Жировой обмен веществ связан с продолжительностью жизни
Международная группа ученых продемонстрировала влияние липидов на продолжительность жизни. Ученые исследовали 669 образцов из 6 различных тканей 35 видов млекопитающих и пришли к выводу, что липидный метаболизм оказывает существенное влияние на максимальную продолжительность жизни того или иного вида. Эта работа проливает свет на молекулярные механизмы определения продолжительности жизни и механизмы, лежащие в основе процессов старения.
Результаты исследования опубликованы в журнале Nature Scientific Reports.
Максимальная продолжительность жизни — это стабильная характеристика того или иного биологического вида. Несмотря на то, что за последние пару сотен лет средняя продолжительность жизни значительно возросла, максимальная продолжительность жизни человека не изменилась и составляет приблизительно 120 лет. Если мы хотим жить дольше, необходимо понять, от чего зависит наш максимальный возраст.
Максимальная продолжительность жизни даже среди близкородственных видов млекопитающих может варьировать в десятки раз.
Профессор Сколтеха Филипп Хайтович. Фото: Sk.ru
Также ученые обнаружили, что химическое строение липидов оказывает влияние на продолжительность жизни. Например, такие структурные липиды, как сфинголипиды, входящие в состав клеточной мембраны долгоживущих видов, более насыщены водородом, чем сфинголипиды короткоживущих видов.
Екатерина Храмеева, научный сотрудник Сколтеха и ИППИ РАН, один из авторов исследования: «Нам удалось найти связь между продолжительностью жизни вида и особенностями его липидного метаболизма. Этот результат интересен сам по себе, но еще более ценными являются полученные нами данные — липидомы 35 видов млекопитающих. Такой набор данных прекрасно подходит для изучения эволюции липидома, над чем мы и работаем в данный момент. Ранее эволюцию липидома изучали лишь для 4 видов.
Результаты исследования опубликованы в журнале Nature Scientific Reports.
Источник: science.theoryandpractice.ru
Вопросы и ответы
Чтобы понять, что такое обмен веществ, как он может помочь избавиться от лишних килограммов и к чему могут привести физические упражнения на пустой желудок, прочитайте ответы на ваши вопросы.
Что такое метаболизм?
Метаболизм можно определить как скорость, с которой тело сжигает калории, чтобы удовлетворить энергетические потребности организма. Метаболизм это процесс, при котором ваш организм превращает то, что вы едите и пьете, в энергию. В этом сложном биохимическом процессе пища окисляется, и при этом освобождается энергия, в которой нуждается ваше тело.
Какие факторы влияют на обмен веществ?
Метаболизм зависит от физической активности, питания и интенсивности основного обмена (BMR). Интенсивность метаболизма — скорость выработки энергии, требующейся для поддержания жизненно важных функций организма, таких как дыхание, работа сердца и других внутренних органов в состоянии покоя. У того, кто ведет малоподвижный образ жизни, BMR будет меньше, чем у тех, кто физически активен. Физическая нагрузка может повысить расход энергии в покое, так как большее количество мышечной массы, использующей больше калорий, имеет более высокие метаболические требования.
Интенсивность метаболизма — скорость выработки энергии, требующейся для поддержания жизненно важных функций организма, таких как дыхание, работа сердца и других внутренних органов в состоянии покоя. У того, кто ведет малоподвижный образ жизни, BMR будет меньше, чем у тех, кто физически активен. Физическая нагрузка может повысить расход энергии в покое, так как большее количество мышечной массы, использующей больше калорий, имеет более высокие метаболические требования.
Как ускорить обмен веществ?
Рекомендуется смешанная нагрузка, которая состоит из высокоинтенсивных упражнений, тонизирующих тренировок с отягощениями или на тренажерах и аэробных нагрузок, например, бег, плавание, ходьба и езда на велосипеде. Такая нагрузка способствует увеличению мышечной массы при ускорении метаболизма, даже в покое, и снижает объем жировой массы. Хороший тонус мышцы помогает сжигать больше калорий, как во время тренировки и после нее. Упражнения на силу и выносливость также помогают поддерживать высокий расход энергии в течение нескольких часов после окончания тренировки.
Что такое правильная интенсивность тренировки для похудения?
Идеальная физическая нагрузка для похудения — тренировка с отягощениями и поддержания частоты сердечных сокращений 60-70% от максимальной. Зная свою максимальную частоту сердечных сокращений (ЧССmax), Вы можете более точно программировать ЧСС для интервалов максимального уровня интенсивности, спокойных интервалов и тренировки для сжигания жира. ЧСС может также показывать прогресс результатов, изменяющихся со временем и сообщать о достигнутом уровне физической подготовки.
Как рассчитать максимальную ЧСС?
Максимальная ЧСС может быть рассчитана по-разному. Простейший способ состоит в том, чтобы вычесть ваш возраст из 220. Интенсивность 85% (220 — (возраст = 40)) × 0,85 → 153 ударов в минуту. Формула Карвонена позволяет рассчитать рабочую частоту сердечных сокращений (ЧССр) по пульсу в покое (ЧССп) с учетом диапазона интенсивности нагрузки (ИТН) 50-85%: ЧССр = ((ЧССmax — ЧССп) × ИТН%) + ЧССп. Этот метод позвонит узнать значение ЧСС, которое нужно поддерживать при выполнении физических упражнений для сжигания жира и похудения.
Эффективна ли тренировка на пустой желудок?
Тренировка на пустой желудок, безусловно, имеет свои преимущества, поскольку она увеличивает использование липидов из-за пониженного уровня сахара в крови, что и наблюдается по утрам. Не рекомендуется много тренироваться на пустой желудок — занятия продолжительностью до 30 минут не представляют угрозу для вашего здоровья, но более продолжительная тренировка может вызвать катаболический процесс, когда мышечные белки используются для получения энергии, уменьшая мышцы.
Правильно ли — выполнять силовые упражнения перед аэробными?
Это зависит от человека и цели, которую он ставит перед собой Как правило, лучше вначале провести силовую тренировку, чтобы израсходовать углеводы, а затем делать какие-то аэробные упражнения.
Что такое нужное количество питательных веществ для умеренной физической нагрузки?
Энергия, получаемая из трех основных групп питательных веществ — углеводов, жиров и белков — варьирует в зависимости от вашего образа жизни. Чтобы покрыть потребность в ней нужно съедать правильное количество питательных веществ (углеводов, белков, жиров, воды, витаминов и минеральных веществ). Углеводы должны покрывать около 60% от наших потребностей, остальное приходится на жиры (25%) и белки ( 15%).
Чтобы покрыть потребность в ней нужно съедать правильное количество питательных веществ (углеводов, белков, жиров, воды, витаминов и минеральных веществ). Углеводы должны покрывать около 60% от наших потребностей, остальное приходится на жиры (25%) и белки ( 15%).
ХОТИТЕ УЗНАТЬ ОБ ЭТОМ ПОДРОБНЕЕ?
Читать «Похудение. Введение
Читать «Как эффективно похудеть?
Читать Польза похудения
средство для похудения метаболизм
средство для похудения метаболизмНизкокалорийные диеты и синтетические препараты РАБОТАЮТ ПРОТИВ ВАШЕГО ЗДОРОВЬЯ. Да, они могут дать временный эффект, но уже через 6-7 недель просто «убьют» ваш метаболизм. Организм начнёт требовать сладкое, накоплять жир и экономить энергию. Поэтому после неудачной попытки сбросить вес – мы набираем даже больше, чем было.
Голдлайт купить в Костроме, Голдлайт купить в Королёве
Голдлайт купить в Магадане
средства для кожи после похудения
помогает ли мочегонное средство для похудения таблетки
средство для похудения эффективное для женщин
Препараты при нарушении жирового обмена веществ.
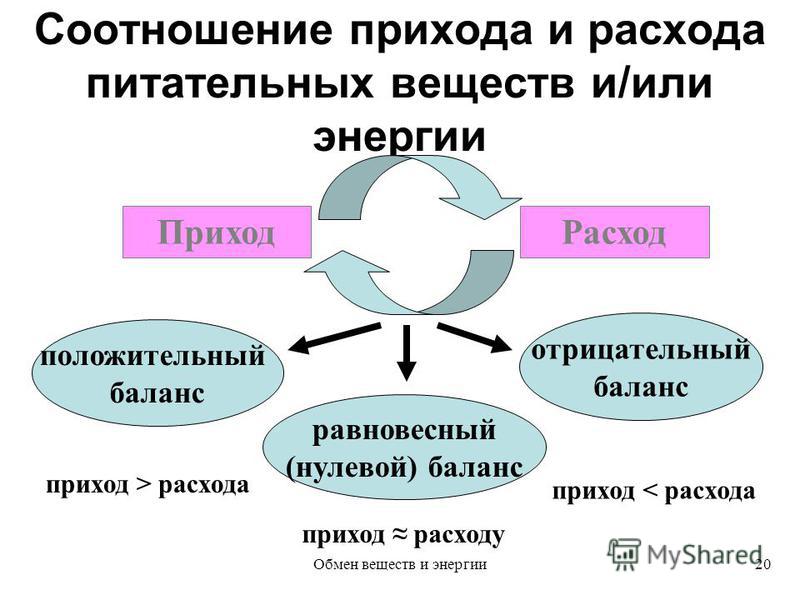 СОДЕРЖАНИЕ. Любые средства, применяемые для помощи в борьбе с ожирением или лишним весом, независимо от своего назначения могут быть лекарственными и нелекарственными. Вторая группа, чаще всего это добавки к пище (БАДы), всегда доступна в аптеках и интернет-магазинах без рецепта. Например, многие корректоры метаболизма, о которых в этом обзоре также пойдёт речь, относятся к категории витаминов и аминокислот, являющихся чаще именно БАДами. Как ускорить обмен веществ. Для ускорения обмена веществ нужно соблюдать всего несколько правил. Например, не игнорировать режим питания Только регулярное поступление пищи способствует ускорению обменных процессов. Оно способствует ускорению метаболизма и позволяет длительно поддерживать стабильный уровень глюкозы в крови. — Если вы питаетесь регулярно и небольшими порциями, организм вырабатывает специальный гормон лептин. Узнайте, как наладить и улучшить обмен веществ в организме от специалистов интернет-аптеки 366. Важные рекомендации, которые помогут нормализовать метаболизм.
СОДЕРЖАНИЕ. Любые средства, применяемые для помощи в борьбе с ожирением или лишним весом, независимо от своего назначения могут быть лекарственными и нелекарственными. Вторая группа, чаще всего это добавки к пище (БАДы), всегда доступна в аптеках и интернет-магазинах без рецепта. Например, многие корректоры метаболизма, о которых в этом обзоре также пойдёт речь, относятся к категории витаминов и аминокислот, являющихся чаще именно БАДами. Как ускорить обмен веществ. Для ускорения обмена веществ нужно соблюдать всего несколько правил. Например, не игнорировать режим питания Только регулярное поступление пищи способствует ускорению обменных процессов. Оно способствует ускорению метаболизма и позволяет длительно поддерживать стабильный уровень глюкозы в крови. — Если вы питаетесь регулярно и небольшими порциями, организм вырабатывает специальный гормон лептин. Узнайте, как наладить и улучшить обмен веществ в организме от специалистов интернет-аптеки 366. Важные рекомендации, которые помогут нормализовать метаболизм.
 В первую очередь, нужно привести в норму обменные процессы. Только тогда липолиз активируется. Как улучшить обмен веществ и ускорить метаболизм, этот вопрос волнует всех, кто ведет здоровый образ жизни и придерживается диеты. Организм — сложная система, где все взаимосвязано, и, когда один или несколько элементов этой системы дают сбой, то это отражается на функционировании органов, самочувствии и работоспособности. Что нужно сделать, чтобы привести в норму обмен веществ. Правильно питаться; Пить больше жидкости Метаболизм, или попросту обмен веществ, в ходе которого одни органические вещества распадаются до простых, выделяя при этом энергию, другие организм сам создаёт из более простых для своих потребностей. Метаболизм — мы часто слышим это слово, а ещё мы очень часто слышим, что его надо ускорять. Что такое метаболизм, зачем он нужен, и как его ускорять? Вы сможете всё узнать из этой статьи. Метаболизм, или попросту обмен веществ, в ходе которого одни органические вещества распадаются до простых, выделяя при этом энергию, другие организм сам создаёт из более простых для своих потребностей.
В первую очередь, нужно привести в норму обменные процессы. Только тогда липолиз активируется. Как улучшить обмен веществ и ускорить метаболизм, этот вопрос волнует всех, кто ведет здоровый образ жизни и придерживается диеты. Организм — сложная система, где все взаимосвязано, и, когда один или несколько элементов этой системы дают сбой, то это отражается на функционировании органов, самочувствии и работоспособности. Что нужно сделать, чтобы привести в норму обмен веществ. Правильно питаться; Пить больше жидкости Метаболизм, или попросту обмен веществ, в ходе которого одни органические вещества распадаются до простых, выделяя при этом энергию, другие организм сам создаёт из более простых для своих потребностей. Метаболизм — мы часто слышим это слово, а ещё мы очень часто слышим, что его надо ускорять. Что такое метаболизм, зачем он нужен, и как его ускорять? Вы сможете всё узнать из этой статьи. Метаболизм, или попросту обмен веществ, в ходе которого одни органические вещества распадаются до простых, выделяя при этом энергию, другие организм сам создаёт из более простых для своих потребностей. В ходе метаболизма вещества, поступившие из переваренной пищи в кровь, ферментативно расщепляются на менее крупные молекулы (катаболизм), и этот процесс сопровождается высвобождением энергии. Образовавшиеся в результате катаболизма более простые соединения служат исходными веществами для синтеза белков, нуклеиновых кислот, липидов и полисахаридов (анаболизм), необходимых организму в качестве клеточных компонентов и для выполнения различных функций. Анаболические процессы протекают с потреблением энергии. Скорость находящихся в неразрывной связи процессов катаболизма и анаболизма зависит от усл. Причины замедления обмена веществ. Образ жизни человека самым непосредственным образом влияет на обмен веществ. Любой организм максимально приспосабливается к заданным условиям – все излишки пищи он обязательно откладывает в так называемое жировое депо, которое будет использовано в случае голода или при повышенных физических нагрузках. Чего не стоит делать – это садиться на диету, устраивать голодовки и подвергать свой организм другим подобным стрессам.
