л., Кей С.А.
2002
. Критическая роль CCA1 и LHY1 в поддержании циркадной ритмичности у Arabidopsis .Current Biology
12
,757
–761.Варфоломей ДП , Кадзимин СБ.
1977
. Ананас. В: Alvim P de T, Kozlowski TT, eds. Экофизиология тропических культур . Нью-Йорк: Academic Press,113
–156.Bohn A , Geist A, Rascher U, Lüttge U.
2001
. Ответы на различные внешние световые ритмы циркадным ритмом метаболизма крассуловой кислоты в Kalanchoë daigremontiana .Завод, клетки и окружающая среда
24
,811
–820.Borland AM , Dodd AN.
2002
. Разделение углеводов на заводах CAM: устранение потенциальных конфликтов интересов.Функциональная биология растений
29
,707
–716.Borland AM , Griffiths H.
1997
. Сравнительное исследование регуляции процессов карбоксилирования C 3 и C 4 в конститутивном CAM-растении Kalanchoë daigremontiana и промежуточном C 3 -CAM Clusia minor .Планта
201
,368
–378.Borland AM , Griffiths H, Broadmeadow MSJ, Fordham MC, Maxwell C.
1993
. Кратковременные изменения в дискриминации изотопов углерода в промежуточном продукте C 3 -CAM Clusia minor L., растущем в Тринидаде.Oecologia
95
,444
–453.Borland AM , Griffiths H, Maxwell C, Broadmeadow MSJ, Fordham MC.
1996
. Индукция САМ у Clusia minor L. во время перехода от влажного к засушливому сезону в Тринидаде: роль видообразования органических кислот и декарбоксилирования.Завод, клетки и окружающая среда
19
,655
–664.Borland AM , Hartwell J, Jenkins GI, Wilkins MB, Nimmo HG.
1999
. Контроль метаболитов преобладает над циркадной регуляцией киназы PEPc и фиксацией CO 2 в метаболизме крассулейной кислоты (CAM).Физиология растений
121
,889
–896.Borland AM , Tecsi L, Leegood RC, Walker RP.
1998
. Индуцируемость метаболизма крассулоидной кислоты (CAM) у видов Clusia ; физиологическая / биохимическая характеристика и межклеточная локализация процессов карбоксилирования у трех видов, которые показывают разную степень САМ.Планта
205
,342
–351.Боксолл SF , Bohnert HJ, Cushman JC, Nimmo HG, Hartwell J.
2001
. Циркадные часы и метаболизм крассулоидной кислоты в Mesembryanthemum crystalinum . Плакат S18 ‐ D12. 12-й Международный конгресс фотосинтеза , Брисбен.Боксолл SF , Bohnert HJ, Cushman JC, Nimmo HG, Hartwell J.
2002
. На пути к пониманию молекулярных событий, лежащих в основе циркадного контроля фиксации CO 2 в метаболизме крассулейной кислоты с использованием модельного растения CAM Mesembryanthemum crystalinum .Плакат 520. ASPB Plant Biology 2002 , Денвер.Бранко JB , Agarie S, Elliott S, Borland AM, Cushman JC.
2003
. Выделение и анализ CAM-дефектных мутантов у обыкновенного ледяного растения, Mesembryanthemum crystalinum . Американское общество биологов растений. Плакат 403. ASPB Биология растений 2003 , Гонолулу, Гавайи.Брюльферт J , Guerrier D, Queiroz O.
1975
.Фотопериодизм и ферментные ритмы. Кинетические характеристики фотопериодической индукции метаболизма крассулоидной кислоты.Planta
125
,987
–995.Картер PJ , Nimmo HG, Fewson CA, Wilkins MB.
1991
. Циркадные ритмы активности протеинкиназы растений.EMBO Journal
10
,2063
–2068.Chollet R , Vidal J, O’Leary MH.
1996
. Phospho енол пируваткарбоксилаза: широко распространенный, строго регулируемый фермент в растениях.Ежегодный обзор физиологии растений и молекулярной биологии растений
47
,273
–298.Кристофер JT , Holtum JAM.
1996
. Закономерности распределения углеводов в листьях видов крассулообразных с кислотным метаболизмом при раскислении.Физиология растений
112
,393
–399.Cushman JC , Bohnert HJ.
1999
. Метаболизм крассуловой кислоты: молекулярная генетика.Ежегодный обзор физиологии растений и молекулярной биологии растений
50
,305
–332.Cushman JC , Borland AM.
2002
. Индукция метаболизма крассулоидной кислоты за счет ограничения воды.Завод, клетки и окружающая среда
25
,297
–312.Cushman JC , Michalowski CB, Bohnert HJ.
1990
. Контроль развития индуцируемости метаболизма крассулоидной кислоты солевым стрессом у обыкновенного ледяного растения.Физиология растений
94
,1137
–1142.DeRocher EJ , Bohnert HJ.
1993
. Стресс, связанный с развитием и окружающей средой, использует разные механизмы в экспрессии семейства генов растений.Заводская ячейка
5
,1611
–1625.Додд AN , Borland AM, Haslam RP, Griffiths H, Maxwell K.
2002
. Метаболизм крассуловой кислоты: пластичный, фантастический.Журнал экспериментальной ботаники
53
,569
–580.Додд AN , Griffiths H, Taybi T, Cushman JC, Borland AM.
2003
.Интеграция метаболизма крахмала с циркадным ритмом и регуляцией CAM в окружающей среде в Mesembryanthemum crystalinum .Планта
216
,789
–797.Данлэп JC .
1999
. Молекулярные основы циркадных часов.Ячейка
96
,271
–290.Эдвардс GE , Dai Z, Cheng SH, Ku MSB.
1996
.Факторы, влияющие на индукцию метаболизма крассуловой кислоты в Mesembryanthemum crystalinum . В: Winter K, Smith JAC, ред. Метаболизм крассуловой кислоты. Биохимия, экофизиология и эволюция . Берлин: Springer ‐ Verlag,119
–143.Emmerlich V , Linka N, Reinhold T, Hurth MA, Traub M, Martinoia E, Neuhaus HE.
2003
. Растительный гомолог человеческого натрия / дикарбонового котранспортера представляет собой вакуолярный малатный носитель.Proceedings of the National Academy of Sciences, USA
(в печати).Эрикссон ME , Миллар А.Дж.
2003
. Циркадные часы. Лучший друг растения в мире прядения.Физиология растений
132
,732
–738.Гейгер DR , Шей W ‐ J, Yu X ‐ M.
1995
. Фотосинтетический метаболизм и транслокация углерода у мутантов дикого типа и с дефицитом крахмала Nicotiana sylvestris L.Физиология растений
107
,507
–514.Гриффитс H .
1989
. Механизмы концентрации углекислого газа и эволюция САМ в сосудистых эпифитах. В: Lüttge U, ed. Сосудистые растения как эпифиты . Берлин: Springer ‐ Verlag,42
–86.Griffiths H , Helliker B, Roberts A, Haslam RP, Girnus J, Robe WE, Borland AM, Максвелл К.
2002
. Регулирование активности Rubisco у растений САМ: лучше поздно, чем никогда.Функциональная биология растений
29
,689
–696Hafke JB , Hafke Y, Smith JAC, Lüttge U, Thiel G.
2003
. Поглощение вакуолярного малата опосредуется анион-селективным внутренним выпрямителем.Заводской журнал
35
,116
–128.Harmer SL , Hogenesch JB, Straume M, Chang H ‐ S, Han B, Zhu T, Wang Z, Kreps JA, Kay SA.
2000
. Организованная транскрипция ключевых путей в Arabidopsis по циркадным часам.Наука
290
,2110
–2113.Хармер SL , Panda S, Kay SA.
2001
. Молекулярные основы циркадных ритмов.Ежегодный обзор клеточной биологии и биологии развития
17
,215
–253.Hartwell J , Gill A, Nimmo GA, Wilkins MB, Jenkins GI, Nimmo HG.
1999
. Phospho енол киназа пируваткарбоксилазы представляет собой новую протеинкиназу, уровень экспрессии которой регулируется.Заводской журнал
20
,333
–342.Hartwell J , Smith LH, Wilkins MB, Jenkins GI, Nimmo HG.
1996
. Киназа фосфо енола пируваткарбоксилазы высших растений регулируется на уровне транслируемой мРНК в ответ на свет или циркадный ритм.Заводской журнал
10
,1071
–1078.Хаслам R , Borland A, Griffiths H.
2002
. Кратковременная пластичность экспрессии метаболизма крассуловой кислоты в эпифитной бромелии Tillandsia usneoides L.Функциональная биология растений
29
,749
–756Häusler RE , Baur J, Тейхманн Т., Эйкс М., Фишер К.Л., Флюгге У-И, Шуберт С., Вебер А., Фишер К.
2000
. Переносчики пластидных метаболитов и их физиологические функции в индуцибельном растении метаболизма крассулообразной кислоты Mesembryanthemum crisinum .Заводской журнал
24
,285
–296.Крепс JA , Wu Y, Chang H ‐ S, Zhu T, Wang X, Harper JF.
2002
. Изменения транскриптома Arabidopsis в ответ на солевой, осмотический и холодовой стресс.Физиология растений
130
,2129
–2141.Li B , Geiger DR, Shieh W ‐ J.
1992
. Доказательства циркадной регуляции синтеза крахмала и сахарозы в листьях сахарной свеклы.Физиология растений
99
,1393
–1399.Лю Y , Циноремас Н.Ф., Джонсон СН, Лебедека Н.В., Голден СС, Ишиура М., Кондо Т.
1995
.Циркадная оркестровка экспрессии генов у цианобактерий.Гены и развитие
9
,1469
–1478.Лю Z , Taub CC, McClung CR.
1996
. Идентификация минимального промотора Arabidopsis thaliana рибулозо-1,5-бифосфаткарбоксилазы / оксигеназы активазы (RCA), регулируемого светом и циркадными часами.Физиология растений
112
,43
–51.Lüttge U .
2000
. Тонопласт функционирует как главный переключатель циркадной регуляции метаболизма кислоты крассулей.Планта
211
,761
–769.Lüttge U .
2002
а . Циркадная ритмичность: «Биологические часы» — оборудование или программное обеспечение?Прогресс ботаники
64
,277
–319.Lüttge U .
2002
b . CO 2 -концентрирование: последствия для метаболизма крассулевой кислоты.Журнал экспериментальной ботаники
53
,2131
–2142.McRae SR , Christopher JT, Smith JAC, Holtum JAM.
2002
. Транспорт сахарозы через вакуолярную мембрану Ananas comosus .Функциональная биология растений
29
,717
–724.Максвелл К .
2002
. Полезна устойчивость: суточные паттерны фотосинтеза в C 3 и эпифитные бромелии, метаболизм крассуловой кислоты.Функциональная биология растений
29
,679
–687.Максвелл K , von Caemmerer S, Evans JR.
1997
. Является ли низкая внутренняя проводимость к CO 2 следствием сочности растений с метаболизмом крассулевой кислоты?Австралийский журнал физиологии растений
24
,777
–786.Максвелл К , Борланд А.М., Хаслам Р.П., Хелликер Б.Р., Робертс А., Гриффитс Х.
1999
. Модуляция активности Rubisco во время суточных фаз метаболизма кислоты крассулейных растений Kalanchoë daigremontiana .Физиология растений
121
,849
–856.Майкл TP , McClung CR.
2003
. Захват энхансеров выявляет широко распространенный контроль транскрипции суточных часов у Arabidopsis .Физиология растений
132
,629
–639.Nimmo GA , Nimmo HG, Hamilton ID, Fewson CA, Wilkins MB.
1986
. Очистка фосфорилированной ночной формы и дефосфорилированной дневной формы фосфо енол пируваткарбоксилазы из Bryophylum fedtschenkoi .Биохимический журнал
239
,213
–220.Nimmo GA , Wilkins MB, Fewson CA, Nimmo HG.
1987
. Устойчивый циркадный ритм в состоянии фосфорилирования фосфо енол пируваткарбоксилазы из листьев Bryophylum fedtschenkoi и его чувствительность к ингибированию малатом.Planta
170
,408
–415.Ниммо HG .
2000
. Регулирование фосфо- енол пируваткарбоксилазы в САМ-растениях.Тенденции в растениеводстве
5
,75
–80.Нобель ПС .
1996
. Высокая урожайность некоторых агрономических видов САМ. В: Winter K, Smith JAC, ред. Метаболизм крассуловой кислоты. Биохимия, экофизиология и эволюция . Берлин: Springer ‐ Verlag,255
–265.Rascher U , Hütt MT, Siebke K, Osmond CB, Beck F, Lüttge U.
2001
. Пространственно-временные вариации метаболизма в циркадном ритме растений: биологические часы как совокупность связанных индивидуальных осцилляторов.Proceedings of the National Academy of Sciences, USA
98
,11801
–11805.Raven JA , Spicer RA.
1996
. Эволюция метаболизма крассулоидной кислоты. В: Winter K, Smith JAC, ред. Метаболизм крассуловой кислоты. Биохимия, экофизиология и эволюция . Берлин: Springer ‐ Verlag,360
–385.Робертс A , Borland AM, Griffiths H.
1997
. Дискриминационные процессы и сдвиги в карбоксилировании на фазах метаболизма крассулообразной кислоты.Физиология растений
113
,1283
–1292.Рокель B , Blasius B, Beck F, Ratajcek R, Lüttge U.
1997
. Эндогенные колебания количества транскриптов субъединицы c V-АТФазы Mesembryanthemum crisinum с резонансами гармонической частоты при непрерывном освещении.Письма о клеточной и молекулярной биологии
2
,69
–76.Шалфей РФ .
2002
. Несовместимы ли метаболизм крассуловой кислоты и фотосинтез C 4 ?Функциональная биология растений
29
,775
–785.Шаффер R , Ландграф Дж., Акцерби М., Саймон В., Ларсон М., Висман Э.
2001
. Анализ микрочипов суточных и регулируемых циркадным ритмом генов в Arabidopsis .Заводская ячейка
13
,113
–123.Shearman LP , Sriram S, Weaver DR, и др. .
2000
. Взаимодействующие молекулярные петли в циркадных часах млекопитающих.Наука
288
,1013
–1019Смикенс S .
2000
. Передача сигнала, индуцированная сахаром, у растений.Ежегодный обзор физиологии растений и молекулярной биологии растений
51
,49
–81.Smith JAC , Ingram J, Tsiantis MS, Barkla BJ, Bartholomew DM, Bettey M, Pantoja O, Pennington AJ.
1996
. Транспорт через вакуолярную мембрану у растений САМ. В: Winter K, Smith JAC, ред. Метаболизм крассуловой кислоты. Биохимия, экофизиология и эволюция . Берлин: Springer ‐ Verlag,53
–71.Ститт М , Мюллер К., Мэтт П., Гибон И., Карилло П., Моркуенде Р., Шайбл В-Р, Крапп А.
2002
. Шаги к интегрированному взгляду на метаболизм азота.Журнал экспериментальной ботаники
53
,959
–970.Тайби Т , Cushman JC.
1999
. Сигнальные события, ведущие к индукции метаболизма крассулоидной кислоты у обыкновенного ледяного растения.Физиология растений
121
,545
–555.Тайби T , Cushman JC, Borland AM.
2002
. Экологическая, гормональная и циркадная регуляция экспрессии метаболизма крассулоидной кислоты.Функциональная биология растений
29
,669
–678.Тайби Т , Ниммо HG, Borland AM.
2004
. Экспрессия генов фосфо- енол пируваткарбоксилазы (PEPC) и PEPC-киназы: влияние на генотипическую способность и фенотипическую пластичность в экспрессии метаболизма крассулатовой кислоты.Физиология растений
(в печати).Тайби Т , Патил С, Чолле Р, Кушман Дж.
2000
. Минимальная серин / треониновая протеинкиназа циркадно регулирует активность фосфо енол пируваткарбоксилазы в листьях обыкновенного ледяного растения, индуцированных метаболизмом кислоты крассулосовых.Физиология растений
123
,1471
–1481.Taybi T , Sotta B, Gehrig HH, Güclü S, Kluge M, Brulfert J.
1995
. Дифференциальные эффекты абсцизовой кислоты на фосфо енол пируваткарбоксилазу и работу САМ в Kalanchoë blossfeldiana .Botanica Acta
198
,240
–246.Уэбб AAR .
2003
. Физиология циркадных ритмов растений.Новый фитолог
160
,281
–304.Уилкинс МБ .
1992
. Циркадные ритмы: их происхождение и контроль.Новый фитолог
121
,347
–375.Уильямс LE , Lemoine R, Sauer N.
2000
. Транспортеры сахара у высших растений — разнообразие ролей и сложная регуляция.Тенденции в растениеводстве
5
,283
–290.Зима K , Smith JAC.
1996
а .Таксономическое распределение метаболизма крассулообразной кислоты. В: Winter K, Smith JAC, ред. Метаболизм крассуловой кислоты. Биохимия, экофизиология и эволюция . Берлин: Springer ‐ Verlag,1
–13.Зима K , Smith JAC.
1996
b . Метаболизм крассуловой кислоты: современное состояние и перспективы. В: Winter K, Smith JAC, ред. Метаболизм крассуловой кислоты. Биохимия, экофизиология и эволюция .Берлин: Springer ‐ Verlag,389
–426.Wyka TP , Lüttge U.
2003
. Вклад карбоксилирования C 3 в циркадный ритм поглощения углекислого газа в растении с метаболизмом крассуловой кислоты Kalanchoë daigremontiana .Журнал экспериментальной ботаники
54
,1471
–1480.Янг МВт , Kay SA.
2001
.Часовые пояса: сравнительная генетика циркадных часов.Nature Reviews Genetics
2
,702
–715.Общество экспериментальной биологии
Влияние циркадного ритма на метаболические процессы и регуляцию энергетического баланса — FullText — Annals of Nutrition and Metabolism 2019, Vol. 74, № 4
Абстрактные
Предпосылки: Система суточного времени или циркадные часы играет решающую роль во многих биологических процессах, таких как цикл сна и бодрствования, секреция гормонов, здоровье сердечно-сосудистой системы, гомеостаз глюкозы и регулирование температуры тела.Энергетический баланс также является одним из важнейших краеугольных камней метаболических процессов, тогда как энергетический дисбаланс связан со многими заболеваниями (например, ожирением, диабетом, сердечно-сосудистыми заболеваниями). Циркадные часы являются основным регулятором метаболизма, и этот анализ дает обзор двунаправленного влияния циркадного ритма на метаболические процессы и энергетический баланс. Резюме: Система суточного времени или циркадные часы играет решающую роль во многих биологических процессах, но увеличение активности, которая работает круглосуточно и без выходных, и обычное использование телевидения, Интернета и мобильных телефонов почти 24 часа в сутки приводит к тому, что постепенное уменьшение адекватного времени сна.Согласно недавним исследованиям, долгосрочные нарушения циркадного ритма связаны со многими патологическими состояниями, такими как преждевременная смертность, ожирение, нарушение толерантности к глюкозе, диабет, психические расстройства, тревога, депрессия и прогрессирование рака, тогда как краткосрочные нарушения связаны с ухудшением самочувствия. , утомляемость и потеря концентрации. В этом обзоре были изучены циркадные ритмы метаболических процессов и их влияние на энергетический баланс. Ключевые сообщения: Циркадный ритм имеет двунаправленное взаимодействие почти со всеми метаболическими процессами.Следовательно, понимание основной причины, влияющей на циркадные часы, и создание рекомендаций по лечению с использованием циркадного ритма может повысить эффективность лечения заболевания. Хронофармакология, хроническое питание и хроноупражнения — это новые подходы к лечению метаболического баланса.
© 2019 S. Karger AG, Базель
Введение
Термин «циркадный» образован от двух латинских слов: circa (приблизительно) и dies (день), обозначающих приблизительно «один день» [1].Циркадный ритм также называется биологическими / циркадными часами [2] и относится к поведенческим, физиологическим и молекулярным изменениям с продолжительностью цикла около 24 часов [3]. Циркадные часы можно разделить на 2 части: центральные часы, расположенные в супрахиазматическом ядре (SCN) гипоталамуса, которое принимает световые сигналы, и периферические часы, находящиеся в различных тканях по всему телу. Периферические часы играют неотъемлемую и уникальную роль в каждой из своих тканей, управляя циркадной экспрессией определенных генов, участвующих в различных физиологических функциях [4].