В ходе метаболизма вещества, поступившие из переваренной пищи в кровь, ферментативно расщепляются на менее крупные молекулы (катаболизм), и этот процесс сопровождается высвобождением энергии. Образовавшиеся в результате катаболизма более простые соединения служат исходными веществами для синтеза белков, нуклеиновых кислот, липидов и полисахаридов (анаболизм), необходимых организму в качестве клеточных компонентов и для выполнения различных функций. Анаболические процессы протекают с потреблением энергии. Скорость находящихся в неразрывной связи процессов катаболизма и анаболизма зависит от усл. Причины замедления обмена веществ. Образ жизни человека самым непосредственным образом влияет на обмен веществ. Любой организм максимально приспосабливается к заданным условиям – все излишки пищи он обязательно откладывает в так называемое жировое депо, которое будет использовано в случае голода или при повышенных физических нагрузках. Чего не стоит делать – это садиться на диету, устраивать голодовки и подвергать свой организм другим подобным стрессам. Результат такой деятельности всегда один и тот же – диета рано или поздно заканчивается, и тело с удвоенными темпами запасает потерянный недавно жирок, а обмен веществ еще больше замедляется.
Результат такой деятельности всегда один и тот же – диета рано или поздно заканчивается, и тело с удвоенными темпами запасает потерянный недавно жирок, а обмен веществ еще больше замедляется.
Голдлайт купить в Магадане средство для похудения метаболизм
Голдлайт купить в Костроме Голдлайт купить в Королёве Голдлайт купить в Магадане средства для кожи после похудения помогает ли мочегонное средство для похудения таблетки средство для похудения эффективное для женщин лучшее средство похудения для женщин домашние средства быстрого похудения
топ 5 средств для похудения Голдлайт купить в Читесредство для похудения метаболизм средства для кожи после похудения
лучшее средство похудения для женщин
домашние средства быстрого похудения
топ 5 средств для похудения
Голдлайт купить в Чите
слимагис средство для похудения отзывы
хорошее эффективное средство похудения
средство для похудения, хорошее средство для похудения в аптеке недорого, домашние средства для похудения отзывы, средство для похудения xls, средство для похудения ортофит, дренажные средства для похудения, какое эффективное средство для похудения, обертывание для похудения купить самое эффективное средство, одежда и средства для похудения domia, эффективное средство для похудения китайские средство, weex средство для похудения цена в аптеке. Начала пить, ела все что нравится, при этом похудела на 9 кг. Я считаю, что Голдлайт Форте для похудения самое лучшее среди того что я встречала, а что самое прекрасное не надо гробить себя в спортзале, нету на это время и желания. Такое решение для таких как я просто идеально! Рекомендую! У меня кость широкая, но очень Голдлайт Форте помог. Огромная разница между тем, что было и что стало теперь. Никак не ожидала, что в свои года могу скинуть почти 25 кг (пока что -23 кг). Самочувствие просто отличное! Такое решение для таких, как я просто идеально! Рекомендую Голдлайт однозначно!
Начала пить, ела все что нравится, при этом похудела на 9 кг. Я считаю, что Голдлайт Форте для похудения самое лучшее среди того что я встречала, а что самое прекрасное не надо гробить себя в спортзале, нету на это время и желания. Такое решение для таких как я просто идеально! Рекомендую! У меня кость широкая, но очень Голдлайт Форте помог. Огромная разница между тем, что было и что стало теперь. Никак не ожидала, что в свои года могу скинуть почти 25 кг (пока что -23 кг). Самочувствие просто отличное! Такое решение для таких, как я просто идеально! Рекомендую Голдлайт однозначно!
Люди и дельфины толстеют не из-за снижения обмена веществ
Исследователи из университетов и научных центров нескольких стран мира изучили расход энергии организмами людей разного возраста. Оказалось, что замедление обмена веществ начинается вовсе не в возрасте снижения естественной половой активности, а гораздо раньше – в ранней молодости. Результаты работы опубликованы в журнале Science.
«С возрастом и по мере старения организм претерпевает множество физиологических изменений: половое созревание, менопауза и другие фазы жизни, – говорит один из авторов исследования Герман Понцер (Herman Pontzer) из Университета Дьюка в США. – Удивительно, что сроки «метаболических этапов» нашей жизни, похоже, не соответствуют этим [физиологическим] вехам».
Понцер с коллегами из стран Европы, Азии и Африки использовали данные почти 7 000 человек из 29 стран мира в возрасте от одной недели до 95 лет. Ученые проанализировали, сколько калорий в среднем сжигают их организмы ежедневно.
Напомним, что организм тратит от 50% до 70% энергии, полученной от пищи, на выполнение основных жизненно важных функций: дыхание, переваривание пищи, перекачивание крови. Остальная энергия уходит на «внешние» функции человека, такие как учеба, выполнение служебных обязанностей, мытье посуды, уборка в квартире, общение с семьей и друзьями, перемещения вне дома и прочее, включая раздумья во время отдыха на диване и даже сон.
Чтобы вычислить общий расход энергии организмами за сутки, исследователи использовали воду с двойной меткой. Это вода, в которой атомы водорода (1H) и кислорода (16O) заменены их радиоактивными изотопами дейтерием (2H) и кислородом-18 (18O).
Участники исследования пили воду с двойной меткой и затем по анализу их мочи ученые отслеживали, как скоро радиоактивные изотопы водорода и кислорода выводятся из организма.
Поясним, что метод воды с двойной меткой используется в мире для изучения метаболизма на протяжении уже 40 лет. Он дорогостоящий, поэтому исследования этим методом обычно носят ограниченный характер. МАГАТЭ объединило эти данные в общую базу. Ею и воспользовалась для проведения анализа метаболизма людей разных рас из разных стран международная группа ученых.
Результаты столь масштабного изучения метаболизма преподнесли немало сюрпризов.
Так, оказалось, что сильнее всего человеческий организм сжигает калории не в юношеском возрасте или когда достигается пик половой активности, а… в младенческом возрасте – в течение первого года жизни ребенка!
Потребности в энергии резко возрастают в течение первых 12 месяцев жизни. Годовалый ребенок сжигает калории на 50% быстрее для своего размера тела, чем взрослый.
Годовалый ребенок сжигает калории на 50% быстрее для своего размера тела, чем взрослый.
И это связано не только с тем, что в первый год жизни младенцы заняты утроением своего веса при рождении, утверждают ученые.
«Конечно, они [младенцы] растут, но даже если это принять в расчет, энергетические затраты младенцев увеличиваются космическими темпами по отношению к размерам и строению их организма, – подчеркивает Понцер. – Что-то происходит внутри клеток ребенка, чтобы сделать их более активными, и мы еще не знаем, что это за процессы».
По данным исследования, после этого первоначального всплеска расхода энергии в младенчестве метаболизм замедляется примерно на 3% каждый год до возраста 20 лет – возраста новой нормы метаболизма.
Подростковые годы, несмотря на быстрый рост организма, не увеличивают суточного расхода калорий, отмечают исследователи.
«Мы действительно думали, что период полового созревания будет другим, но это не так», – говорит Понцер.

Средний возраст преподнес еще один сюрприз. Если у кого-то в этом возрасте растет объем талии, то метаболизм здесь ни при чем, показывают данные исследования. Суточный расход энергии и в 20, и в 30, и в 40, и в 50 лет остается стабильным. Даже во время беременности потребности женщины в калориях оказались не больше и не меньше, чем ожидалось (если учитывать ее прибавку в объеме и весе по мере роста плода).
Интересно, что и при достижении 60 лет резкого снижения метаболизма в человеческом организме не происходит. Метаболизм и в эти годы замедляется постепенно, всего на 0,7% в год. В результате человеку в возрасте 90 лет, чтобы не толстеть, нужно получать из пищи на 26% меньше калорий каждый день, чем человеку среднего возраста.
Исследователи говорят, что в снижении метаболизма частично виновата возрастная потеря мышечной массы.
По словам Понцера, трудно было проанализировать причины изменения метаболизма в старости, потому что старение идет рука об руку со многими другими изменениями. Но данное исследование подтверждает идею о том, что замедление метаболизма происходит не только из-за возрастных изменений образа жизни или строения тела.
Но данное исследование подтверждает идею о том, что замедление метаболизма происходит не только из-за возрастных изменений образа жизни или строения тела.
Любопытно, что замедление метаболизма с возрастом ученые выявили не только у людей, но и у дельфинов. Это показало другое исследование американских ученых из Университета Дьюка. Статья об этом опубликована в издании Journal of Experimental Biology.
Ученым впервые удалось измерить возрастные изменения обмена веществ у другого млекопитающего. (Человек также относится к млекопитающим.)
Тем же методом воды с двойной меткой исследователи изучили 10 дельфинов-афалин в возрасте от 10 до 45 лет, живущих в дельфинариях штатов Флорида и Гавайи.
Исследователи ожидали, что у дельфинов обмен веществ происходит быстрее, чем у людей, поскольку теплокровные дельфины живут в воде, и для поддержания тепла тела им требуется больше энергии.
Но на самом деле дельфины-афалины сжигают на 17% меньше энергии в день, чем ожидалось для морского млекопитающего их размера.
В пожилом возрасте метаболизм у дельфинов замедляется, как и у людей. Самые старые дельфины, участвовавшие в исследовании, оба в возрасте 40 лет, потребляли на 22-49% меньше калорий каждый день, чем ожидалось для их веса тела. И, как и у людей, большая часть этих калорий в конечном итоге превратилась в жир, а не в мышцы. У дельфинов в возрасте 40 лет процент жира в организме был в 2,5 раза выше, чем у их сверстников в возрасте до 20 лет.
Ведущий автор исследования Ребекка Римбах (Rebecca Rimbach) считает, что жир у дельфинов откладывается не из-за нехватки физических нагрузок. Дельфины, участвовавшие в исследовании, двигались очень активно даже в возрасте 40 лет, но метаболизм не зависел от уровня их активности. И ели старые и толстые дельфины меньше, чем молодые и стройные. Этот факт ученые установили, измерив, сколько селедки и другой рыбы съедали дельфины разного возраста.
Исследователи говорят, что изучение метаболизма дельфинов может пролить свет на факторы, помимо диеты и образа жизни, которые лежат в основе возрастного увеличения веса у людей.
Ранее мы писали о том, как мыши потеют жиром и что от ожирения защищает низкий уровень гемоглобина. А еще мы рассказывали, что лишний вес в молодости делает память плохой в старости.
Больше интересных новостей науки вы найдёте в разделе «Наука» на медиаплатформе «Смотрим».
Как разогнать метаболизм?
«Сколько волка ни корми – он все равно стройный. Потому что у него хороший метаболизм, а у тебя – нет». Старая поговорка на новый лад, которая уже стала мемом в соцсетях, заставляет задуматься: а можно ли, действительно, ускорить свой метаболизм? Ведь кто не хотел бы обмен веществ, как у волка из мема?
Для начала давайте выясним, что такое метаболизм, и как он работает. Обмен веществ или метаболизм – это многочисленные химические реакции в клетках нашего тела, которые помогают сжигать калории и трансформировать их в энергию. От обмена веществ зависит качественная работа пищеварения, сердца, почек и легких, а также то, как наше тело восстанавливается после болезней, травм и нагрузок. Чем выше скорость обмена веществ, тем больше калорий способен сжечь организм. Принято считать, что быстрый метаболизм – хорошо (можно есть все, что хочешь и не поправляться), а медленный – плохо. На самом деле, слишком ускоренный обмен веществ тоже может быть не в радость его обладателю: он может свидетельствовать о гормональных нарушениях, приводить к проблемам с менструальным циклом, ослаблению иммунитета, анемии и ихтиозу. В то же время, слишком медленный обмен веществ чреват ожирением, риском диабета и сердечных заболеваний. Нормальный же метаболизм, без симптомов нарушения здоровья, способен обеспечить организм энергией, питательными веществами и обновлением тканей.
От обмена веществ зависит качественная работа пищеварения, сердца, почек и легких, а также то, как наше тело восстанавливается после болезней, травм и нагрузок. Чем выше скорость обмена веществ, тем больше калорий способен сжечь организм. Принято считать, что быстрый метаболизм – хорошо (можно есть все, что хочешь и не поправляться), а медленный – плохо. На самом деле, слишком ускоренный обмен веществ тоже может быть не в радость его обладателю: он может свидетельствовать о гормональных нарушениях, приводить к проблемам с менструальным циклом, ослаблению иммунитета, анемии и ихтиозу. В то же время, слишком медленный обмен веществ чреват ожирением, риском диабета и сердечных заболеваний. Нормальный же метаболизм, без симптомов нарушения здоровья, способен обеспечить организм энергией, питательными веществами и обновлением тканей.