Основным стимулом для SCN является свет [5]. Однако у слепых людей есть циркадные циклы (например, циклы сна и бодрствования), и этот цикл длится более 24 часов. Это открытие привело к мысли, что другие стимулы, помимо света, могут действовать как стимул для биологических часов человека [6]. Обзорное исследование показало, что температура, гормоны, питательные вещества, распределение питательных веществ, некоторые питательные вещества (только, например, глюкоза, аминокислоты, этанол и ретиноевая кислота), состояние кормления / голодания, состояние сна-бодрствования, физическая активность являются эффективными стимулами для циркадный цикл в различных периферических путях [5].Интересно, что сообщалось, что наблюдались различия во влиянии циркадного ритма у мужчин по сравнению с женщинами [7].
В основном цикл сна и бодрствования регулирует циркадный ритм. Однако в современном мире рост активности, которая осуществляется круглосуточно и без выходных, и обычное использование телевидения, Интернета и мобильных телефонов почти 24 часа в сутки, приводит к постепенному сокращению необходимого времени для сна [8]. Эпидемиологическое исследование показало, что продолжительность ночного сна уменьшилась на 18 минут за последние 30 лет [9].Ухудшение цикла сна-бодрствования, особенно у здоровых людей, может быть основной причиной многих заболеваний, таких как преждевременная смертность, ожирение, нарушение толерантности к глюкозе, диабет, психические расстройства, тревога, депрессия и прогрессирование рака, усталость и потеря. концентрации [10, 11]. Другое эпидемиологическое исследование показало, что работа не менее 3 ночей в месяц в течение 15 и более лет может увеличить риск рака прямой кишки у женщин [12]. У крыс нарушение циркадных обмороков ускоряет развитие диабета, вызывая функцию и потерю массы бета-клеток [13].В экспериментальном исследовании нарушение циркадного ритма приводило к поражению сердечно-сосудистой системы и почек у хомяков [14]. В исследовании, проведенном с пациентами с шизофренией, участники имели тяжелые циркадные расстройства сна и бодрствования, хотя их настроение, психический статус и психотические эпизоды были стабильными [15]. В этом обзоре изучалась роль циркадного ритма в метаболических процессах и двунаправленное влияние циркадного ритма на энергетический баланс. В соответствии с этой целью были проанализированы исследования, опубликованные с 2000 по 2018 год.Кроме того, были добавлены 3 важные статьи, опубликованные в 1984, 1993 и 1996 годах. В основном для сканирования научных статей использовались базы данных Google Academic (библиографическая база данных), PubMed, Scopus, Web of Science и Science Direct. Кроме того, были просканированы ресурсы Центральной библиотеки Университета Гази. Такие термины, как «циркадные часы или циркадный ритм» или «биологические часы» и «гормоны или метаболизм, или энергетический баланс, или расход энергии, или термогенез, метаболический гомеостаз или метаболическая регуляция, или гены часов, или диетический состав, режим питания или время приема пищи или физический активность, упражнения или спорт » были использованы в качестве ключевых слов.В результате сканирования было найдено 4 456 статей. После проверки заголовков и аннотаций были исключены повторяющиеся статьи и статьи, опубликованные не на английском языке. Окончательное количество статей составило 82, которые были бесплатными и доступными в виде полных текстов. Кроме того, главы книги также использовались для изучения предмета.
Циркадная синхронизация в метаболическом гомеостазе
Многие исследования объясняют взаимосвязь между физиологией человека, некоторыми заболеваниями и циркадным ритмом [12-15].Метаболический гомеостаз — важный компонент, регулирующий энергетический обмен, особенно в жировой ткани. Жировая ткань является центральным метаболическим органом, который регулирует энергетический гомеостаз всего тела. Белая жировая ткань функционирует как ключевой резервуар энергии для других органов, тогда как коричневая жировая ткань накапливает липиды для индуцированного холодом адаптивного термогенеза. Жировая ткань секретирует различные гормоны, цитокины и метаболиты (называемые адипокинами), которые контролируют системный энергетический баланс, регулируя сигналы аппетита от центральной нервной системы, а также метаболическую активность в периферических тканях [16].Например, лептин имеет специфические рецепторы на гипоталамусе и высвобождается из основных адипоцитов. Этот гормон играет регулирующую роль в энергетическом обмене, увеличивая активацию симпатической нервной системы и увеличивая термогенез за счет увеличения гормонов щитовидной железы. В термогенезе белок UCP (разобщающий) подавляет синтез АТФ в митохондриях, позволяя потреблять энергию в виде тепла. Лептин увеличивает уровень гормонов щитовидной железы и активацию симпатической центральной нервной системы, что приводит к большему образованию UCP и, следовательно, большему потреблению энергии [17].Высвобождение гормона лептина происходит в циркадном цикле, а пик уровня лептина в сыворотке приходится на ночь [18]. Таким образом, нарушение циркадного баланса может косвенно влиять на секрецию лептина, термогенез и энергетический гомеостаз.
Напротив, некоторые гормоны, выделяемые гипоталамусом, проявляют большую активность ночью. Самый яркий пример этого — «гормон роста». Уровень гормона роста достигает пика между 2:00 и 4:00 утра.Поэтому необходимо уделять особое внимание режиму сна детей [19].
Кортизол — стероидный гормон, секретируемый надпочечниками. Он регулирует многие метаболические процессы, такие как гликогенолиз, липолиз и протеолиз [20]. Количество и частота секреции кортизола регулируются циркадным ритмом. Концентрация кортизола в кровообращении достигает пика прямо перед пробуждением утром. Кортизол постепенно снижается в течение дня.Наименьшего уровня он достигает во время сна после полуночи [21]. Кортизол — это главный гормон, регулирующий метаболические процессы в организме. Это увеличивает использование кортизола, глюкозы, свободных жирных кислот и аминокислот из эндогенных запасов топлива. Следовательно, высокий уровень кортизола действует как катаболический гормон, который снижает мышечную массу и мышечную массу, а также увеличивает потребление энергии [22]. Кроме того, толерантность к глюкозе и секреция инсулина меняются в течение дня. При естественном ходе метаболизма чувствительность к инсулину и секреция инсулина снижаются ночью (особенно между 3:00 и 5:00 a.м) по сравнению с утренними часами. Этот метаболический процесс, феномен рассвета, подчеркивает влияние контроля циркадных ритмов на метаболизм глюкозы [23]. В естественных физиологических процессах организма гормоны, действующие как антагонисты инсулина (особенно гормон роста), проявляют гиперинсулинемическую активность из-за снижения секреции инсулина между 3:00 и 5:00 утра, так что уровень сахара в крови возвращается к норме. Этому противодействует дополнительная физиологическая секреция инсулина у людей, не страдающих диабетом или инсулинозависимых.И наоборот, когда высвобождение инсулина нарушено, действие гормона роста, высвобождаемого в течение ночи, особенно у пациентов с диабетом, не может быть уменьшено. Это приводит к патологическому циркадному ритму, который может привести к утренней гипергликемии независимо от режима питания [24].
Мелатонин — важный гормон в циркадной синхронизации. Этот гормон участвует во многих биологических и физиологических регуляторах организма. Это эффективный гормон биоритма человека (циркадного ритма).Основная роль этого гормона — поддерживать биологические часы и регулировать ритм тела [25]. Синтез и высвобождение мелатонина стимулируются в темноте, ночью, тогда как днем он подавляется светом [1]. Особенно между 23:00. в 5:00 секреция мелатонина достигает пика, и его концентрация в крови увеличивается в 3–10 раз [25]. Однако воздействие света в ночное время вызывает снижение уровня мелатонина в плазме [1].
На метаболизм липидов также влияют циркадные колебания.Исследования показывают, что многие белки, связанные с метаболизмом липидов (например, ApoB, ApoA1 и ApoA4), белок кишечного микросомального транспорта триглицеридов и белок, связывающий кишечные жирные кислоты, демонстрируют изменения в течение дня [26–28]. Кроме того, исследования на мышах показывают, что абсорбция холестерина и липидов в темной фазе выше, чем в светлой фазе [28]. Некоторые продукты липидного обмена также демонстрируют циркадный ритм. Например, циркулирующие неэтерифицированные жирные кислоты у людей выше ночью из-за повышенной липолитической активности [29].Кроме того, эпизоды инфаркта миокарда и астма связаны с циркадным циклом метаболизма. Эти приступы достигают пика ночью или рано утром [30, 31]. В ранние утренние часы сердечно-сосудистая система улучшается в ответ на активацию активности симпатических нервов, а в вечерние часы — артериальное давление и пик пульса. Сердечные приступы, особенно рано утром или вечером, могут быть вызваны этим суточным ритмом сердечно-сосудистой системы.Суточный ритм гормонов и некоторые метаболические процессы показаны на рисунке 1.
Рис. 1.
Суточный ритм гормонов и некоторых метаболических процессов. Высвобождение гормона лептина происходит в циркадном цикле, и пик уровня лептина в сыворотке крови приходится на ночь. Уровень гормона роста достигает пика между 02:00 и 04:00 утра.Концентрация кортизола в кровообращении достигает пика перед пробуждением утром. Снижение секреции инсулина ночью (особенно с 03:00 до 05:00 a.м.). Синтез и высвобождение мелатонина стимулируются ночью в темноте, а днем он подавляется светом. Поглощение липидов в темной фазе выше, чем в светлой фазе. Пик инфаркта миокарда ночью или рано утром.
Метаболическая регуляция циркадных ритмов
У млекопитающих циркадные ритмы контролируются главным образом SCN, которые называются главными часами. Супрахиазматические ядра состоят из множества одноклеточных циркадных осцилляторов, расположенных в передней области гипоталамуса головного мозга и производящих согласованные циркадные сигналы при синхронизации [32].SCN активируется через нервный пучок, называемый «ретиногипоталамический тракт». Таким образом, SCN регулирует биологические часы у живых существ и способствует физиологическим процессам, стимулируя другие области мозга [2].
Супрахиазматический синапс ядра непосредственно с вентральной и дорсальной субвентрикулярной областями, клеточными телами, расположенными в вентральной и дорсальной субвентрикулярной областях, и дорсомедиальным гипоталамусом. Несмотря на то, что эти области взаимодействуют друг с другом, нейроны, расположенные в дорсальной наджелудочковой области, более эффективны в регуляции термогенеза, тогда как вентральная наджелудочковая область в основном играет роль в регуляции циклов сна-бодрствования и активности.Кроме того, паравентрикулярный гипоталамус отвечает за высвобождение кортикостероидов, тогда как боковой гипоталамус отвечает за питание и бдительность [33].
Хотя SCN функционирует как основные биологические часы метаболизма, исследования, проведенные в 2000-х годах, показали, что автономные циркадные осцилляторы, присутствующие в периферических органах и тканях, таких как печень, кишечник, сердце и сетчатка, вносят вклад в метаболические процессы через клеточные гены часов в этих органах / тканях [32, 34, 35].Супрахиазматические ядра играют важную роль в регуляции метаболизма глюкозы. Чувствительность к инсулину и поглощение глюкозы нарушены у крыс с повреждениями в их SCN [36]. Ухудшение циркадных часов может привести к нарушению секреции инсулина и гипоинсулинемии [29]. Связанные с циркадным ритмом белки CLOCK и BMAL1 участвуют в производстве и высвобождении инсулина, связываясь с регулирующими циркадный ритм дистальными областями β-клеток поджелудочной железы [37]. И диабет 1-го и 2-го типа имеет недостаточность или отсутствие инсулина из-за повреждения β-клеток.Следовательно, не следует упускать из виду роль циркадного ритма в этиологии диабета с точки зрения повреждения β-клеток [38]. Гены часов и задачи, регулирующие циркадные ритмы у млекопитающих, сведены в Таблицу 1 [2].
Таблица 1.
Гены циркадного ритма и их роли [2]
У млекопитающих гомеостаз циркадных часов обеспечивается механизмами обратной связи (отрицательная) и прямой (положительная), влияющими на транскрипцию, трансляцию и посттрансляционную трансляцию. события [5, 29].Паттерн транскрипционной обратной связи опосредуется белками Cry1, Cry2, Per1 и Per2. Белки CLOCK и BMAL1 связываются с областью E-промотора генов Per и Cry и индуцируют их экспрессию. На более поздней стадии Per и Cry гетеродимеризуются и перемещаются из цитоплазмы в ядро, чтобы ингибировать CLOCK / BMAL1-индуцированную экспрессию генов [37, 39]. Этот механизм отрицательной обратной связи важен для эффективной работы циркадной системы.
Циркадный ритм и энергетический гомеостаз
Энергетический метаболизм регулируется многими гормонами, ферментами и транспортными системами, а циркадный ритм эффективно модулирует их экспрессию, секрецию и / или активацию [40].Метаболические пути, обеспечивающие энергетический гомеостаз, координируются метаболитами, которые предлагают изменения и активно подготавливают молекулярную среду, а также системы острой сигнализации, которые мгновенно реагируют на изменения циркадных часов [41].
Рецепторы гормонов, взаимодействия между генами и внутриклеточные реакции окисления / восстановления регулируют энергетический обмен на клеточном уровне. Эти пути имеют важное взаимодействие с часами биологического метаболизма.Циркадные часы могут влиять на функцию рецепторов гормонов (рецептор, активируемый пролифератором пероксисом [PPAR] α, PPARγ и REV-ERBα) и некоторых генов на клеточном уровне (сиртруин) [41, 42].
Рецепторы ядерных гормонов (PPARα, PPARγ, REV-ERBα, RORα, HNF4α, TRα и NURR1) и лиганды, присутствующие в метаболических тканях, действуют как сенсоры, объединяющие циркадные и метаболические пути [41]. Например, PPAR являются связующим звеном между циркадными часами и энергетическим метаболизмом. PPARγ локализуется в жировой ткани и активирует факторы транскрипции, которые увеличивают липогенез и накопление липидов.PPARα из ядерных рецепторов гормонов запускает кетогенез и окисление жирных кислот печени в ответ на голодание. PPARδ является наиболее распространенным рецептором ядерных клеток в организме и способен коррелировать ежедневные изменения температуры тела с циркадными часами [43].
Ритмическая экспрессия и активация метаболических путей в основном связаны с координацией часовых генов (BMAL1, Per2, Per1, Per3, Cry1 и Cry2) в печени и жировой ткани. Существует связь между белком BMAL1, липогенными путями и механизмами клеточных часов.REV-ERBα (репрессор транскрипции BMAL1) и RORα (положительный регулятор BMAL1) являются рецепторами ядерных гормонов, регулирующими липогенез. Более того, оба модулируются ЧАСОМ: BMAL1. PPARα играет роль в метаболизме липидов и липопротеинов. PPARα напрямую связывается с промоторной областью белка BMAL1, который регулирует экспрессию PPARα через гетеродимер CLOCK: BMAL1 [44].
Еще одним фактором, влияющим на функцию циркадного ритма, являются гены сиртуинов (SIRT). Гены семейства SIRT представляют собой NAD + -зависимые ферменты деацетилазы класса III, которые влияют на многие клеточные функции, включая метаболизм человека, старение, рак и клеточное старение.Существует 7 вариантов семейства SIRT (SIRT 1–7). Регуляция экспрессии генов — самый важный механизм, на который влияет семейство SIRT. SIRT1, SIRT6 и SIRT7 преимущественно локализуются в ядре клетки, SIRT2 — в цитоплазме, а SIRT3, SIRT4 и SIRT5 — в митохондриях. SIRT1 играет важную роль в регуляции метаболических процессов, таких как чувствительность к инсулину, метаболизм липидов и глюконеогенез, а также продолжительность жизни человека [45]. SIRT1 модулирует активность CLOCK: BMAL1 и участвует в циркадном цикле.Гены SIRT 3-5 регулируют внутриклеточные пути, такие как окисление жирных кислот, кетогенез, цикл мочевины и окислительное фосфорилирование [33].
Внутриклеточное окислительно-восстановительное (окислительно-восстановительное) состояние — важный фактор, регулирующий гены часов в периферических тканях. Белки CLOCK могут эффективно связываться только в присутствии восстановленных последовательностей NADH и NADPH, BMAL1 и E-box. Напротив, формы никотинамидадениндинуклеотидоксидазы (NAD + и NADP +) ингибируют связывание комплекса CLOCK: BMAL1 с ДНК [32].Таким образом, окислительно-восстановительный статус NAD / NADH клетки может приводить к изменениям циркадной фазы, влияя на транскрипционную активность генов BMAL1: CLOCK [46].
Внутриклеточные уровни НАД +, а также повышенные уровни АМФ (аденозинмонофосфата) являются индикаторами низкой энергии. Когда внутриклеточные уровни АТФ снижаются, AMPK (AMP-активирующая протеинкиназа) функционирует как пищевой сенсор и активирует внутриклеточные пути снабжения энергией. Таким образом, соотношение АМФ / АТФ также может быть связующим звеном между циркадным ритмом и энергетическим метаболизмом [47].
Влияние циркадного ритма на энергетический баланс
Энергия, производимая и сохраняемая в процессе метаболизма, используется для поддержания метаболической активности, такой как основной уровень метаболизма, физическая активность и тепловой эффект пищевых продуктов [48].
Фрэнсис Дж. Бенедикт впервые описал циркадные изменения энергетического метаболизма в 1915 году [49]. Кроме того, Haugen et al. [50] обнаружили, что скорость метаболизма в покое в полдень была на 6% выше, чем в утренние часы. Одним из наиболее важных факторов, влияющих на скорость основного обмена, является режим сна.Сон и циркадный ритм являются основными компонентами регуляции энергетического обмена [48, 51]. Есть 2 фазы сна: быстрое движение глаз (REM) и не-REM [52]. Активность симпатической нервной системы и сновидения усиливаются в период REM. Температура тела, частота сердечных сокращений, частота дыхания и артериальное давление повышаются во время фазы быстрого сна. Нарушения в периоде быстрого сна увеличиваются из-за активности симпатической нервной системы [53]. Из-за повышения температуры тела и расхода энергии в головном мозге (потребление энергии мозгом в этот период составляет примерно 25%), скорость метаболизма во время сна достигает наивысшей точки в фазе быстрого сна [54].Следовательно, ухудшение цикла сна из-за позднего сна, смены часовых поясов, сменной работы и т. Д. Может привести к снижению основной скорости метаболизма из-за изменения времени фазы быстрого сна.
Помимо основной скорости метаболизма / метаболизма в покое, физическая активность является важным компонентом общих затрат энергии [55]. Однако исследования подчеркивают различные метаболические эффекты типа, продолжительности и продолжительности упражнений. Например, обзорное исследование подчеркивает, что максимальная производительность при краткосрочных анаэробных упражнениях достигается ближе к вечеру, обычно в полдень [56], что также соответствует максимальной температуре тела [56, 57].Температура тела рассматривается как «основная переменная» циркадного ритма и используется как маркер циркадного ритма [58]. Разница в температуре тела между утренними и вечерними часами составляет 0,9 ° C [59]. Температура тела влияет на мышечную активность [60]. Эта разница между утренними и вечерними часами может повлиять на выполнение упражнений и, косвенно, на скорость основного обмена.
Время приема пищи влияет на физическую работоспособность. Упражнения до и после еды по-разному влияют на окисление жирных кислот и метаболизм аппетита [61–63].Однако до сих пор неясно, когда (тренировка до еды / после еды) является наиболее эффективным временем для похудения. Подход к снижению массы тела и облегчению сжигания жира заключается в выполнении постпрандиальных аэробных упражнений после ночного голодания [64]. Упражнения во время голодания вызывают использование отложений гликогена для увеличения окисления жиров и снижает инсулин в плазме за счет увеличения уровней адреналина и норадреналина в плазме, вызывая липолиз. Напротив, некоторые исследования были посвящены положительному влиянию постпрандиальных упражнений на контроль массы тела по сравнению с упражнениями во время голодания из-за их положительного влияния на аппетит и метаболизм в покое [65].Исследование показало, что 36-минутные умеренные упражнения на беговой дорожке значительно увеличили скорость метаболизма в состоянии покоя через 40 минут после завтрака в средиземноморском стиле в первые 24 часа [66]. Во многих исследованиях общее потребление энергии, режимы питания, физические характеристики, продолжительность и тяжесть упражнений различаются, что приводит к различной интерпретации результатов.