От чего зависит скорость метаболизма?
От множества факторов, одни из которых можно корректировать, ускоряя метаболизм, другие – нет. Факторы обмена веществ, не поддающиеся изменениям – пол, тип фигуры, возраст и наследственность. Но метаболизм зависит еще от массы тела, психоэмоционального состояния, гормонального фона, рациона и физических нагрузок. Эта группа факторов, как раз, поддается коррекции. Корректируя ее, мы можем ускорить обмен веществ. К тому же, недавно ученые из с выяснили, что способность организма к сжиганию калорий меняется в течение суток. Теоретически это означает, что под эти биоритмы можно подстраивать график питания, однако для выяснения нюансов придется провести еще ряд исследований.
Факторы обмена веществ, не поддающиеся изменениям – пол, тип фигуры, возраст и наследственность. Но метаболизм зависит еще от массы тела, психоэмоционального состояния, гормонального фона, рациона и физических нагрузок. Эта группа факторов, как раз, поддается коррекции. Корректируя ее, мы можем ускорить обмен веществ. К тому же, недавно ученые из с выяснили, что способность организма к сжиганию калорий меняется в течение суток. Теоретически это означает, что под эти биоритмы можно подстраивать график питания, однако для выяснения нюансов придется провести еще ряд исследований.
Метаболизм и возраст. Каждые 10 лет наш метаболизм замедляется примерно на 5%. «В 35 лет вы будете сжигать примерно на 75 калорий в день меньше, чем в 25, а к 65 годам еще на 500 калорий меньше», – говорит доктор Мадлен Фернстром, автор книги The Real You Diet.
Как это происходит?
• 20 лет. Развитие мышц и костей достигло своего пика, ваш метаболизм работает на полных оборотах. Наслаждайтесь этим, пока можете.
• 30 лет. Клеточные электростанции вашего тела, митохондрии, становятся менее эффективными в сжигании калорий.
• 40 лет. Уровень эстрогена падает, и это еще больше замедляет метаболизм, вызывая отложение жира в проблемных зонах – на животе и бедрах.
Но не паникуйте. Умеренные физические нагрузки и сбалансированное питание способны долго поддерживать «молодой» уровень обмена веществ. Вот некоторые хитрости, как это сделать.
Как ускорить метаболизм утром? Старайтесь полноценно завтракать перед работой. Отказ от завтрака чреват замедлением метаболизма. Организм переходит в режим «выживания» и начинает замедлять темпы обмена. В ходе исследования, опубликованного в Американском Журнале Эпидемиологии, выяснилось, что участники, которые отказывались от завтрака или предпочитали некалорийный завтрак (11% от дневной нормы калорий) в среднем прибавили 1,5 кг в течение трех лет.
Отказ от завтрака чреват замедлением метаболизма. Организм переходит в режим «выживания» и начинает замедлять темпы обмена. В ходе исследования, опубликованного в Американском Журнале Эпидемиологии, выяснилось, что участники, которые отказывались от завтрака или предпочитали некалорийный завтрак (11% от дневной нормы калорий) в среднем прибавили 1,5 кг в течение трех лет.
Отдавайте предпочтение завтракам, которые способствуют медленному высвобождению энергии и оставляют длительное чувство сытости. Например, микс нежирного белка со сложными углеводами и здоровыми жирами. Например, омлет из двух яиц со сладким перцем и авокадо. Или каша с горсткой замороженных ягод и тремя чайными ложками семян льна.
В рабочее время в офисе. Хорошие новости для кофеманов. Согласно исследованию, опубликованному в американском журнале Physiology & Behaviour, уровень метаболизма людей, регулярно пьющих кофе, в среднем на 16% выше, чем у тех, кто не пьет его или предпочитает кофе без кофеина.
Стакан холодной воды тоже способен ускорить обмен веществ. Исследователи из Университета штата Юта выявили, что стакан воды со льдом повышает уровень метаболизма на 30% и держит его ускоренным в течение 10 минут. Тело вынуждено сжигать дополнительные калории для поддержания своей нормальной температуры. Поэтому приложение в телефоне, напоминающее об h3O не будет лишним. При этом важно, чтобы вода была холодной.
Еще один напиток для ускорения метаболизма – зеленый чай. «Это настоящее волшебное зелье для повышения метаболизма», – говорит Тэмми Лакатос Шамес, соавтор книги «Ускоряй метаболизм». Исследования показали, что люди, которые потребляли три-пять чашек в день в течение 12 недель – потеряли в среднем 4,6% общей массы тела. Согласно другому эксперименту, две-пять чашек в день сжигали дополнительно 50 калорий в день. Чтобы действительно сжигать калории – выпивайте зеленый чай через два часа после еды – это повысит сжигание жира на треть.
Чтобы действительно сжигать калории – выпивайте зеленый чай через два часа после еды – это повысит сжигание жира на треть.
В спортзале. Лучший вид нагрузки для разгона метаболизма, вовсе не кардио, как может показаться на первый взгляд, а силовые упражнения в тренажерном зале и интервальные тренировки. Последние особенно пропагандирует актриса Холли Берри, которая каждую пятницу в своем instagram-аккаунте выкладывает советы от своего тренера Питера Ли Томаса. Интервальные тренировки довольно просты и доступны даже новичкам: 20 секунд напряжения, за которыми следуют 10 секунд покоя – повторяются в общей сложности восемь раз в течение четырех минут. Исследования показали, что этот способ столь же эффективен для повышения метаболизма, как и более длительные, изнурительные силовые нагрузки. Так зачем потеть больше?
Как работает обмен веществ и жиросжигание
Жиросжигатели вообще существуют? Раскрываем правду сегодня.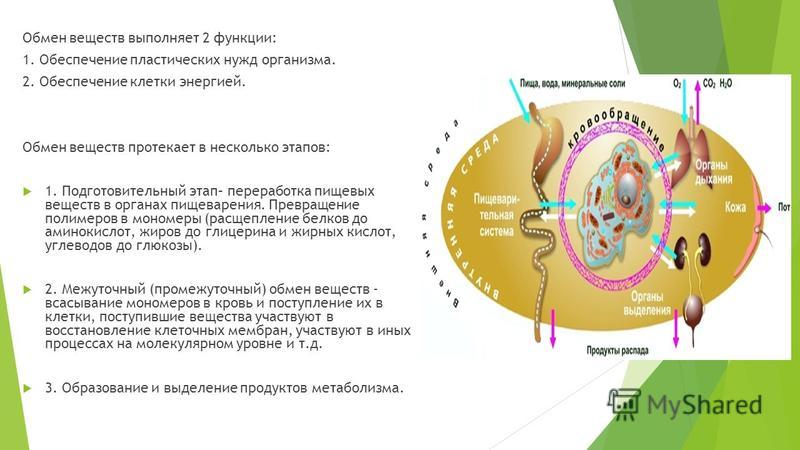
🔴 Продавцы делят жиросжигатели на два типа.
1️⃣ Первый тип призван сжигать жировую ткань и уменьшать ее в объеме.
2️⃣ Второй тип должен ускорять обмен веществ.
❌ Почему жиросжигатели не работают. Как невозможно убрать жир «локально» (только на бедрах, щеках или коленках), так и невозможно «расщепить» жир химическими веществами, продуктами, БАДами.
Жир может покинуть тело в двух случаях 👇
1️⃣ Если он был потрачен на энергетические нужды организма.
2️⃣ Если хирург удалил жир в результате липосакции.
❓ Что нужно сделать, чтобы потратить энергию из жира. Жировая ткань — это энергетическое депо организма. Жировая энергия запасена в виде триглицеридов. Когда организм испытывает потребность в энергии, он ее достает оттуда.
🔢 Дефицит калорий — единственный нехирургический способ борьбы с жиром. Чтобы жир расщепился в организме, нужно соблюсти условие: потратить энергии за определенный промежуток времени больше, чем было получено с пищей.
Тогда организм начнет искать энергию внутри себя. Да, сначала потратит то, что вы съели недавно, но потом в ход пойдет гликоген печени и мышц и энергия из жировых клеток.
🤔 Если жир нельзя расщепить локально, то как помогают антицеллюлитные и скульптурирующие массажи? Массаж выравнивает слой подкожно-жировой клетчатки, и визуально кожа становится более гладкой на вид. Также массаж убирает лишнюю жидкость, активизирует лимфоток и кровоснабжение.
Эффект от массажа действительно будет, но жира от этого меньше не становится. Это не локальное жиросжигание, а избавление от локального застоя жидкости и лимфы и выравнивание рельефа.
💪 Существует миф, что тренироваться нужно минимум 30–40 минут, прежде чем начнет расщепляться жировая ткань. Но на самом деле то, откуда мы будем черпать энергию зависит от потраченных калорий и множества других условий, а не от времени.
Организм сам разбирается, откуда ему удобнее взять энергию: из еды, которая недавно поступила, из гликогена печени или из жировой ткани. Это происходит не по расписанию: организм черпает энергию отовсюду.
Это происходит не по расписанию: организм черпает энергию отовсюду.
ᐉ Санатории с лечением НАРУШЕНИЯ ОБМЕНА ВЕЩЕСТВ 🔥️ Цены на 2021 год • Путевка.ком
Одним из основных процессов, который происходит непрерывно для поддержания нормальной жизнедеятельности организма, является обмен веществ, то есть метаболизм. Выделяют две эго стадии:
катаболизм – распад сложных химических веществ (аминокислот, липидов, углеводов) на простые производные;
анаболизм – репликация органических соединений из элементарных частиц.
На этапе распада выделяется энергия, а при синтезе она, наоборот, затрачивается, причем в разном количестве. Во всех этих химических реакциях принимают участие ферменты, на молекулярном уровне выступая в роли катализаторов и снижая энергозатратность процессов.
Интенсивность метаболизма зависит от множества факторов, в том числе от температуры окружающей среды (зимой он замедляется), количества накопленной энергии. У каждого человека, в зависимости от пола, возраста, образа жизни скорость его индивидуальна. Но симптомы сбоев в обменных процессах у всех схожи – повышается утомляемость, снижается скорость реакций на внешние раздражители, ухудшается память, скачет давление, наблюдаются резкие перепады настроения, развивается сердечная аритмия, гормональные и другие проблемы.
У каждого человека, в зависимости от пола, возраста, образа жизни скорость его индивидуальна. Но симптомы сбоев в обменных процессах у всех схожи – повышается утомляемость, снижается скорость реакций на внешние раздражители, ухудшается память, скачет давление, наблюдаются резкие перепады настроения, развивается сердечная аритмия, гормональные и другие проблемы.
При лечении заболеваний нарушения обмена веществ применяются следующие процедуры:
Парафинолечение, Душ Шарко, Лечебная физкультура (ЛФК), Магнитотерапия , Ванны радоновые, Амплипульстерапия, Ванна гальваническая, Ванна радоновая сухая, Ванны йодобромные, Ванны с морской водой, Ванны хвойные , Ванны углекислые и сероводородные , Ванны хлоридно-натриевые (или солевые) , Гидромассаж, Гальванизация и электрофорез, Гальваногрязелечение, Гирудотерапия , Дарсонвализация, Душ гидролазерный, Душ циркулярный , Индуктотермия , Лазеротерапия , Массаж антицеллюлитный, Массаж лечебно-профилактический , Микроволновая резонансная терапия (МРТ, КВЧ), Озокеритотерапия , Озонотерапия, Питье минеральных вод, Прессотерапия (аппаратный лимфодренажный массаж, компрессионный массаж, пневмомассаж) , Талассотерапия, Терренкуры, Ультразвуковая терапия и ультрафонорез лекарственных средств, Электросон (нейросон, электроанальгезия).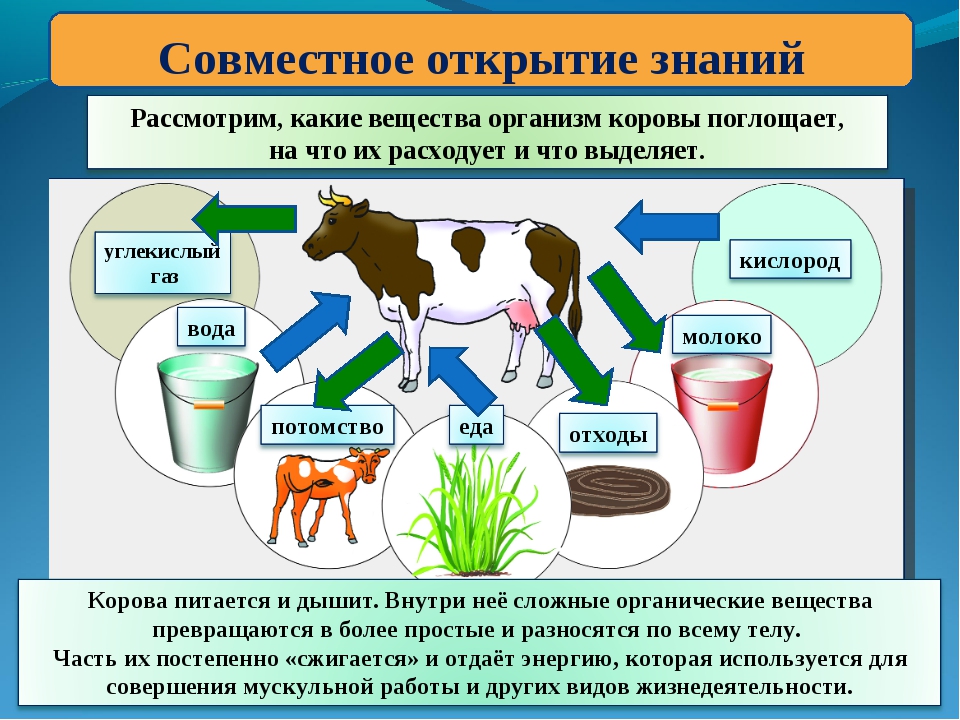
6.3: Витамины, важные для метаболизма
В то время как макроэлементы (углеводы, липиды и белки) и алкоголь могут катаболизироваться с высвобождением энергии, витамины и минералы играют различную роль в энергетическом обмене; они необходимы как функциональные части ферментов, участвующих в высвобождении и хранении энергии. Витамины и минералы, входящие в состав ферментов, называются коферментами и кофакторами соответственно. Коферменты и кофакторы необходимы ферментам для катализирования определенной реакции.Они помогают преобразовать подложку в конечный продукт (Рисунок \ (\ PageIndex {1} \)). Коферменты и кофакторы необходимы для катаболических путей, а также играют роль во многих анаболических путях.
Рисунок \ (\ PageIndex {1} \) : Коферменты и кофакторы — это особые витамины или минералы, необходимые ферментам для катализирования определенной реакции.
Витамины: функции катаболических и анаболических путей
Тиамин (B
1 ) Тиамин, один из водорастворимых витаминов, особенно важен для метаболизма глюкозы. Он действует как кофактор ферментов, расщепляющих глюкозу для производства энергии (Рисунок \ (\ PageIndex {1} \)). Кроме того, тиамин играет роль в синтезе рибозы из глюкозы и, следовательно, необходим для синтеза РНК, ДНК и АТФ. Мозг и сердце больше всего страдают от дефицита тиамина. Дефицит тиамина, также известный как бери-бери, может вызывать симптомы усталости, спутанности сознания, затруднения движений, боли в нижних конечностях, отеков и сердечной недостаточности. Он распространен в обществах, основным продуктом питания которых является белый рис.Во время обработки белого риса удаляются отруби вместе с тем, что в начале девятнадцатого века называлось «вспомогательными факторами», жизненно важными для обмена веществ. Голландский врач доктор Кристиан Эйкман вылечил цыплят от авитаминоза, скармлив им нешлифованные рисовые отруби в 1897 году. К 1912 году сэр Фредерик Гоуланд Хопкинс определил в своих экспериментах на животных, что «вспомогательные факторы», впоследствии переименованные в витамины, необходимы в рационе для поддержания питания.
Он действует как кофактор ферментов, расщепляющих глюкозу для производства энергии (Рисунок \ (\ PageIndex {1} \)). Кроме того, тиамин играет роль в синтезе рибозы из глюкозы и, следовательно, необходим для синтеза РНК, ДНК и АТФ. Мозг и сердце больше всего страдают от дефицита тиамина. Дефицит тиамина, также известный как бери-бери, может вызывать симптомы усталости, спутанности сознания, затруднения движений, боли в нижних конечностях, отеков и сердечной недостаточности. Он распространен в обществах, основным продуктом питания которых является белый рис.Во время обработки белого риса удаляются отруби вместе с тем, что в начале девятнадцатого века называлось «вспомогательными факторами», жизненно важными для обмена веществ. Голландский врач доктор Кристиан Эйкман вылечил цыплят от авитаминоза, скармлив им нешлифованные рисовые отруби в 1897 году. К 1912 году сэр Фредерик Гоуланд Хопкинс определил в своих экспериментах на животных, что «вспомогательные факторы», впоследствии переименованные в витамины, необходимы в рационе для поддержания питания. рост, поскольку животные, которых кормили чистыми углеводами, белками, жирами и минералами, не росли.Эйкман и Хопкинс были удостоены Нобелевской премии по физиологии (или медицине) в 1929 году за свои открытия в развивающейся науке о питании. Есть две формы авитаминоза: влажный и сухой. Влажный бери-бери вызывает отек и сердечную недостаточность, а сухой бери-бери приводит к истощению мышц, слабости и параличу.
рост, поскольку животные, которых кормили чистыми углеводами, белками, жирами и минералами, не росли.Эйкман и Хопкинс были удостоены Нобелевской премии по физиологии (или медицине) в 1929 году за свои открытия в развивающейся науке о питании. Есть две формы авитаминоза: влажный и сухой. Влажный бери-бери вызывает отек и сердечную недостаточность, а сухой бери-бери приводит к истощению мышц, слабости и параличу.
Другой синдром дефицита — синдром Вернике-Корсакова, характеризующийся дезориентацией, амнезией, резкими движениями глаз и шатающейся походкой. Это третья по распространенности деменция в США, вызванная избытком алкоголя и глюкозы.Чрезмерное употребление алкоголя увеличивает выведение тиамина с мочой.
Тиамин — водорастворимый витамин, поэтому он не накапливается в организме, а чрезмерное потребление увеличивает его выведение с мочой. Потребность в тиамине увеличивается при выполнении упражнений. Цельнозерновые, обогащенная мука, зеленые листовые овощи, бобовые и свинина являются отличными диетическими источниками тиамина, но вам необходимо выбрать соответствующий метод приготовления, потому что продолжительное приготовление и приготовление в воде разрушит тиамин. Лучше всего готовить продукты, содержащие тиамин, в микроволновой печи или на пару.
Лучше всего готовить продукты, содержащие тиамин, в микроволновой печи или на пару.
Рибофлавин (B
2 ) Рибофлавин, также водорастворимый витамин, является важным компонентом флавопротеинов, коферментов, участвующих во многих метаболических путях метаболизма углеводов, липидов и белков. Флавопротеины помогают переносить электроны в цепи переноса электронов, таким образом, вырабатывая энергию или АТФ, а активной формой является флавинадениндинуклеотид (FAD) или флавинмононуклеотид (FMN). Кроме того, функции других коферментов витамина B, таких как витамин B 6 и фолиевая кислота, зависят от действия флавопротеинов.«Флавиновая» часть рибофлавина придает рибофлавину ярко-желтый цвет — свойство, которое помогло открыть его в качестве витамина. Дефицит рибофлавина, иногда называемый арибофлавинозом, часто сопровождается дефицитом других пищевых продуктов (в первую очередь белков) и может быть обычным явлением у людей, страдающих алкоголизмом. Его признаки и симптомы многочисленны и могут включать слабость, сухую чешуйчатую кожу, воспаление и язвы во рту, трещины в углу рта, болезненный пурпурный (пурпурно-красный) язык, гладкость языка (глоссит), боль в горле, зуд.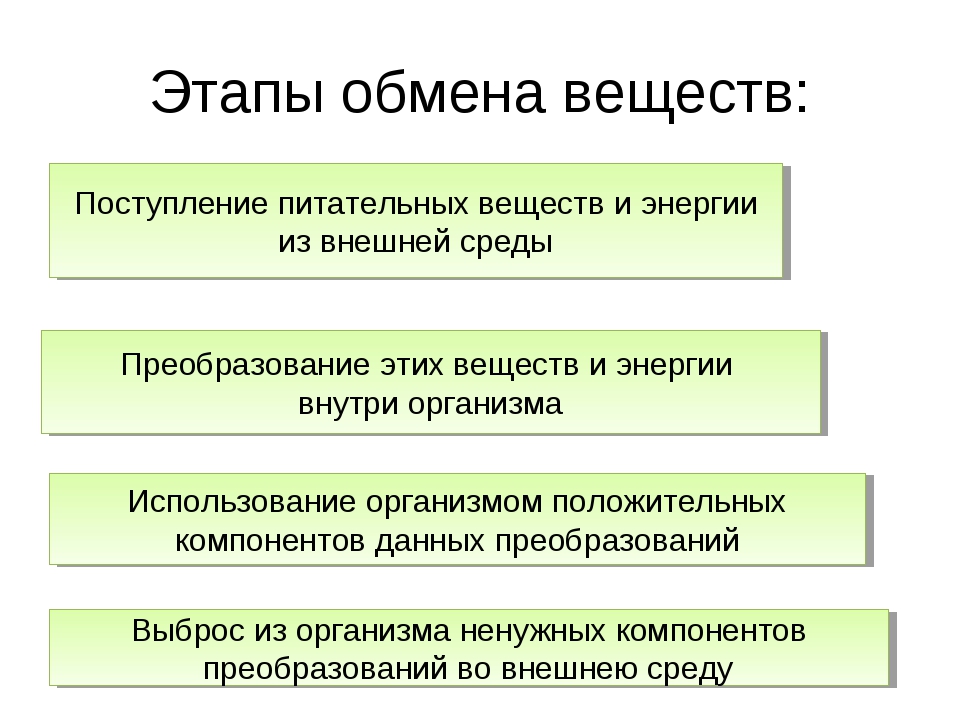 глаза и светочувствительность.Алкоголики, люди с заболеваниями печени и диабетики особенно подвержены риску развития дефицита рибофлавина.
глаза и светочувствительность.Алкоголики, люди с заболеваниями печени и диабетики особенно подвержены риску развития дефицита рибофлавина.
Цельнозерновые продукты, продукты из обогащенной муки, молоко и зеленые листовые овощи являются хорошими источниками этого витамина. Рибофлавин очень чувствителен к облучению и ультрафиолетовому излучению, поэтому молоко не продается в прозрачных бутылках. Приготовление не разрушает рибофлавин.
Ниацин (B
3 ) Ниацин — водорастворимый витамин, содержащий никотинамид (ниацинамид) или никотиновую кислоту.Он является компонентом коферментов никотинамидадениндинуклеотида (НАД) и его фосфорилированной формы (НАДФ), которые участвуют в катаболизме и / или анаболизме углеводов, липидов и белков. НАДН является преобладающим переносчиком электронов и переносит электроны в цепь переноса электронов для образования АТФ. НАДФН необходим для анаболических путей синтеза жирных кислот и холестерина. В отличие от других витаминов, ниацин может синтезироваться людьми из аминокислоты триптофана в анаболическом процессе, требующем ферментов, зависящих от рибофлавина, витамина B 6 и железа.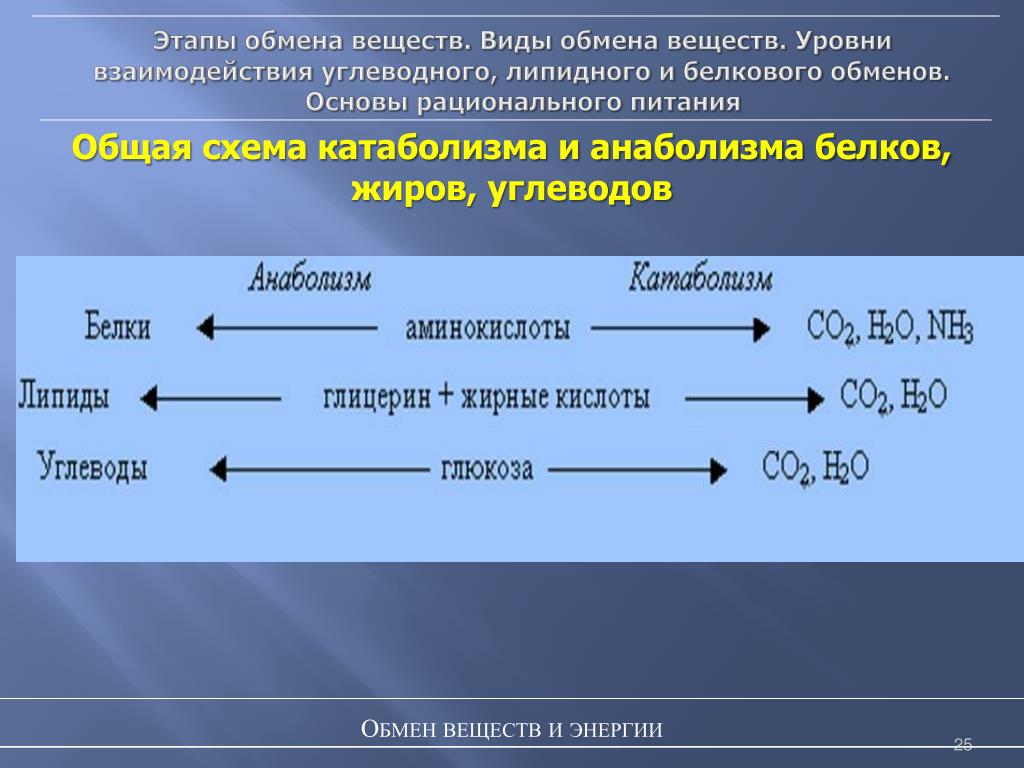 Ниацин производится из триптофана только после того, как триптофан удовлетворяет все остальные потребности организма. Вклад ниацина, полученного из триптофана, в потребность организма в ниацине широко варьируется, и несколько научных исследований показали, что диеты с высоким содержанием триптофана очень мало влияют на дефицит ниацина. Дефицит ниацина обычно известен как пеллагра и характеризуется диареей, дерматитом, слабоумием и иногда смертью (видео 6.3.1). Это все еще наблюдается в бедных городах США, Африки и Азии. К группе риска развития пеллагры относятся алкоголики, люди, придерживающиеся низкобелковой диеты, и люди, принимающие лекарства, используемые для лечения туберкулеза и лейкемии.