Еще одна составляющая общих затрат энергии — тепловое воздействие питательных веществ. На тепловой эффект продуктов влияет макроэлементный состав рациона.Липиды обладают наименьшим (0–3%), а белки — наибольшим (20–30%) термогенным эффектом [67]. Кроме того, время приема пищи является важным фактором, влияющим на тепловой эффект продуктов. Термогенез, вызванный диетой, выше в утренние часы по сравнению с вечерними и ночными часами [68]. Аналогичным образом в исследовании Morris et al. [69], тепловой эффект от еды утром был на 44% выше, чем вечером. Снижение теплового эффекта пищи с утра до вечера может быть в первую очередь связано с влиянием эндокринной циркадной системы на физиологию желудочно-кишечного тракта.Периодичность кишечника в утренние часы более эффективна, чем в вечерние. Два исследования на здоровых взрослых показали, что скорость опорожнения желудка утром (8.00) была выше, чем вечером (20.00–11.00) [70]. Кроме того, нарушения циркадного ритма, такие как посменная работа и нарушение биоритмов, вызывают расстройства желудочно-кишечной системы, такие как боли в животе, вздутие живота, диарея или запор. Эти наблюдения показывают функциональную корреляцию между суточными ритмами и физиологией желудочно-кишечного тракта.Hoogerwerf et al. [71] показали, что экспрессия белков PER2 и BMAL1 была связана с циркадным ритмом в области миентерального сплетения, который играет важную роль в координации эпителиальных клеток толстой кишки и моторики толстой кишки.
Влияние приема пищи и физической активности на циркадные ритмы
Здесь подчеркивается влияние циркадного ритма на компоненты расхода энергии и его роль в энергетическом балансе. Однако некоторые исследования показали, что некоторые факторы окружающей среды (т.е., пища, состав пищи, время кормления и упражнения) могут быть эффективны для периферических часов, которые существуют в нескольких частях тела, таких как печень, поджелудочная железа или сердце [72–75]. Периферийные часы играют неотъемлемую и уникальную роль в каждой из своих тканей, управляя циркадной экспрессией определенных генов, участвующих в различных физиологических функциях. Существование всех этих часов, работающих вместе и синхронизируемых центральными часами, со многими гормонами и физиологическими или экологическими переменными, изменяющимися в течение дня, делает это двунаправленное взаимодействие в циркадной системе довольно сложным [4].
Питание — один из внешних синхронизаторов наших периферийных часов. Основная роль циркадных часов — вовлекать организм в сигналы окружающей среды; это позволяет организмам прогнозировать наличие пищи. Ограничение доступа к пище определенным временем дня оказывает глубокое влияние на поведение и физиологию организмов [72]. Damiola et al. [76] показали, что временное ограничение питания в условиях свет-темнота или темнота-темнота может изменить фазу экспрессии циркадных генов в типах периферических клеток на срок до 12 часов, не затрагивая при этом фазу экспрессии циклических генов в SCN.
Время кормления оказывает значительное влияние на репертуар, фазу и амплитуду экспрессии ритмических генов. В исследовании было показано, что и временной характер потребления пищи, и циркадные часы влияют на транскрипцию печеночных генов у мышей дикого типа [73].
Состав рациона — еще один важный фактор, влияющий на циркадные часы. Kohsaka et al. [74] показали, что диета с высоким содержанием жиров у мышей приводила к изменениям периода ритма двигательной активности и изменениям в экспрессии и цикличности генов канонических циркадных часов, ядерных рецепторов, которые регулируют факторы транскрипции часов, и участвующих в них генов, контролируемых часами. в использовании топлива в гипоталамусе, печени и жировой ткани.
Одним из примечательных нефотических сигналов для регулирования периферийных часов являются упражнения. Предполагается, что физическая активность или упражнения вызывают некоторые физиологические изменения, такие как изменения температуры тела и гормонального статуса, которые, как известно, влияют на периферические часы через активацию симпатической нервной системы и высвобождение глюкокортикоидов [75]. Повышение температуры тела может действовать как сигнал для циркадного водителя ритма у млекопитающих [77]. Кроме того, упражнения способствуют выработке и высвобождению мелатонина.Благоприятный эффект 4-недельного лечения мелатонином для модуляции циркадных компонентов цикла сна и бодрствования обычно приводит к улучшению качества сна [78].
Напротив, молекулярные циркадные часы в периферических тканях могут реагировать на время выполнения упражнений, предполагая, что физическая активность обеспечивает важную временную информацию для синхронизации циркадных часов по всему телу. Хотя точная продолжительность и интенсивность упражнений, необходимых для изменения циркадной ритмики, не были определены, одно исследование на мышах показало, что упражнения на выносливость низкой интенсивности, поддерживаемые курсом по 2 часа в день в течение 4 недель, были достаточными, чтобы захватить циркадные часы и изменить циркадную ритмичность [79].
Обычно, когда периферийные часы десинхронизируются с центральными часами, это приводит к сбою хронометража [80]. Это физиологическое изменение связано с различными заболеваниями, такими как рак, сердечно-сосудистые заболевания, депрессия, ожирение и метаболический синдром [72]. Например, при лечении ожирения основным подходом к диетическому лечению является ограничение потребления энергии [81]. Как правило, факторы, которые напрямую влияют на биологические ритмы, такие как время приема пищи и время сна, обычно не исследуются при планировании диеты.Циркадные часы играют важную роль в энергетическом гомеостазе и метаболических процессах. Следовательно, оценка факторов (сменная работа, нерегулярный сон, бессонница и т. Д.), Которые могут привести к нарушениям циркадного ритма у лиц с метаболическими заболеваниями, такими как ожирение, и планирование тренировок и времени приема пищи в соответствии с нормальными биологическими ритмами ( например, утренний выбор белковой пищи для увеличения термогенеза) может повысить эффективность лечения.
В последние годы для поддержания метаболического здоровья были разработаны методы лечения, зависимые от циркадного ритма, диетические вмешательства и упражнения, которые получили название «хронофармакология», «хроническое питание» и «хроноупражнения» соответственно.Хронофармакология изучает правильное время введения доз лекарственного средства для повышения эффективности, абсорбции и / или эффективности лекарственного средства [65]. Например, фермент HMG-COA, ограничивающий уровень холестерина, демонстрирует циркадный ритм у людей. Этот фермент достигает пика ночью, поэтому рекомендуется принимать на ночь препараты, снижающие уровень холестерина, такие как статины, чтобы максимизировать их эффективность [19]. Хроническое питание — это подход к определению оптимального усвоения питательных веществ для поддержания здоровья и регулирования циркадного ритма [82].Например, кофеин, нобилетин (флавоноид, содержащийся в цитрусовых) и ресвератрол в пищевых продуктах могут вызывать изменения циркадного ритма на молекулярном или поведенческом уровнях [83]. Chronoexercise в первую очередь исследует влияние продолжительности упражнений на поддержание здоровья и спортивные результаты, быстрые изменения в системе внутренних часов или повторное регулирование циркадных часов [84].
В результате циркадный ритм имеет двунаправленное взаимодействие почти со всеми метаболическими процессами и является основным фактором, влияющим на цикл сна и бодрствования.Следовательно, изучение и использование режима сна, информации о качестве и составление руководств по лечению с использованием циркадного ритма может повысить эффективность лечения заболевания. По этой причине могут быть разработаны новые подходы, перспективы и стратегии лечения метаболического баланса.
Благодарности
Нет.
Заявление об этике
Эта статья не содержит исследований с участием людей или животных, проведенных кем-либо из авторов.
Заявление о раскрытии информации
Авторы заявляют, что у них нет конфликта интересов.
Источник финансирования
Нет.
Вклад авторов
Ю.С. провели обзор литературы и N.A.T. организовал и подготовил рукопись. Все авторы читали и одобрили окончательный вариант рукописи.
Список литературы
- Акынджи Э., Орхан ФО.Sirkadiyen ritim uyku bozuklukları. Псикиятр Гунджел Якласимлар. 2016; 8 (2): 178–89.
- Özbayer C, Değirmenci İ. Sirkadiyen saat, hücre döngüsü ve kanser. Дикле Тип Дергиси; 2011. с. 38.
- Гумз М.Л., редактор.Циркадные часы: роль в здоровье и болезнях. Springer; 2016 г.
- Ричардс Дж., Гумз МЛ. Успехи в понимании периферических циркадных часов. FASEB J. 2012 сентябрь; 26 (9): 3602–13.
- Güldür T, Otlu HG.Циркадный ритм у млекопитающих: время есть и время спать. Biol Rhythm Res. 2017; 48 (2): 243–61.
- Аллен Р.П. В рецензируемой статье: увлечение мелатонином свободных циркадных ритмов у слепых людей. Sleep Med. Март 2001 г., 2 (2): 167–8.
- Санти Н., Лазар А.С., МакКейб П.Дж., Ло Дж.К., Грёгер Дж. А., Дейк Д.Дж.Половые различия в суточной регуляции сна и познания в бодрствовании у людей. Труды Национальной академии наук. 2016: 201521637.
- Ферри Дж. Э., Кумари М., Сало П., Сингх-Ману А., Кивимяки М. Эпидемиология сна — быстро развивающаяся область. Издательство Оксфордского университета; 2011 г.
- Кронхольм Э., Партонен Т., Лаатикайнен Т., Пелтонен М., Хярма М., Хублин С. и др. Тенденции изменения продолжительности сна и симптомов, связанных с бессонницей, в Финляндии с 1972 по 2005 год: сравнительный обзор и повторный анализ выборок финского населения. J Sleep Res. Март 2008 г., 17 (1): 54–62.
- Чжу Л., Зи ПК. Расстройства циркадного ритма сна. Neurol Clin. 2012 ноябрь; 30 (4): 1167–91.
- Авраам Ф.Обзор функциональных причин бесплодия у коров. JFIV Reprod Med Genet. 2017; 5 (2): 203.
- Schernhammer ES, Laden F, Speizer FE, Willett WC, Hunter DJ, Kawachi I, et al. Работа в ночную смену и риск колоректального рака в исследовании состояния здоровья медсестер. J Natl Cancer Inst.2003 июн; 95 (11): 825–8.
- Гейл Дж. Э., Кокс Х. И., Цянь Дж., Блок Г. Д., Колвелл К. С., Матвеенко А. В.. Нарушение циркадных ритмов ускоряет развитие диабета из-за потери и дисфункции бета-клеток поджелудочной железы. J Biol Rhythms. 2011 Октябрь; 26 (5): 423–33.