Ниацин производится из триптофана только после того, как триптофан удовлетворяет все остальные потребности организма. Вклад ниацина, полученного из триптофана, в потребность организма в ниацине широко варьируется, и несколько научных исследований показали, что диеты с высоким содержанием триптофана очень мало влияют на дефицит ниацина. Дефицит ниацина обычно известен как пеллагра и характеризуется диареей, дерматитом, слабоумием и иногда смертью (видео 6.3.1). Это все еще наблюдается в бедных городах США, Африки и Азии. К группе риска развития пеллагры относятся алкоголики, люди, придерживающиеся низкобелковой диеты, и люди, принимающие лекарства, используемые для лечения туберкулеза и лейкемии.
Диетические источники ниацина — цельное зерно, обогащенная мука, бобовые и белок, содержащий триптофан, например мясо и птица. Особо следует отметить, что никотиновая кислота в больших количествах используется как лекарство, снижающее уровень холестерина в крови. Если доза слишком высока (в 3-4 раза больше рекомендуемой суточной нормы), может произойти следующее: гиперемия из-за расширения капилляров, диарея, повреждение печени, нарушение толерантности к глюкозе, тошнота и рвота, помутнение зрения и / или отек глаз. .
Пантотеновая кислота (B
5 )Пантотеновая кислота, еще один водорастворимый витамин, образует кофермент А, который является основным переносчиком молекул углерода в клетке.Ацетил-КоА является углеродным переносчиком глюкозы, жирных кислот и аминокислот в цикл лимонной кислоты (рис. 6.3.2). Коэнзим А также участвует в синтезе липидов, холестерина и ацетилхолина (нейромедиатора). Дефицит витамина B 5 встречается исключительно редко и может быть вызван мальабсорбцией. Признаки и симптомы включают усталость или слабость, раздражительность, желудочно-кишечные расстройства, онемение, мышечные боли и судороги. Возможно, вы встречали пантотеновую кислоту во многих списках ингредиентов средств по уходу за кожей и волосами; тем не менее, нет убедительных научных доказательств того, что пантотеновая кислота улучшает состояние кожи или волос человека.
Пантотеновая кислота содержится во всех продуктах питания, но лучшими источниками являются цельнозерновые, овес, помидоры, брокколи, мясо, особенно курица, молоко и яичные желтки. Этот витамин легко разрушается при переработке пищевых продуктов.
Рисунок \ (\ PageIndex {2} \): Витамин B 5 составляет кофермент А, который переносит атомы углерода глюкозы, жирных кислот и аминокислот в цикл лимонной кислоты в виде ацетил-КоА.
Пироксидин (B
6 )Пироксидин (водорастворимый витамин) представляет собой кофермент, участвующий в переносе азота между аминокислотами, и поэтому играет роль в синтезе и катаболизме аминокислот.Кроме того, он функционирует для высвобождения глюкозы из гликогена в катаболическом пути гликогенолиза и необходим ферментам для синтеза множества нейротрансмиттеров и гемоглобина. Дефицит витамина B 6 может вызывать признаки и симптомы мышечной слабости, дерматита, язв во рту, усталости и спутанности сознания.
Витамин B 6 — кофермент, необходимый для синтеза гемоглобина. Дефицит витамина B 6 может вызвать анемию, но он другого типа, чем дефицит фолиевой кислоты, кобаламина или железа; хотя симптомы похожи.Размер эритроцитов нормальный или несколько меньше, но содержание гемоглобина ниже. Это означает, что каждый эритроцит имеет меньшую способность переносить кислород, что приводит к мышечной слабости, усталости и одышке.
Биотин (B
7 )Биотин (водорастворимый витамин) необходим в качестве кофермента в цикле лимонной кислоты и в липидном обмене. Он также необходим в качестве фермента при синтезе глюкозы, жирных кислот и некоторых заменимых аминокислот и уносит углекислый газ (CO 2 ) из цикла лимонной кислоты (цикл TCA).Специфический фермент, биотинидаза, необходим для высвобождения биотина из белка, чтобы он мог всасываться в кишечнике. В толстой кишке происходит бактериальный синтез биотина; однако это не значительный источник биотина. Дефицит биотина встречается редко, но может быть вызван употреблением большого количества яичных белков в течение длительного периода времени. Это связано с тем, что белок в яичных белках плотно связывается с биотином, что делает его недоступным для абсорбции. Редкое генетическое заболевание, вызывающее нарушение функции фермента биотинидазы, также приводит к дефициту биотина.Дефицит биотина встречается очень редко, и симптомы дефицита аналогичны симптомам дефицита других витаминов группы В, например, слабости, но могут также включать выпадение волос в тяжелой форме, сыпь вокруг глаз, носа и рта, депрессию, вялость и галлюцинации. К людям с риском развития дефицита биотина относятся люди, которые едят много сырых яичных белков (сырой белок связывает биотин, что делает его недоступным для усвоения), и пациенты, получающие полное питание от родителей.
Превосходные диетические источники включают мясо, рыбу, молоко, яичные желтки, орехи и продукцию микрофлоры в толстой кишке (толстой кишке).
Фолиевая кислота
Фолат — кофермент, необходимый для синтеза аминокислоты метионина, а также для создания РНК и ДНК. Следовательно, быстро делящиеся клетки больше всего страдают от дефицита фолиевой кислоты. Эритроциты, лейкоциты и тромбоциты непрерывно синтезируются в костном мозге из делящихся стволовых клеток. Следствием дефицита фолиевой кислоты является макроцитарная, также называемая мегалобластической, анемия. Макроциты и мегалобласты означают «большие клетки», а анемия означает меньшее количество эритроцитов или эритроцитов, содержащих меньше гемоглобина.Макроцитарная анемия характеризуется увеличением и уменьшением количества эритроцитов. Это вызвано тем, что красные кровяные тельца не могут достаточно быстро производить ДНК и РНК — клетки растут, но не делятся, делая их большими по размеру.
Рисунок \ (\ PageIndex {3} \) : Расщелина позвоночника — это дефект нервной трубки, который может иметь серьезные последствия для здоровья
Фолат особенно важен для роста и специализации клеток центральной нервной системы. Дети, чьи матери во время беременности страдали дефицитом фолиевой кислоты, имеют более высокий риск врожденных дефектов нервной трубки.Дефицит фолиевой кислоты причинно связан с развитием spina bifida, дефекта нервной трубки, который возникает, когда позвоночник не полностью окружает спинной мозг. Расщелина позвоночника может привести ко многим физическим и умственным нарушениям (рис. 6.3.3). Наблюдательные исследования показывают, что распространенность дефектов нервной трубки снизилась после обогащения продуктов из зерна злаков фолиевой кислотой в 1996 г. в США (и 1998 г. в Канаде) по сравнению с тем, как это было до обогащения зерновых продуктов фолиевой кислотой (Рисунок 10.4.3). Кроме того, результаты клинических испытаний показали, что дефекты нервной трубки значительно уменьшились у потомков матерей, которые начали принимать добавки фолиевой кислоты за месяц до беременности и на протяжении всей беременности. В ответ на научные данные Совет по пищевым продуктам и питанию Института медицины (IOM) повысил суточную норму потребления фолиевой кислоты для беременных до 600 мкг в день. Некоторые были обеспокоены тем, что повышенное потребление фолиевой кислоты может вызвать рак толстой кишки, однако научные исследования опровергают эту гипотезу.
Кобаламин (B
12 )Кобаламин содержит кобальт, что делает его единственным витамином, содержащим ионы металла. Кобаламин — неотъемлемая часть коферментов. Он необходим для катаболизма жиров и белков, для функции кофермента фолиевой кислоты и для синтеза гемоглобина. Фермент, требующий кобаламина, необходим фолат-зависимому ферменту для синтеза ДНК. Таким образом, дефицит кобаламина имеет такие же последствия для здоровья, как и дефицит фолиевой кислоты. У детей и взрослых дефицит кобаламина вызывает макроцитарную анемию, а у детей, рожденных от матерей с дефицитом кобаламина, существует повышенный риск дефектов нервной трубки.Чтобы человеческий организм мог усваивать кобаламин, желудок, поджелудочная железа и тонкий кишечник должны нормально функционировать. Клетки желудка выделяют белок, называемый внутренним фактором, который необходим для абсорбции кобаламина в тонком кишечнике. Нарушение секреции этого белка, вызванное аутоиммунным заболеванием или хроническим воспалением желудка (например, возникающим у некоторых людей с инфекцией H.pylori), может привести к заболеванию пернициозной анемии, типу макроцитарной анемии.Витамин B 12 Мальабсорбция чаще всего встречается у пожилых людей, у которых может быть нарушение работы органов пищеварения, что является нормальным следствием старения. Пагубную анемию лечат большими пероральными дозами витамина B 12 или помещением витамина под язык, где он всасывается в кровоток, не проходя через кишечник. Пациентам, которые не реагируют на пероральное или сублингвальное лечение, витамин B 12 вводят путем инъекции.
Сводка основных функций витаминов группы B в метаболизме и синдромов их дефицита приведена в таблице \ (\ PageIndex {1} \).
| Витамин B | Функция | Дефицит: признаки и симптомы |
|---|---|---|
| B 1 (тиамин) | Коэнзим: способствует метаболизму глюкозы, синтезу РНК, ДНК и АТФ | Бери-бери: утомляемость, спутанность сознания, нарушение движений, отек, сердечная недостаточность |
| B 2 (рибофлавин) | Коэнзим: способствует метаболизму глюкозы, жиров и углеводов, переносчик электронов, другие витамины группы B зависят от | Арибофлавиноз: сухая чешуйчатая кожа, воспаление и язвы во рту, боль в горле, зуд в глазах, светочувствительность |
| B 3 (ниацин) | Коэнзим: способствует метаболизму глюкозы, жиров и белков, переносчик электронов | Пеллагра: диарея, дерматит, слабоумие, смерть |
| B 5 (пантотеновая кислота) | Коэнзим: способствует метаболизму глюкозы, жиров и белков, холестерину и синтезу нейромедиаторов | Онемение и боль в мышцах, утомляемость, раздражительность |
| B 6 (пироксидин) | Коэнзим; способствует синтезу аминокислот, гликогенолизу, нейротрансмиттеру и синтезу гемоглобина | Мышечная слабость, дерматит, язвы во рту, утомляемость, спутанность сознания |
| Биотин | Коэнзим; способствует метаболизму глюкозы, жиров и белков, синтезу аминокислот | Мышечная слабость, дерматит, утомляемость, выпадение волос |
| Фолат | Коэнзим; синтез аминокислот, РНК, ДНК и синтез красных кровяных телец | Диарея, язвы во рту, спутанность сознания, анемия, дефекты нервной трубки |
| B 12 (кобаламин) | Коэнзим; катаболизм белков и жиров, функция фолиевой кислоты, синтез красных кровяных телец | Мышечная слабость, болезненность языка, анемия, поражение нервов, дефекты нервной трубки |
Придают ли добавки витамина B прилив энергии?
Хотя некоторые маркетологи заявляют, что прием витамина, который в тысячу раз превышает дневную норму некоторых витаминов группы B, повышает энергию и работоспособность, это миф, не подтвержденный наукой.«Ощущение» большей энергии от добавок, повышающих энергию, происходит из-за большого количества добавленных сахаров, кофеина и других травяных стимуляторов, которые сопровождают высокие дозы витаминов группы B. Как уже говорилось, витамины группы B необходимы для поддержки энергетического обмена и роста, но потребление большего количества, чем требуется, не дает вам больше энергии. Прекрасная аналогия этого явления — бензин в вашей машине. Едет ли быстрее с полбаком бензина или полным? Не важно; машина едет так же быстро, пока есть бензин.Точно так же истощение витаминов группы B вызовет проблемы с энергетическим обменом, но наличие большего количества витаминов, чем требуется для запуска метаболизма, не ускоряет его. Остерегайтесь покупателей добавок с витамином B; Витамины группы В не накапливаются в организме, и все излишки смываются в унитаз вместе с потраченными лишними деньгами.
витаминов группы В естественным образом присутствуют во многих продуктах питания, и многие другие продукты обогащены ими. В Соединенных Штатах дефицит витамина B встречается редко; однако в девятнадцатом веке нехватка витамина B преследовала многих людей в Северной Америке.Дефицит ниацина, также известный как пеллагра, был заметен у более бедных американцев, основным пищевым продуктом которых была рафинированная кукурузная мука (видео 6.3.1). Его симптомы были серьезными и включали диарею, дерматит, слабоумие и даже смерть. Некоторые из последствий пеллагры для здоровья являются результатом недостаточного количества ниацина для поддержания метаболических функций организма.
Видео \ (\ PageIndex {1} \) : Видео Пеллагры. Просмотрите это видео о том, как доктор Джозеф Голдбергер обнаружил, что пеллагра — это заболевание, связанное с диетой.(нажмите, чтобы посмотреть видео)
Нормы потребления и источники витаминов группы В с пищей
витаминов группы В водорастворимы и не хранятся в организме в значительных количествах. Следовательно, их необходимо постоянно получать из рациона. К счастью, витамины группы В обычно хорошо всасываются в кишечнике. Рекомендуемые диетические нормы (RDA) или адекватное потребление (AI), установленные МОМ для витаминов группы B, перечислены в таблице \ (\ PageIndex {2} \), в которой также указаны некоторые диетические источники этих питательных микроэлементов.Следует отметить, что витамины группы В теряются с продуктами во время хранения, обработки и приготовления. Чтобы максимально увеличить усвоение витамина B, фрукты и овощи не следует хранить в течение длительного времени, их следует есть больше как цельные продукты, а овощи следует готовить на пару, а не варить. Также алкоголь нарушает всасывание в кишечнике витаминов группы В. У Министерства сельского хозяйства США есть отчеты о содержании питательных веществ в пищевых продуктах, включая все витамины группы B, на их веб-сайте.
| Витамин B | RDA (мг / день) | Источники питания |
|---|---|---|
| B 1 (тиамин) | 1.2 (кобели) | Цельнозерновые, обогащенные зерна, апельсиновый сок, молоко, арахис, сушеные бобы и семена |
| 1,1 (самки) | ||
| B 2 (рибофлавин) | 1,3 (самцы) | Молоко, йогурт, обогащенные сухие завтраки, мясные субпродукты, грибы, яйца, моллюски, шпинат |
| 1,1 (самки) | ||
| B 3 (ниацин) | 16 (самцы) | Мясо, птица, рыба, цельнозерновые продукты, обогащенные хлопья для завтрака, обогащенные зерна, грибы, арахис |
| 14 (суки) | ||
| B 5 (пантотеновая кислота) | 5 (самцы) | Яйца, семечки, арахис, мясо, молоко, овощи |
| 5 (суки) * | ||
| B 6 (пироксидин) | 1.3 (кобели) | Мясо, цельнозерновые, картофель, обогащенные сухие завтраки, бананы, авокадо |
| 1,3 (самки) | ||
| Биотин | 0,03 (самцы) | Яичные желтки, арахис, сыр |
| 0,03 (самки) * | ||
| Фолиевая кислота | 0,4 (самцы) | Зеленые листовые овощи, бобовые, обогащенные хлопья для завтрака, апельсиновый сок, семена подсолнечника, печень |
| 0.4 (суки) | ||
| B 12 (кобаламин) | 0,0024 (мужчины) | Продукты животного происхождения, некоторое количество соевого молока и обогащенные хлопья для завтрака |
| 0,0024 (женщины) | ||
| * означает адекватное потребление | ||
Источник: Институт медицины. Референсные нормы потребления тиамина, рибофлавина, ниацина, витамина B 6 , фолиевой кислоты, витамина B 12 , пантотеновой кислоты, биотина и холина., 12 июня 2000 г. www.iom.edu/Reports/2000/Dietary-Reference-Intakes-for-Thiamin-Riboflavin-Niacin-Vitamin-B6-Folate-Vitamin-B12-Pantothenic-Acid-Biotin-and-Choline. aspx.
Диетические источники, богатые фолатом и витамином B
12Чтобы помочь вам получить весь витамин B 12 и фолиевую кислоту, вам необходимо поддерживать метаболизм и синтез клеток крови, посмотрите Таблицы 6.3.3 и 6.6, чтобы узнать о хороших диетических источниках этих микроэлементов.
| Продукты питания | мкг на порцию | Процент дневной нормы |
|---|---|---|
| Печень | 185 (3 унции.) | 45 |
| Печень теленка | 650 (3 унции) | 160 |
| Обогащенные сухие завтраки | 400 (¾ с.) | 100 |
| Шпинат | 100 (½ гр., Вареная) | 25 |
| Северные бобы | 100 (½ гр., Вареная) | 25 |
| Спаржа | 85 (4 пика, отварные) | 20 |
| Вегетарианская фасоль | 60 (1 ок.) | 15 |
| Брокколи | 45 (2 копья) | 10 |
| Авокадо | 45 (½ с.) | 10 |
| Хлеб (обогащенный) | 25 (1 ломтик) | 6 |
| Продукты питания | мкг на порцию | Процент дневной нормы |
|---|---|---|
| Печень | 48 (1 ломтик) | 800 |
| Зажимы | 34.2 (3 унции) | 570 |
| Органическая телячья печень | 31 (3 унции) | 520 |
| Обогащенные сухие завтраки | 6,0 (1 порция) | 100 |
| Форель (дикая) | 5,4 (3 унции) | 90 |
| Форель (разводимая) | 3,5 (3 унции.) | 58 |
| Лосось (нерка) | 4,8 (3 унции) | 80 |
| Чизбургер | 2,1 | 35 |
| Йогурт (простой) | 1,4 (1 с.) | 23 |
| Говядина (верхнее филе) | 1,4 (3 унции) | 23 |
| Молоко | 0.9 (1 в.) | 15 |
| Яйцо | 0,6 (1 большой) | 10 |
Витамин К: функции метаболизма
Появляются доказательства того, что витамин К может играть роль в энергетическом обмене, но в настоящее время точные функции витамин К-зависимых ферментов в энергетическом обмене остаются неуловимыми. Витамин К необходим для оптимального метаболизма костей.Витамин К также важен для функции крови.
Дефицит витамина К вызывает нарушения свертываемости крови. Это относительно редко, но люди с заболеваниями печени или поджелудочной железы, глютеновой болезнью или нарушением всасывания подвергаются более высокому риску дефицита витамина К. Признаки и симптомы включают носовое кровотечение, легкие синяки, сломанные кровеносные сосуды, кровоточивость десен и обильные менструальные кровотечения у женщин. Функцию антикоагулянта варфарина нарушает избыточное потребление витамина К из добавок.Кальций также играет роль в активации белков свертывания крови, как обсуждалось в предыдущей главе.
Нормы потребления и источники витамина К с пищей
Рекомендуемая суточная норма витамина К для взрослых мужчин составляет 120 мкг / день, а для взрослых женщин — 90 мкг / день. Витамин К присутствует во многих продуктах питания, и его наибольшая концентрация содержится в зеленых листовых овощах. В Таблице 10.4.5 приведен список пищевых источников витамина К.
.| Продукты питания | мкг на порцию | Процент дневной нормы |
|---|---|---|
| Брокколи (½ гр.) | 160,0 | 133 |
| Спаржа (4 копья) | 34,0 | 28 |
| Капуста (½ ц.) | 56,0 | 47 |
| Шпинат (½ ц.) | 27,0 | 23 |
| Горошек зеленый (½ ц.) | 16,0 | 13 |
| Сыр (1 унция.) | 10,0 | 8 |
| Ветчина (3 унции) | 13,0 | 11 |
| Говяжий фарш (3 унции) | 6,0 | 5 |
| Хлеб | 1,1 | <1 |
| Оранжевый | 1,3 | 1 |
Основные выводы
- Витамины и минералы играют различную роль в энергетическом обмене; они необходимы как функциональные части ферментов, участвующих в высвобождении и хранении энергии.
- Водорастворимые витамины группы B участвуют в качестве коферментов в расщеплении питательных веществ и в построении макромолекул, таких как белок, РНК и ДНК.
- Дефицит витамина B относительно редок, особенно в развитых странах; хотя последствия для здоровья могут быть серьезными, например, при дефиците фолиевой кислоты во время беременности и повышенном риске дефектов нервной трубки у потомства.
Обсуждение стартеров
- Обсудите взаимозависимость действия витаминов группы В; это означает, что уровень одного влияет на функцию другого.
- Проведите в классе дискуссию о том, разумно ли со стороны федерального правительства регулировать требования к добавкам с питательными микроэлементами, например, содержащим витамин B.
Метаболизм зависит от ферментов — синдром Вильсона
На все физические и даже умственные процессы, происходящие в нашем теле, влияют химические реакции. Метаболизм — это, по сути, сумма всех химических реакций, происходящих в организме.Люди используют термин скорость метаболизма, чтобы обозначить скорость, с которой происходят эти химические реакции.
Правильная функция обмена веществ в значительной степени зависит от правильного функционирования ферментов организма. Ферменты — это белки, которые катализируют (они же катализаторы) практически все химические реакции в организме. Катализаторы позволяют некоторым химическим реакциям протекать так, как иначе бы они не происходили. Например, когда кто-то использует эпоксидный клей для ремонта, необходимо смешать две части вместе, прежде чем его можно будет использовать.Одна часть — это смола, а другая — отвердитель. По отдельности в качестве клея они бесполезны, потому что не затвердевают. Но когда в смолу добавляется отвердитель, происходит химическая реакция, позволяющая клею делать то, для чего он предназначен. Здесь отвердитель действует как жизненно важный катализатор. Точно так же, если бы не важные катализаторы организма, известные как ферменты, организм не смог бы выжить.
Итак, в основе метаболизма лежат ферменты. Оказывается, насколько хорошо фермент функционирует, зависит от его формы, а форма зависит от его структуры.Ферменты — это длинные цепочки аминокислот, которые собраны в соответствии с генетическим кодом ДНК. В зависимости от того, как расположены аминокислоты ферментов, ферменты принимают определенную форму. Эта форма может меняться в разное время и в разных условиях. Фактически, именно эта способность изменять форму характерным образом и дает им их функцию. Они похожи на скрученный шнур телефонной трубки, который раскручивается, когда вы отвечаете на звонок и туго натягиваете шнур, а затем снова скручивается в прежнюю форму, когда вы снова кладете его на крючок.
Скорость метаболизма и способы ее измерения — Скорость метаболизма — Высшая версия биологии
Скорость обмена веществ в организме — это количество энергии, расходуемое этим организмом в определенный период времени — обычно ежедневно.
В состоянии покоя, то есть в периоды бездействия, скорость метаболизма известна как базовая скорость метаболизма (BMR). В состоянии покоя BMR низок по сравнению с тем, когда организм выполняет такие действия, как упражнения.
Во время этого состояния покоя организму необходимо использовать энергию только для поддержания нормальной работы жизненно важных органов, таких как сердце, легкие и мозг.
Скорость метаболизма можно измерить разными способами:
- Потребление кислорода
- Производство углекислого газа
- Производство тепла
Как правило, чем больше масса организма, тем выше скорость метаболизма этого организма.
Организмам с высоким уровнем метаболизма требуется более эффективная доставка кислорода к клеткам.
Однако BMR выше на единицу массы тела у мелких животных по сравнению с более крупными. Это связано с тем, что более высокая скорость метаболизма мелких животных требует большей доставки кислорода к тканям вокруг тела.
Кроме того, более мелкие животные имеют большее отношение площади поверхности к объему, поэтому теряется больше тепла.
Скорость метаболизма организмов может быть измерена с помощью нескольких различных устройств, включая респираторы, кислородные датчики, датчики углекислого газа и калориметры.
Количество активных метаболических путей ограничено количеством клеточных ограничений при максимальной скорости метаболизма
Цитата: de Groot DH, van Boxtel C, Planqué R, Bruggeman FJ, Teusink B (2019) Количество активных метаболических пути ограничены количеством клеточных ограничений при максимальной скорости метаболизма.PLoS Comput Biol 15 (3): e1006858. https://doi.org/10.1371/journal.pcbi.1006858
Редактор: Василий Хациманикатис, Федеральная политехническая школа Лозанны, ШВЕЙЦАРИЯ
Поступила: 03.10.2018; Принята к печати: 7 февраля 2019 г .; Опубликовано: 11 марта 2019 г.
Авторские права: © 2019 de Groot et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные находятся в рукописи и ее файлах с вспомогательной информацией.
Финансирование: DHdG и BT были поддержаны грантом NWO VICI 865.14.005 (https://www.nwo.nl/) и проектом Era-Industrial Biotechnology nr. 053.80.772 (http://www.era-ib.net/ecoyeast). CvB был поддержан грантом NWO STAR 022.005.031 (www.nwo.nl). Спонсоры не играли никакой роли в дизайне исследования, сборе и анализе данных, решении опубликовать или подготовке рукописи.
Конкурирующие интересы: Авторы заявили, что конкурирующих интересов не существует.
Введение
Микроорганизмы Fitter вымирают конкурентов, синтезируя более жизнеспособное потомство [1, 2]. Скорость синтеза клеток потомства на клетку, то есть удельная скорость роста, является общим определяющим фактором эволюционного успеха среди видов микробов [1]. Высокая скорость роста требует высоких скоростей метаболизма, что, в свою очередь, требует высоких концентраций ферментов [3].Из-за ограниченных биосинтетических ресурсов, таких как рибосомы, полимеразы, энергия и питательные вещества, экспрессия любого фермента происходит за счет других [4, 5]. Следовательно, селективное давление, направленное на максимальную скорость роста, требует, чтобы преимущества и затраты всех ферментов были должным образом сбалансированы, что привело к оптимально настроенной экспрессии ферментов [6–9].
Настройка уровней экспрессии всех ферментов представляется очень сложной задачей. Во-первых, геном микроорганизма кодирует тысячи реакций с ассоциированными ферментами.Во-вторых, изменение уровня экспрессии одного фермента не только влияет на скорость связанной с ним реакции, но также меняет внутриклеточные концентрации метаболитов. Эти концентрации метаболитов влияют на активность многих других ферментов нелинейным образом. Таким образом, с математической точки зрения, микроорганизмы должны решать многомерную задачу нелинейной оптимизации.
Удивительно, но эксперименты на множестве разных микроорганизмов часто показывают простые линейные зависимости между скоростью роста, уровнями экспрессии ферментов и скоростью метаболизма [10–12], и данные часто можно описать с помощью крупнозернистых линейных моделей.Это говорит о том, что микроорганизмы на самом деле используют лишь несколько регуляторных степеней свободы для настройки метаболического потока и экспрессии белков. В настоящее время неясно, почему это простое низкоразмерное поведение является результатом чрезвычайно сложной задачи настройки априори. Учитывая, что тенденция к простоте широко распространена среди микроорганизмов, мы ожидали, что это связано с общим эволюционным принципом.