- Мартино Т.А., Аудит Г.Ю., Герценберг А.М., Тата Н., Колетар М.М., Кабир Г.М. и др.Дезорганизация циркадного ритма вызывает у хомяков серьезные сердечно-сосудистые и почечные заболевания. Am J Physiol Regul Integr Comp Physiol. 2008 Май; 294 (5): R1675–83.
- Бакли П. Сон и нарушение циркадного ритма при шизофрении. Ежегодник психиатрии и прикладного психического здоровья; 2013.С. 411–2.
- Чхве СС, Ху Джи, Хван Ай Джей, Ким Джи, Ким Джи Би. Ремоделирование жировой ткани: его роль в энергетическом обмене и метаболических нарушениях. Фронт-эндокринол (Лозанна). 2016 Апрель; 7:30.
- On M, Dalğin D, Cenesiz M, Cenesiz S.Лептин ве адипонектинин enerji ve egzersiz ilişkisi. 2017 г.
- Цуджино Н., Сакураи Т. [Циркадный ритм лептина, орексина и грелина]. Нихон Риншо. 2012 июль; 70 (7): 1121–5.
- Сато Т, Ида Т, Кодзима М.Роль биологических ритмов в выполнении физических нагрузок. J Phys Fit Sports Med. 2017; 6 (3): 125–34.
- МакГиннис Г.Р., Молодой ME. Циркадная регуляция метаболического гомеостаза: причины и последствия. Nat Sci Sleep. 2016 Май; 8: 163–80.
- İbrahim Erdemir ET.Kortizol Sirkadiyen Ritmini Etkileyen Bazı Fiziksel ве Fizyolojik Parametrelerin Karşılaştırılması. Balıkesir Üniversitesi Sosyal Bilimler Enstitüsü Dergisi. 2008; 11: 1–10.
- Christiansen JJ, Djurhuus CB, Gravholt CH, Iversen P, Christiansen JS, Schmitz O, et al. Влияние кортизола на метаболизм углеводов, липидов и белков: исследования острого синдрома отмены кортизола при надпочечниковой недостаточности.J Clin Endocrinol Metab. 2007 сентябрь; 92 (9): 3553–9.
- Bolli GB, De Feo P, De Cosmo S, Perriello G, Ventura MM, Calcinaro F и др. Демонстрация феномена рассвета на нормальных людях-добровольцах. Диабет. 1984 декабрь; 33 (12): 1150–3.
- Рыбицка М., Крысяк Р., Окопень Б.Феномен рассвета и эффект Сомоджи — два явления утренней гипергликемии. Эндокринол Pol. 2011. 62 (3): 276–84.
- Özçelik F, Erdem M, Bolu A, Gülsün M. Melatonin: genel özellikleri ve psikiyatrik bozukluklardaki rolü. Псикиятр Гунджел Якласимлар. 2013; 5 (2).
- Pan X, Zhang Y, Wang L, Hussain MM.Суточная регуляция MTP и триглицеридов плазмы с помощью CLOCK опосредуется SHP. Cell Metab. 2010 август; 12 (2): 174–86.
- Пан X, Хуссейн MM. Суточная регуляция микросомальных белков-переносчиков триглицеридов и уровней липидов в плазме. J Biol Chem. 2007 август; 282 (34): 24707–19.
- Пан X, Хуссейн MM.Часы важны для питания и суточной регуляции абсорбции макроэлементов у мышей. J Lipid Res. 2009 Сен; 50 (9): 1800–13.
- Бейли С.М., Удох США, Янг МЭ. Циркадная регуляция обмена веществ. J Endocrinol. 2014 Август; 222 (2): R75–96.
- Стивенсон Р.Циркадные ритмы и нарушения дыхания во сне. Sleep Med. 2007 сентябрь; 8 (6): 681–7.
- Muller JE, Stone PH, Turi ZG, Rutherford JD, Cheisler CA, Parker C, et al. Циркадные вариации частоты возникновения острого инфаркта миокарда. N Engl J Med. 1985 ноя; 313 (21): 1315–22.
- Фрой О. Метаболизм и циркадные ритмы — значение для ожирения. Endocr Rev.2010, февраль; 31 (1): 1–24.
- Крамер А., Мерроу М., редакторы.Циркадные часы. Springer; 2013.
- Браун С.А., Аззи А. Периферийные циркадные осцилляторы у млекопитающих; Циркадные часы. Springer; 2013. С. 45–66.
- Дэвидсон А.Дж., Лондон Б., Блок Г.Д., Менакер М.Сердечно-сосудистые ткани содержат независимые циркадные часы. Clin Exp Hypertens. 2005, февраль-апрель; 27 (2-3): 307–11.
- la Fleur SE, Kalsbeek A, Wortel J, Fekkes ML, Buijs RM. Суточный ритм толерантности к глюкозе: роль супрахиазматического ядра. Диабет. 2001 июн; 50 (6): 1237–43.
- Харада Н., Инагаки Н. Роль часовых генов в секреции инсулина. J. Исследование диабета. 2016 ноя; 7 (6): 822–3.
- Mıcılı S, Özoul C.Диябетте Кёк Хюкрелер. Dokuz Eylül Üniversitesi Tıp Fakültesi Dergisi. 2007; 21: 109–17.
- Sözlü S, anlier N. Sirkadiyen Ritim, Sağlık ve Beslenme İlişkisi. Turkiye Klinikleri Journal of Health Sciences. 2017; 2: 100–9.
- Кесслер К, Пивоварова О, Пфайффер АФ.[Циркадные часы и энергетический обмен: значение для здоровья]. Dtsch Med Wochenschr. 2014 Апрель; 139 (14): 684–6.
- Санкар Г., Бруннер М. Циркадные часы и энергетический метаболизм. Cell Mol Life Sci. Июль 2014 г .; 71 (14): 2667–80.
- Масри С.Сиртуин-зависимый контроль часов: новые достижения в метаболизме, старении и раке. Curr Opin Clin Nutr Metab Care. 2015 ноя; 18 (6): 521–7.
- Ян Х, Даунс М., Ю РТ, Буккаут А.Л., Хе В., Страуме М. и др. Экспрессия ядерных рецепторов связывает циркадные часы с метаболизмом. Клетка. 2006 август; 126 (4): 801–10.
- Фрой О., Мискин Р. Взаимосвязь кормления, циркадных ритмов и старения. Prog Neurobiol. 2007 июнь; 82 (3): 142–50.
- Байрам А., Мехри И.Sirtuin Genleri ve İşlevleri. Firat Tip Derg. 2013; 18: 136–40.
- Раттер Дж., Рейк М., Ву Л.С., Макнайт С.Л. Регулирование часов и связывания ДНК NPAS2 окислительно-восстановительным состоянием кофакторов NAD. Наука. Июль 2001 г., 293 (5529): 510–4.
- Ум Дж. Х., Ян С., Ямадзаки С., Канг Х., Виоллет Б., Форец М. и др.Активация 5′-AMP-активированной киназы лекарственным средством от диабета метформином вызывает зависимую от казеинкиназы ипсилон (CKIepsilon) деградацию часового белка mPer2. J Biol Chem. Июль 2007 г., 282 (29): 20794–8.
- Kumar Jha P, Challet E, Kalsbeek A. Циркадные ритмы метаболизма глюкозы и липидов у ночных и дневных млекопитающих.Mol Cell Endocrinol. 2015 декабрь; 418 (Pt 1): 74–88.
- Бенедикт Ф.Г. Факторы, влияющие на основной обмен. Proc Natl Acad Sci USA. 1915 Февраль; 1 (2): 105–9.
- Haugen HA, Melanson EL, Tran ZV, Kearney JT, Hill JO.Вариабельность измеренной скорости метаболизма в состоянии покоя. Am J Clin Nutr. 2003 декабрь; 78 (6): 1141–5.
- Лапоски А.Д., Басс Дж., Кохсака А., Турек Ф.В. Сон и циркадные ритмы: ключевые компоненты регуляции энергетического обмена. FEBS Lett. Январь 2008 г., 582 (1): 142–51.
- Алгин Д.И., Акдаг Г., Эрдинч О.О.Калители уйку ве уйку бозуклуклари. Османгази медицинский журнал. 2016; 38.
- Шахин Л., Ашиоглу М., Ташкин Э. Уйку ве уйкунун дюзенленмеси. Sağlık Bilimleri Dergisi. 2013; 22: 93–8.
- Boscolo RA, Esteves AM, de Santana MG, Viana VAR, Grassmann V, Tufik S, de Mello MT.Есть ли связь между составом тела, основной скоростью метаболизма и сном у пожилых пациентов с синдромом обструктивного апноэ во сне и без него? Наука о сне. 2013. 6 (4): 129–134.
- Knutson KL, Spiegel K, Penev P, Van Cauter E. Метаболические последствия лишения сна.Sleep Med Rev.2007 июн; 11 (3): 163–78.
- Chtourou H, Souissi N. Эффект тренировки в определенное время суток: обзор. J Strength Cond Res. 2012 Июль; 26 (7): 1984–2005.
- Суиси Н., Готье А., Сесбое Б., Лару Дж., Давенн Д.Циркадные ритмы в двух типах упражнений для ног анаэробного цикла: сила-скорость и 30-секундный тест Вингейта. Int J Sports Med. 2004 Янв; 25 (1): 14–9.
- Kinişler A. Anaerobik Performansta sirkadiyen değişimlerin incelenmesi. Spor Bilimleri Dergisi. 2005. 16: 174–84.
- Кройчи К.Как регулируется циркадный ритм основной температуры тела? Clin Auton Res. 2002 июн; 12 (3): 147–149.
- Райт К.П. младший, Халл Дж. Т., Чейслер, Калифорния. Взаимосвязь между бдительностью, работоспособностью и температурой тела у людей. Am J Physiol Regul Integr Comp Physiol. 2002 декабрь; 283 (6): R1370–7.
- Беннард П., Дусе Э. Острые эффекты времени выполнения упражнений и гликемического индекса завтрака на окисление жиров, вызванное физической нагрузкой. Appl Physiol Nutr Metab. 2006 Октябрь; 31 (5): 502–11.
- Катсанос К.С., Моффатт Р.Дж.Острые эффекты упражнений перед едой по сравнению с упражнениями после еды на постпрандиальную гипертриглицеридемию. Clin J Sport Med. 2004, январь; 14 (1): 33–9.
- Фарах Н.М., Гилл Дж. М.. Влияние упражнений до или после приема пищи на баланс жиров и постпрандиальный метаболизм у мужчин с избыточным весом. Br J Nutr.2013 июн; 109 (12): 2297–307.
- Канг Дж., Рейнс Э., Розенберг Дж., Ратамесс Н., Наклерио Ф., Файгенбаум А. Метаболические реакции во время постпрандиальных упражнений. Res Sports Med. 2013. 21 (3): 240–52.
- Шибата С., Тахара Ю.Циркадный ритм и упражнения. J Phys Fit Sports Med. 2014; 3 (1): 65–72.
- Паоли А., Марколин Дж., Зонин Ф., Нери М., Сивьери А., Пачелли QF. Делать упражнения натощак или кормить для ускорения похудания? Влияние приема пищи на соотношение дыхательных путей и избыточное потребление кислорода после тренировки после тренировки на выносливость.Int J Sport Nutr Exerc Exerc Metab. 2011 Февраль; 21 (1): 48–54.
- Таппи Л. Термический эффект питания и деятельность симпатической нервной системы человека. Reprod Nutr Dev. 1996. 36 (4): 391–7.
- Romon M, Edme JL, Boulenguez C, Lescroart JL, Frimat P.Циркадные вариации термогенеза, вызванного диетой. Am J Clin Nutr. 1993, апрель; 57 (4): 476–80.
- Моррис С.Дж., Гарсия Д.И., Майерс С., Ян Дж., Триенекенс Н., Шеер Ф.А. Циркадная система человека играет доминирующую роль в возникновении утренних и вечерних различий в термогенезе, вызванном диетой.Ожирение (Серебряная весна). 2015 Октябрь; 23 (10): 2053–8.
- Grammaticos PC, Doumas A, Koliakos G. Половина утреннего и ночного опорожнения желудка различалась более чем на 220% у двух молодых здоровых взрослых людей. Ад J Nucl Med. 2015, январь-апрель; 18 (1): 60–2.
- Hoogerwerf WA.Роль часовых генов в моторике желудочно-кишечного тракта. Am J Physiol Gastrointest Liver Physiol. 2010 сентябрь; 299 (3): G549–55.
- Гараулет М., Гомес-Абеллан П. Время приема пищи и ожирение: новая ассоциация. Physiol Behav. 2014 июль; 134: 44–50.
- Фоллмерс К., Гилл С., Ди Таккио Л., Пуливарти С.Р., Ле HD, Панда С.Время кормления и внутренние ритмы привода циркадных часов в экспрессии генов печени. Proc Natl Acad Sci U S. A. 2009, 15 декабря; 106 (50): 21453–8.
- Kohsaka A, Laposky AD, Ramsey KM, Estrada C, Joshu C., Kobayashi Y, et al. Диета с высоким содержанием жиров нарушает поведенческие и молекулярные циркадные ритмы у мышей.Cell Metab. 2007 ноябрь; 6 (5): 414–21.
- Тахара Ю., Аояма С., Шибата С. Циркадные часы млекопитающих и их увлечение стрессом и физическими упражнениями. J Physiol Sci. 2017 Янв; 67 (1): 1–10.
- Дамиола Ф., Ле Минь Н., Прейтнер Н., Корнманн Б., Флери-Олела Ф., Шиблер У.Ограниченное питание отделяет циркадные осцилляторы в периферических тканях от центрального водителя ритма в супрахиазматическом ядре. Genes Dev. 2000 декабрь; 14 (23): 2950–61.
- Brown SA, Zumbrunn G, Fleury-Olela F, Preitner N, Schibler U. Ритмы температуры тела млекопитающих могут поддерживать периферические циркадные часы.Curr Biol. 2002 Сен; 12 (18): 1574–83.
- Леонардо-Мендонса Р.К., Мартинес-Николас А., де Тереза Гальван С., Оканья-Вильхельми Дж., Русанова И., Герра-Эрнандес Е. и др. Преимущества четырехнедельного лечения мелатонином на циркадные ритмы у спортсменов, тренирующихся с отягощениями. Chronobiol Int.2015; 32 (8): 1125–34.
- Wolff G, Esser KA. Запланированная фаза упражнений сдвигает циркадные часы в скелетных мышцах. Медико-спортивные упражнения. 2012 сентябрь; 44 (9): 1663–70.
- Гараулет М., Ордовас Я. М., Мадрид, Я.Хронобиология, этиология и патофизиология ожирения. Int J Obes. 2010 декабрь; 34 (12): 1667–83.
- Фрой О. Циркадные ритмы и ожирение у млекопитающих. ISRN Obes. 2012 18 ноября; 2012: 437198.
- Тахара Ю., Шибата С.Хронобиология и питание. Неврология. 2013 декабрь; 253: 78–88.
- Ойке Х. Модуляция суточных часов питательными веществами и пищевыми факторами. Biosci Biotechnol Biochem. 2017 Май; 81 (5): 863–70.
- Накао Р.Циркадные часы скелетных мышц: текущие исследования. Хронофизиология и терапия. 2017; 7: 47–57.
Автор Контакты
Nilüfer Acar Tek
Факультет медицинских наук, Департамент питания и диетологии
Университет Гази, район Эмниет, улица Муаммера Яшара Бостанджи
Номер 16, TR – 06560 Анкара (Турция)
Электронная почта acarnil @ hotmail.com
Подробности статьи / публикации
Предварительный просмотр первой страницы
Получено: 26 июня 2018 г.
Принято: 30 марта 2019 г.
Опубликовано онлайн: 23 апреля 2019 г.
Дата выпуска: июнь 2019 г.
Количество страниц для печати: 9
Количество рисунков: 1
Количество столов: 1
ISSN: 0250-6807 (печать)
eISSN: 1421-9697 (онлайн)
Для дополнительной информации: https: // www.karger.com/ANM
Авторские права / Дозировка препарата / Заявление об ограничении ответственности
Авторские права: Все права защищены. Никакая часть данной публикации не может быть переведена на другие языки, воспроизведена или использована в любой форме или любыми средствами, электронными или механическими, включая фотокопирование, запись, микрокопирование, или с помощью какой-либо системы хранения и поиска информации, без письменного разрешения издателя. .
Дозировка лекарств: авторы и издатель приложили все усилия, чтобы гарантировать, что выбор и дозировка лекарств, указанные в этом тексте, соответствуют текущим рекомендациям и практике на момент публикации.Однако ввиду продолжающихся исследований, изменений в правительственных постановлениях и постоянного потока информации, касающейся лекарственной терапии и реакций на них, читателю настоятельно рекомендуется проверять листок-вкладыш для каждого препарата на предмет любых изменений показаний и дозировки, а также дополнительных предупреждений. и меры предосторожности. Это особенно важно, когда рекомендованным агентом является новое и / или редко применяемое лекарство.
Отказ от ответственности: утверждения, мнения и данные, содержащиеся в этой публикации, принадлежат исключительно отдельным авторам и соавторам, а не издателям и редакторам.Появление в публикации рекламы и / или ссылок на продукты не является гарантией, одобрением или одобрением рекламируемых продуктов или услуг или их эффективности, качества или безопасности. Издатель и редактор (-ы) не несут ответственности за любой ущерб, причиненный людям или имуществу в результате любых идей, методов, инструкций или продуктов, упомянутых в контенте или рекламе.
метаболических процессов в раковых клетках, изученных с помощью фармако-метаболомического подхода
Аспирант Цзяцзюнь Ду (Jiajun Du) делает в лаборатории изображение раковых клеток меланомы с помощью комбинационного рассеяния света.[Калтех]Одна из самых больших проблем при разработке методов лечения рака заключается в том, что не существует единственного вида рака. В результате исследователи пытаются узнать больше о том, как функционируют определенные виды рака, чтобы создать эффективные методы лечения этих типов рака.
Ученые из Калифорнийского технологического института и сотрудники опубликовали статью «Рамановская субклеточная фармакомаболомика для метастатических клеток меланомы» в журнале Nature Communications , в которой показано, что разработанная ими структура с использованием специального типа микроскопии позволяет им исследовать метаболические процессы внутри рака. клетки.
«Нас интересует вопрос, почему все раковые клетки, на которые мы смотрим, очень различаются по поведению. Поскольку некоторые клетки больше полагаются на некоторые метаболические пути, они более восприимчивы к нарушению этих путей », — говорит Лу Вэй, доктор философии, доцент кафедры химии Калифорнийского технологического института, чья команда работала с исследователями из Института системной биологии в Сиэтле и США. Калифорнийский университет в Лос-Анджелесе (UCLA).
Ученые использовали рамановскую спектроскопию в сочетании с ее усовершенствованной версией, микроскопией вынужденного комбинационного рассеяния (SRS).Рамановская спектроскопия использует естественные колебания, возникающие в связях между атомами, составляющими молекулу.
В этом методе молекула облучается лазерным светом. Когда фотоны лазерного света отражаются от молекулы, они набирают или теряют энергию в результате взаимодействия с колебаниями в связях молекулы. Поскольку каждый вид связи в молекуле воздействует на фотоны уникальным и предсказуемым образом, о структуре молекулы можно судить по тому, как фотоны «выглядят» после того, как отскакивают от нее.
Картируя распределение целевых химических связей, SRS микроскопия затем дает изображения этих молекулярных структур.
Используя эти комбинированные методы, Вэй и ее коллеги исследовали метаболиты, присутствующие в пяти клеточных линиях меланомы, обычно используемых в исследованиях. По словам Вэй, клетки меланомы были выбраны потому, что они обладают широким спектром метаболических характеристик, которые можно изучать.
«Неинвазивное зондирование метаболитов в отдельных живых клетках очень желательно, но сложно.Здесь мы используем спектромикроскопию комбинационного рассеяния для пространственного картирования метаболитов в отдельных клетках с конкретной целью выявления лекарственной метаболической восприимчивости из ряда клеточных линий меланомы, полученных от пациентов. Каждая клеточная линия представляет собой свой характерный уровень де-дифференцировки раковых клеток », — пишут исследователи.
«Во-первых, с помощью спектроскопии комбинационного рассеяния, затем микроскопии стимулированного комбинационного рассеяния (SRS) и анализа транскриптомики, мы идентифицируем путь синтеза жирных кислот как лекарственную чувствительность для дифференцированных меланоцитарных клеток.Затем мы используем гиперспектральную-SRS визуализацию внутриклеточных липидных капель, чтобы идентифицировать ранее неизвестную восприимчивость к липидной мононенасыщенности в недифференцированных мезенхимальных клетках с врожденной устойчивостью к ингибированию BRAF. Введение лекарств в эту мишень приводит к клеточному апоптозу, сопровождающемуся образованием разделенных фазами внутриклеточных мембранных доменов ».