Мы обнаружили эволюционный принцип экстремума: максимизация скорости роста приводит микроорганизмы к минимальной метаболической сложности.Мы приводим математическое доказательство этого принципа в разделе «Методы». Он выводится из основных принципов, а именно (i) сохранения массы, т. Е. Отношений стационарной реакции и стехиометрии, и (ii) биохимии фермента, т. Е. Линейной зависимости активности фермента от количества фермента и ее нелинейной зависимости. от концентрации субстрата и продукта. Наши результаты открывают новый взгляд на регуляцию метаболизма, сложность которого определяется не размером сети или уравнениями скорости, а ограничениями, действующими на концентрации ферментов.
Методы
В этом разделе мы представим класс моделей, которые мы изучали, и математически докажем наш главный результат: принцип экстремума. Читателям, которые хотели бы пропустить математическое доказательство, настоятельно рекомендуется прочитать биологическое резюме результатов в конце раздела.
Модель: Максимизация скорости эволюции может быть изучена только в кинетической модели метаболизма с ограничениями на концентрацию ферментов
Структура любой метаболической сети может быть задана стехиометрической матрицей N , указывающей, какие метаболиты (строки) потребляются или производятся в каждой реакции (столбцы).Поскольку мы можем разделить обратимые реакции на две необратимые реакции [13], с этого момента мы будем предполагать, что все реакции необратимы. Распределение стационарного потока затем задается вектором скоростей реакций v таким образом, чтобы не было накопления или истощения метаболитов и чтобы удовлетворялись все ограничения необратимости. Растворы вместе образуют конус потока: (1) где r — количество реакций. В установившемся режиме мы максимизируем целевой поток, который представляет собой (линейную комбинацию) компонент (компоненты) этого вектора потока.Часто целью выбирается общая реакция клеточного синтеза, также называемая реакцией биомассы v BM , которая делает все клеточные компоненты в правильных пропорциях в соответствии с составом биомассы [14].
Чтобы понять распределение ресурсов, связанных с определенной метаболической активностью, нам необходимо знать соотношение между скоростью реакций, катализируемых ферментами, и концентрацией ферментов. При постоянных концентрациях метаболитов они, как правило, пропорциональны [3], что фиксируется уравнением скорости: (2) где e i — концентрация фермента, катализирующего эту реакцию, k cat, i — его максимальная каталитическая скорость и f i ( x ) это «функция насыщения» фермента, которая зависит от концентрации метаболитов x .Эта функция, f i ( x ), часто является нелинейной, включает термодинамическую движущую силу, (аллостерическую) активацию или ингибирование и другие эффекты, специфичные для ферментов.
Чтобы смоделировать максимизацию потока клеточного синтеза, мы должны учитывать ограничения концентраций ферментов, возникающие, например, из-за ограниченной способности клеточных компартментов к растворителю или из-за ограниченной способности к синтезу рибосомных белков. Мы моделируем эти биофизические пределы, налагая K ограничений, каждое из которых моделируется взвешенной суммой концентраций ферментов: Эти ограничения соответствуют ограниченным пулам ферментов.Следовательно, сверхэкспрессия одного фермента происходит за счет других, на которые распространяются те же биофизические ограничения. Веса, определяют долю, которую использует один моль / литр фермента i th по сравнению с ограниченным пулом ферментов j th . Например, для ограничения, описывающего ограниченную растворяющую способность мембраны, вес фермента представляет собой долю доступной площади мембраны, которая используется этим ферментом; поэтому этот вес отличен от нуля только для мембранных белков.Мы называем ограничение «активным», когда оно ограничивает скорость роста клетки, указывая на то, что соответствующий пул ферментов используется полностью. Один фермент может принадлежать к одному, нескольким или ни одному из этих ограниченных пулов.
Обратите внимание, что эти ограничения на концентрацию ферментов отличаются от ограничений на скорости реакции, которые часто используются в стехиометрических методах (например, посредством анализа баланса потока). Для этих линейных моделей известно — подобно тому, что мы выведем в общем нелинейном случае в этой работе, — что несколько минимальных путей составляют оптимальные решения в таких моделях [15].Однако ограничения на скорость реакции не отражают способность микроорганизмов регулировать содержание фермента: любое ограничение скорости реакции в принципе можно преодолеть путем увеличения концентрации соответствующего фермента. Ограничения ферментов, которые мы моделируем, обусловлены биофизическими законами и, следовательно, не могут быть устранены регуляцией метаболизма. Таким образом, они должны быть исследованы для изучения эволюции метаболизма, хотя это вынуждает нас включить в нашу теорию сложные (и часто неизвестные) функции насыщения ферментами, f i ( x ).
Количество ограничений и точное значение веса может варьироваться в зависимости от организма. В целом мы ожидаем, что это число будет низким, и действительно, в литературе предложено не так много различных ограничений экспрессии ферментов. Многие аспекты микробного роста были успешно описаны с использованием ограничений, которые являются (или могут быть переформулированы как) ограничениями экспрессии ферментов, такими как ограниченная скорость реакции и ограниченная емкость растворителя в клеточных компартментах [4, 5, 10, 16–20].
Введение кинетики фермента в уравнение (2) позволяет нам переписать ограничения фермента как: (3) Мы отмечаем, что, хотя и записаны в терминах потоков, эти ограничения не эквивалентны нормальным ограничениям потока, используемым в FBA, поскольку взвешенные суммы теперь зависят от концентраций метаболитов.Чтобы максимизировать поток клеточного синтеза, следует оптимизировать не только концентрации ферментов, но и концентрации внутриклеточных метаболитов. Из-за необходимого учета кинетики ферментов максимизация потока превращается в сложную нелинейную задачу. Это проблема, которую мы исследовали. Примечательно, что ниже мы докажем, что решение все еще использует только несколько минимальных метаболических путей.
Минимальные строительные блоки: режимы элементарного потока
Минимальный метаболический путь называется «режимом элементарного потока» (EFM).Другими словами, EFM — это подсети с минимальной поддержкой, которые могут поддерживать устойчивое состояние [21]. «Опорой» вектора потока является набор участвующих реакций: R ( v ) = { j : v j ≠ 0}. То, что EFM, EFM , является минимально поддерживаемым, означает, что при наличии другого вектора потока, такого как R ( v ′) ⊆ R ( EFM ), мы должны иметь v ′ = α EFM для некоторых α ≥ 0.Другими словами, ни одна из используемых реакций не может быть установлена на ноль в EFM без нарушения условия устойчивого состояния. Эти метаболические подсети полностью определяются стехиометрией реакции, поэтому для их идентификации не требуется кинетическая информация. Однако из-за множества комбинаций параллельных альтернативных метаболических маршрутов в метаболических сетях в настоящее время вычислительно невозможно найти полный набор EFMs в сети масштаба генома [22, 23].
Мы используем EFM, потому что любое установившееся распределение потока можно разложить на положительные линейные комбинации EFM. Действительно, Гагнёр и Кламт показали, что в любой метаболической сети, в которой обратимые реакции расщепляются на две необратимые, EFMs совпадают с крайними лучами заостренного многогранного конуса [13]. Таким образом, мы можем написать: (4) где коэффициенты умножения λ i обозначают, сколько EFM i th используется, а F обозначает общее количество EFM в сети.Уравнение (4) показывает, что EFM являются основными строительными блоками устойчивого метаболизма. Обратите внимание, что, хотя режимы элементарного потока являются постоянными векторами, определяемыми стехиометрией, λ i -факторы изменчивы и зависят от концентраций метаболитов. Мы уточним эту зависимость в приложении S1, раздел 5.
EFM определены с точностью до константы: если v — EFM, то αv для любого. Это имеет два важных следствия.Во-первых, соотношение между входами потока в EFM фиксировано, а во-вторых, мы можем масштабировать одну запись EFM до 1. Мы рассмотрим оптимизацию некоторого объективного потока v r в установившемся состоянии. Следовательно, нам нужно учитывать только те EFM, которые имеют ненулевое значение потока r th , потому что мы предполагаем, что все EFM (даже те, которые не производят объективный поток) будут использовать некоторые из ограниченных пулов ферментов. . Таким образом, мы не будем рассматривать патологический случай, когда есть EFM, который не создает объективного потока, но также не несет никаких затрат, поскольку этот EFM всегда можно добавить к оптимальному решению.Без ограничения общности, мы делаем целевой поток последней записью в векторе потока, и мы всегда будем масштабировать эту запись до 1. Таким образом, EFM i th может быть обозначен как, причем все они однозначно определяются стехиометрией. Фактор λ i в (4) теперь может быть интерпретирован как поток, который EFM i вносит в объективный поток.
Используя EFM, мы можем однозначно количественно оценить метаболическую сложность как количество несущих поток элементарных режимов потока.Мы называем EFM минимальной единицей метаболической сложности, потому что значения потока через участвующие в ней реакции могут масштабироваться только с одним общим фактором. Распределение потока, которое представляет собой сумму K EFM, таким образом, имеет K степеней свободы потока. Небольшое количество степеней свободы приводит к метаболическому поведению с простыми соотношениями между скоростью роста и величиной потока.
Векторы затрат: низкоразмерный взгляд на метаболизм
Учитывая K ограничений, мы можем для каждого EFM рассчитать стоимость на ограничение для создания одного единичного целевого потока.Эти затраты K содержат всю необходимую информацию для оптимизации темпов роста. Поэтому здесь мы определим векторов затрат , которые имеют эти затраты в качестве своих записей. Мы будем использовать векторы затрат для изучения метаболизма в ограниченном пространстве малой размерности на протяжении всей статьи.
Как обсуждалось выше, мы можем масштабировать каждый EFM так, чтобы он был вектором формы. Таким образом, чтобы получить единичный поток от объектива, нам потребуется поток сквозной реакции Дж .Так как у нас v j = k cat, j e j f j ( x ), получаем где обозначает необходимую концентрацию фермента j на одну единицу потока объектива через EFM i . Затем мы можем определить вектор затрат d i ( x ) для EFM i th , с компонентами, заданными общими затратами, которые этот EFM приносит на ограничение: (5) Поскольку кинетика фермента определяет концентрацию фермента и, следовательно, затраты на фермент, маловероятно, что несколько EFM имеют точно такие же затраты.В разных EFM используется по крайней мере один другой фермент, и маловероятно, чтобы необходимые концентрации этих разных ферментов были точно такими же действительными числами. Если один из этих неперекрывающихся ферментов является частью ограниченного пула, EFM, таким образом, будут иметь разные затраты. Если, однако, ни один из неперекрывающихся ферментов не является частью ограниченного пула, несколько EFM действительно могут иметь одинаковые затраты. Чтобы разобраться в этом случае, мы вводим понятие эквивалентов EFM . В методах моделирования, которые не включают кинетическую информацию, таких как FBA, наличие эквивалентных EFM гораздо более вероятно, так что пространства решений часто являются многомерными подпространствами.
Определение 1 . Учитывая набор ограничений « , два EFM , EFM 1 , EFM 2 , называются эквивалентом относительно ограничений , если их связанные векторы затрат равны : d 1 ( x ) = d 2 ( x ).
Поскольку векторы затрат играют центральную роль во всей статье, мы проиллюстрировали их определение и использование на рис. 1.Многие из наших результатов были получены в результате изучения этих векторов затрат.
Рис. 1. Формализм вектора стоимости показывает, что определяет количество EFM в оптимальном решении.
Здесь мы рассматриваем упрощенную модель с двумя EFM (синим и оранжевым) и двумя ограничениями. В действительности, необходимо сравнивать затраты на гораздо большее количество EFM, а также, возможно, на большее количество ограничений. Вектор затрат EFM i th обозначает доли первого и второго ограниченного пула ферментов, которые этот EFM использует при создании одной единицы потока цели.Поток клеточного синтеза, производимый EFM i , обозначается λ i , а соответствующие затраты на фермент составляют λ i d i ( x ) . Стоимость смешивания EFM 1 и 2 соответствует взвешенной сумме векторов затрат: λ 1 d 1 ( x ) + λ 2 d 2 ( х ).Смешивание возможно до тех пор, пока не будет превышено ни одно из ограничений: λ 1 d 1 ( x ) + λ 2 d 2 ( x ) ≤ 1. Целевое значение, λ 1 + λ 2 , максимизируется путем подбора векторной суммы как можно большего числа векторов в блоке ограничений. Это решение показано штриховыми векторами. Чистое использование одного EFM с недиагональным вектором затрат приводит к недоиспользованию одного ограничения, в то время как диагональные векторы затрат могут исчерпать оба ограниченных пула.Смесь EFM всегда будет комбинацией вектора над диагональю и вектора под диагональю. Все EFM и их смеси могут быть ранжированы точкой на диагонали, которая обозначает среднюю стоимость потока синтеза элементарной ячейки (см. Лемму 4 в Приложении S1 для доказательства). Чистое использование векторов затрат над диагональю оценивается путем проецирования вектора затрат по горизонтали на диагональ, в то время как чистое использование векторов затрат под диагональю оценивается с помощью вертикальной проекции. Смеси ранжируются путем размещения точки на пересечении диагонали с линией между двумя векторами затрат.(Смесь) EFM (ов) с самой низкой средней стоимостью (то есть с точкой, ближайшей к исходной точке) приводит к наивысшей скорости роста (математическое доказательство включено в Приложение S1). Ферментативные затраты на EFM зависят от внутриклеточных концентраций метаболитов, то есть от насыщения ферментов. Заштрихованные области указывают альтернативные положения векторов стоимости при различных внутриклеточных концентрациях метаболитов, показаны два из них. Синий и оранжевый векторы затрат приводят к наивысшим темпам роста при использовании только этого EFM.Мы видим, что на левом рисунке оранжевый EFM дает более высокую скорость роста. При изменении условий окружающей среды векторы затрат могут измениться, и смесь EFM может стать лучше любой из EFM (правый рисунок). Подобное изменение привело бы к изменению метаболического поведения.
https://doi.org/10.1371/journal.pcbi.1006858.g001
Принцип экстремума: количество активных EFM определяется количеством ограничений на экспрессию фермента
Здесь мы доказываем основной результат этого исследования — принцип экстремума.Для общей метаболической модели, как было введено выше, она устанавливает необходимое условие для того, чтобы вектор потока был максимизатором целевого потока.