«Интеграция субклеточной рамановской спектромикроскопии с липидомикой и транскриптомикой предполагает возможные механизмы регуляции липидов, лежащие в основе этого фармакологического лечения.Наш метод должен обеспечивать общий подход к исследованиям метаболомики отдельных клеток с пространственным разрешением ».
Изучая метаболиты клеток, исследователи могут начать делать выводы о том, как работает их метаболизм и как на них могут воздействовать лекарства. Вэй говорит, что команда обнаружила несколько новых метаболических факторов восприимчивости раковых клеток, включая синтез жирных кислот и мононенасыщенность, но добавляет, что прямо сейчас основная цель исследования — заниматься фундаментальной наукой.
«Мы внедрили концепцию рамановской спектроскопии в системную биологию», — говорит она.«И мы используем субклеточную информацию, которую мы собрали вместе с ней, чтобы направлять наше исследование фармакометаболомики — изучение того, как метаболизм влияет на лекарства».
Джеймс Р. Хит, доктор философии из Института системной биологии в Сиэтле и соавтор статьи, говорит, что эта новая технология позволяет исследователям получить более подробный взгляд на раковые клетки, чем когда-либо прежде.
«Методы химической визуализации, разработанные в лаборатории Лу, позволили нам определить метаболическую восприимчивость к лекарственным препаратам на некоторых очень агрессивных моделях рака.Эти метаболические слабости были бы упущены при любом другом аналитическом подходе », — отмечает Хит.
PLOS ONE: метаболические процессы
Ф. М. Я. Нур-Назратул, М. Р. М. Ракиб, М. З. Зайлан, Х. Яакуб
Джили Чжан, Пэн Ван, […], Ся Ву
Сюнь Лю, Киёси Ямагути, […], Цунео Икеноуэ
Ниннин Чжан, Чжанвэй Тэн, […], Тэнъюн Гао
Анастасия Миненкова, Эрвин Э. В. Янсен, […], Саадет Мерсимек-Эндрюс
Аврил Й. Э. фон Хойнинген-Хюене, Табеа Дж. Шлоттауэр, […], Рольф Даниэль
Юнджунг Чой, Бэксун Ким, […], Хе-Ин Им
Конгпхоп Парунякул, Криттика Сришуксай, […], Вирасак Фунгфуанг
Жанин Хофманн, Мебрату А. Биту, […], Фиона М. Сансом
Шуюань Дэн, Бо Ван, […], Фань Чжан
Цзяцзин Ван, Лепин Цзян, Hanlong Sun
Вэньхуа Ян, Цзяньли Чжоу, Жан Дамаскен Хариндинтвали, Сяобинь Юй
Михаил Зарубин, Альберт Гангапшев, […], Елена Кравченко
Редокс-регулирование метаболических процессов — Виртуальная коллекция — Выбор статей — Редокс-биология — Журнал
Может показаться удивительным, что виртуальный выпуск Redox Biology посвящен «окислительно-восстановительной регуляции» метаболических процессов, учитывая, что уже несколько десятилетий известно, что окислительно-восстановительные реакции являются неотъемлемой частью метаболизма топлива, и локальные концентрации кофакторов, таких как НАД (Ф) Н и соотношения в окисленных и восстановленных формах таких кофакторов устанавливают регуляторную сеть, позволяющую протекать реакции, катализируемые оксидоредуктазой.
Тем не менее, исследования «окислительно-восстановительной регуляции» прошли долгий путь с тех славных дней биохимических исследований, и они изменили свое направление по крайней мере дважды; Во-первых, было обнаружено, что электроны сбиваются с пути и вносят свой вклад в образование активных форм, происходящих из молекулярного кислорода, вызывая у клеток склонность страдать от «окислительного стресса» и создавая необходимость в антиоксидантных мерах.
Второе, более недавнее изменение направления привело нас к тому месту, где мы сейчас находимся: электроны могут не просто сбиваться с пути, но и сбиваться с пути, чтобы генерировать реактивные частицы, которые, несмотря на потенциальную опасность, необходимы для основных клеточных процессов. , например, распространение.
Это последняя «окислительно-восстановительная регуляция», о которой идет речь в данном выпуске Redox Biology, подчеркивая тот факт, что от нее зависят основные метаболические процессы, включая энергетический метаболизм [1], протеолиз в нескольких вариантах, включая протеасомный и лизосомный аспекты [2, 3 ], события, зависимые от эндоплазматического ретикулума (такие как стресс ER) [4], метаболизм ксенобиотиков [5] и даже такие деликатные аспекты, как метаболическая зональность печени [6].
В этом выпуске подробно исследуются два конкретных реактивных вида, участвующих в регуляции окислительно-восстановительного потенциала; перекись водорода [7] и пероксинитрит [8] исследуются в отношении последствий их образования, деградации и реакционной способности для клеточной передачи сигналов.Более того, ишемическое прекондиционирование печени исследуется как один из примеров того, как активные формы кислорода участвуют в сигнальных событиях, влияющих на физиологию органов [9].
Поскольку для анализа биологической активности реактивных видов и окислительно-восстановительной передачи сигналов требуется соответствующая методология, две статьи сосредоточены на этих аспектах: Caenorhabditis elegans представлен в качестве подходящего модельного организма в исследованиях регулирующих и пагубных эффектов активных форм кислорода [10 ]; наконец, ряд классических анализов в исследованиях окислительного стресса, используемых для обнаружения образования реактивных частиц, обсуждается с точки зрения их постоянной полезности в исследованиях окислительно-восстановительного потенциала [11].
Этот выпуск Redox Biology основан на летней школе SFRR-Europe, которая проводилась на острове Спецес в Греции в 2016 году (19-25 сентября) как «Продвинутый курс лекций FEBS по окислительно-восстановительной регуляции метаболических процессов», организованный авторами. (Рисунок 1). Мы благодарим Федерацию европейских биохимических обществ (FEBS) и Европейское общество исследований свободных радикалов (SFRR) за поддержку этого мероприятия, а также мы благодарим Европейское сотрудничество в области науки и технологий (COST) за поддержку двух совместных мероприятий COST. , «ПРОТЕОСТАЗ» и «ЭУ-РОС».
Наконец, мы хотели бы выразить нашу благодарность авторам, рецензентам и редактору журнала Redox Biology профессору Тилману Грюну за поддержку этого начинания по сбору серии статей по захватывающей развивающейся теме исследования в области окислительно-восстановительной биологии.
Эта виртуальная коллекция была составлена доктором Ларсом-Оливером Клотцем и доктором Ники Чондроджианни
Адресная корреспонденция на:
Доктор Ларс-Оливер Клотц
Институт питания, Департамент нутригеномики, Университет Фридриха Шиллера, Йена
Дорнбургер Штрассе 29, D-07743 Йена, Германия
Тел.: + 49-3641-949750
Факс: + 49-3641-949752
Электронная почта: [электронная почта защищена]
или на:
Доктор Ники Чондроджианни
Институт биологии, медицинской химии и биотехнологии
Национальный фонд греческих исследований
48 Vassileos Constantinou Avenue, 11635, Афины, Греция
Тел .: + 30-210-7273768
Факс: + 30-210-7273677
E -mail: [адрес электронной почты защищен]
Список литературы
- Редокс-регуляция в метаболическом программировании и воспалении
Хелен Р.Griffiths, Dan Gao, Chathyan Pararasa
Redox Biology, Volume 12, август 2017 г., страницы 50-57 - Редокс-регуляция функции протеасом
Мария Лефаки, Николетта Папаевгениу, Ники Хондроджианни
Редокс-биология, Том 13, октябрь 2017 г., страницы 452-458 - Протеостаз, окислительный стресс и старение
Иоанна Коровила, Мартин Хьюго, Хосе Педро Кастро, Даниэла Вебер, Анника Хён, Тилман Грун, Тобиас Юнг
Биология редокс, Том 13, октябрь 2017 г., страницы 550-567 - Влияние высокого уровня холестерина и стресса эндоплазматического ретикулума на метаболические заболевания: обновленный мини-обзор
Эрди Созен, Несрин Картал Озер
Редокс-биология, Том 12, август 2017, страницы 456-461 - Клеточная адаптация к ксенобиотикам: взаимодействие ксеносенсоров, активных форм кислорода и факторов транскрипции FOXO
Ларс-Оливер Клотц, Хольгер Штайнбреннер
Редокс-биология, том 13, октябрь 2017 г., страницы 646-654 - Метаболическая зональность печени: пересмотр кислородного градиента
Томас Кицманн
Редокс-биология, том 11, апрель 2017 г., страницы 622-630 - Количественная биология передачи сигналов перекиси водорода
Фернандо Антунес, Паула Матос Брито
Биология окислительно-восстановительного потенциала, том 13, октябрь 2017 г., страницы 1-7 - Основы биохимии пероксинитрита и нитрования тирозина
Сильвина Бартесаги, Рафаэль Ради
Доступно онлайн 19 сентября 2017 г. - Гомеостаз АФК, ключевой фактор в прекондиционировании печени ишемией
Игнасио Прието, Мария Монсальве
Биология окислительно-восстановительного потенциала, Том 12, август 2017 г., страницы 1020-1025 - Caenorhabditis elegans как модель для понимания функции АФК в физиологии и болезнях
Антонио Миранда-Визуете, Элизабет А.Телятина
Редокс Биология, Том 11, апрель 2017 г., страницы 708-714 - Использование дубинок для традиционных анализов обнаружения активных форм кислорода и азота и их использование в сердечно-сосудистой системе
Андреас Диабер, Маттиас Олце, Себастьян Стивен, Свенья Креллер-Шен, Томас Мюнцель
Биология окислительно-восстановительного восстановления, Том 12, август 2017 г., стр. 35 -49
Этот виртуальный выпуск Redox Biology основан на летней школе SFRR-Europe, которая проводилась на Спецесе, Греция, в 2016 году (19-25 сентября) как «Продвинутый курс лекций FEBS по окислительно-восстановительной регуляции метаболических процессов», организованный авторами.