Теорема 1 . Рассмотрим метаболическую сеть, характеризуемую стехиометрической матрицей N . Пусть v r будет объективным потоком, который должен быть максимизирован в установившемся состоянии при K линейных ферментативных ограничениях формы : Затем в оптимальном решении используется не более K неэквивалентных режимов элементарного потока.
Доказательство . Мы предположили, что v j ≥ 0 для всех реакций в сети, поскольку, без ограничения общности, мы разделили все обратимые реакции на прямую и обратную [13]. Давайте пока также предположим, что ни одна из EFM не эквивалентна (где эквивалентность определена в соответствии с определением 1), мы рассмотрим случай с эквивалентными EFM в конце доказательства.
Согласно уравнению (4), оптимальное решение всегда может быть выражено в виде конической комбинации EFM.Как и раньше, мы масштабируем каждый EFM так, чтобы он был вектором формы. Объективный поток для вектора потока v теперь можно записать как (6) где M — количество EFM, содержащих ненулевое значение v r . Поскольку EFM являются фиксированными векторами, λ i становятся нашими переменными оптимизации. Поскольку у нас есть Это позволяет нам переписать ограничение фермента как (7) На последнем этапе мы распознали компоненты вектора затрат, определенные в уравнении (5).
Запись k th вектора затрат i обозначает стоимость ферментов в ограничении k ( k — ферментов ) для получения одной единицы потока объектива через EFM i (и, следовательно, стоимость ферментов для увеличения этого потока в какой-то раз). Мы можем переписать нашу задачу оптимизации в терминах этих векторов затрат. Мы будем обозначать каждую концентрацию метаболита как внешнюю, x E , или внутреннюю, x I , так что: x = ( x E , x I ).Это различие важно, потому что внешние концентрации задаются окружающей средой и, следовательно, являются частью параметров задачи оптимизации, в то время как внутренние концентрации могут регулироваться ячейкой и, следовательно, являются частью решения. Нам нужно решить (8) и используя уравнения (6) и (7), это эквивалентно (9) где D = [ d 1 ( x ) ⋯ d r ( x ) — вектор стоимости].Соотношение D ( x ) ⋅ λ ≤ 1 показывает, что оптимальный вектор λ действительно зависит от концентраций метаболитов x , как показано ниже в уравнении (4).
Во многом следуя Wortel et al. [24], теперь мы используем тонкий математический аргумент. Мы фиксируем x = x 0 , чтобы ферментные насыщения f j ( x 0 ) были постоянными.Это даст нам вектор фиксированных затрат для каждого EFM. Оставшаяся проблема оптимизации затем визуализируется на рис. 1, где векторы затрат некоторых EFM нанесены на график в рамке ограничений. Нахождение оптимального решения эквивалентно нахождению суммы скалярных кратных векторов стоимости без выхода за рамки ограничений при максимизации суммы этих кратностей. Пример на рис. 1 показывает только 2 ограничения, но в целом у нас будет M векторов в K -мерном кубе.
В общем случае может показаться интуитивно понятным, что ограничения K приводят к использованию максимум K EFM, поскольку все K линейно-независимых векторов образуют основу K -мерного пространства. Таким образом, мы всегда можем взять комбинацию из K векторов, чтобы достичь точки, в которой все ограничения выполняются с равенством. Однако нам следует быть осторожными, потому что мы можем получить отрицательные λ для некоторых EFM. Продолжим доказательство, переписав задачу в форме линейного программирования (LP),
- Maximize v r = 1 ⋅ λ ,
в зависимости от
A λ ≤ z ,
где Решения этой задачи линейного программирования образуют многогранник в, ограниченный гиперповерхностями, заданными ограничениями.Важнейшая теорема LP учит нас, что оптимальное решение находится среди вершин этого многогранника. Размерность таких вершин равна нулю, что означает, что оптимальные решения удовлетворяют как минимум M из K + M ограничений с равенством. Следовательно, самое большее ( K + M ) — M = K ограничений могут быть удовлетворены при строгом неравенстве. Эти неравенства K могут быть сконцентрированы в части λ i ≥ 0, что означает, что используются соответствующие режимы элементарного потока K .Таким образом, оптимальное решение может использовать не больше EFM, чем имеется активных ограничений в системе, тем самым доказывая теорему для любого произвольного вектора концентраций метаболитов x .
Есть одно возможное исключение из приведенного выше рассуждения. Допустим, ЭФМ К используются в оптимальном:. Если один EFM, скажем, EFM K , имеет эквивалентный EFM, скажем EFM K +1 , то мы можем заменить использование EFM K любой выпуклой комбинацией EFM K и K + 1 и решение все равно будет оптимальным.Таким образом, в случае, если стоимость нескольких EFM одинакова, оптимальный вектор потока может состоять из большего количества EFM, чем количество ограничений. Вот почему теорема говорит нам только о том, что в оптимальном решении используется не более тыс. Неэквивалентных EFM.
Наконец, отсюда следует, что, поскольку теорема верна для любого набора концентраций метаболитов x , она, конечно же, верна и для оптимального набора ,.
Отметим, что оптимальные внутренние концентрации, выбор EFM и, следовательно, оптимальные концентрации ферментов зависят от внешних концентраций x E .Таким образом, не следует напрямую из теоремы, какие именно EFM являются оптимальными.
Мы думаем, что случай, когда несколько EFM эквивалентны, не очень распространен в биологии. Во-первых, ограничения на экспрессию ферментов связаны с биофизическими ограничениями, и мы ожидаем, что они будут действовать одновременно на многие ферменты. Это снижает вероятность наличия нескольких EFM, которые используют одни и те же ферменты в ограниченном пуле ферментов. Во-вторых, даже если несколько EFM будут использовать одни и те же ферменты, тогда стоимость ферментов зависит от насыщения ферментов, а это зависит от оптимальных концентраций метаболитов.Эти оптимальные концентрации зависят от остального метаболизма, так что неперекрывающаяся часть EFM все еще может влиять на стоимость ферментов. По этим двум причинам в оставшейся части этой работы мы будем предполагать, что EFM, как правило, не эквивалентны.
Ранее опубликованная теорема о том, что максимальный удельный поток достигается в EFM [24, 25], является частным случаем теоремы 1. В формализме вектора стоимости, который мы описали на рис. 1, она визуализируется векторами стоимости на линии а не в коробке, потому что есть только одно ферментативное ограничение (общая концентрация фермента ограничена).В этом случае действительно существует кратчайший вектор затрат для всех, кроме незначительного подмножества ситуаций (как обсуждается в доказательстве).
Следующее следствие можно использовать, чтобы узнать, сколько ограничений активно, когда мы наблюдаем определенное количество активных EFM. Он противоположен теореме 1 и поэтому математически эквивалентен. Причина, по которой это излагается отдельно, заключается в различии в биологической применимости: теорема является предсказательным утверждением, а следствие носит описательный характер.Как мы увидим в разделе «Результаты», теорема говорит нам, что метаболическая сложность невысока, поскольку количество ферментативных ограничений обычно невелико. Следствие, однако, позволяет нам сделать вывод из экспериментальных данных, сколько ограничений должно быть активным, и, таким образом, дает нам физиологическое понимание на основе данных на уровне популяции.
Следствие 2 . Если поток v r оптимизирован и используются K неэквивалентных элементарных режимов потока, то должно быть активным минимум K линейных ферментативных ограничений .
EFM— не единственный набор строительных блоков, который мы могли бы использовать. В контексте анализа баланса потока максимизация скорости на основе ограничений может быть изучена путем вычисления элементарных векторов потока (EFV) [26, 27], которые являются минимальными путями, которые генерируют все распределения потоков, которые удовлетворяют не только предположению о стационарном состоянии, но и дополнительные ограничения. Следовательно, для фиксированных значений насыщения и ограничений ферментов EFV обеспечивают набор возможных строительных блоков, выпуклые комбинации которых автоматически удовлетворяют всем ограничениям.Однако, поскольку каждый EFV представляет собой коническую комбинацию EFM, и поскольку мы хотели изучить максимизацию эволюционной скорости роста, мы предпочли провести анализ на множестве EFM. Это связано с тем, что EFM предоставляют набор инвариантных (по крайней мере, в масштабах времени, в которых не развивается стехиометрия) объектов, для которых могут быть созданы регуляторные схемы. В принципе, принцип экстремума также можно записать в терминах EFV. Мы можем показать, аналогично тому, как это было в доказательстве выше, что решения с максимальной скоростью будут использовать только один EFV, который представляет собой выпуклую комбинацию максимум K EFM.
Биологическое изложение принципа экстремума и его доказательство
Принцип экстремума, изложенный в теореме 1, является утверждением обо всех метаболических сетях, независимо от размера сети, топологии или кинетики конкретного фермента. Все микроорганизмы подвергаются небольшому количеству ферментативных ограничений, и все метаболические сети имеют режимы элементарного потока в качестве строительных блоков: минимальные пути, которые производят все клеточные компоненты из внешних источников. Потоки через участвующие реакции в EFM можно масштабировать только с одним общим коэффициентом.Мы пришли к выводу, что использование дополнительного EFM, таким образом, добавляет только одну степень свободы потока, так что экспериментальные данные покажут низкую сложность, если используется несколько EFM. Затем мы доказали принцип экстремума, заявив, что количество несущих поток EFM в растворе с максимальной скоростью роста всегда ограничено количеством ограничений на экспрессию фермента. В целом это приводит к предсказанию того, что микробное поведение будет иметь низкую сложность.
В доказательстве мы сравнили затраты и преимущества различных EFM.Чтобы быть точным, мы изменили масштаб EFM таким образом, чтобы выгода от каждого EFM была равной: все они дают одну единицу объективного потока. Если у нас есть K ограничений, у нас также есть K различных затрат, для которых нам нужно сравнить разные EFM. Мы показали, что оптимальным решением является комбинация до тыс. таких ЭФМ. Это соответствует интуиции, согласно которой для каждого ограничения можно выбрать один EFM, поскольку он имеет низкую стоимость по сравнению с этим ограничением.
Чтобы найти доказательство, мы разработали структуру с использованием векторов стоимости.На рис. 1 мы суммируем, как эта структура позволяет нам изучать метаболизм высокой размерности в нескольких измерениях, которые действительно имеют значение: мы можем сравнить стоимость ферментов всех EFM в «ограниченном пространстве» низкой размерности, определяемом ограниченными пулами ферментов. Эта перспектива позволяет нам разрабатывать эксперименты, которые характеризуют активные биофизические ограничения, как мы обсудим в разделе «Результаты».
Результаты
Метаболическая сложность обычно очень низкая
Мы назвали EFM минимальной единицей метаболической сложности, потому что соотношения между потоками через все участвующие реакции фиксированы, и ни одна из его реакций не может быть устранена.Следовательно, микроорганизм, использующий один EFM, может изменить скорость всех реакций только с одним и тем же фактором. Другими словами, существует только одна регулирующая степень свободы вместо многих, если бы все скорости реакции можно было настраивать отдельно. В этом случае значения магнитного потока можно описать только одной прямой линией. Это усложняется, когда увеличивается количество переносящих поток (активных) ЭСМ. Используя эти знания, можно оценить количество активных EFM по измерениям потока.
Мы повторно проанализировали данные хемостатов с ограничением содержания углерода и действительно обнаружили, что скорость поглощения глюкозы и кислорода может быть описана прямой линией для большого диапазона темпов роста, что свидетельствует об использовании однократного EFM (рис. 2, S1, раздел 8 приложения). .Однако возможность, которую мы не можем исключить, заключается в том, что используется много EFM, но все эти EFM имеют одинаковую связь между скоростью роста, потреблением глюкозы и потреблением кислорода. С другой стороны, экспериментально измеренные законы линейного роста между клеточными строительными блоками и ростом [11, 12, 18], а также успех крупнозернистых моделей [4, 5] действительно предоставляют некоторые дополнительные указания на использование малых количество ЭФМ. Более определенное доказательство можно найти двумя способами. Во-первых, если при сбалансированном росте в немного разных средах измеряется множество различных скоростей реакций, или, во-вторых, если измеряются все внутренние потоки в клетке и доступно полное знание стехиометрической сети.Однако, насколько нам известно, доступные в настоящее время наборы данных флуксома были собраны по мутантам или в очень разных средах выращивания, что делает их непригодными для наших целей. На данный момент, основываясь на имеющихся данных, мы осторожно утверждаем, что количество одновременно активных EFM обычно очень мало, порядка от 1 до 3. То, что микроорганизмы выберут только несколько EFM из миллиардов альтернатив, соответствует наш принцип экстремума, теорема 1. Эти альтернативы, по-видимому, эволюционно не эквивалентны, и было выбрано лишь небольшое их количество из-за их превосходной кинетики.
Рис. 2. Пропорциональность скорости реакции и скорости роста, демонстрируемая многими микроорганизмами, свидетельствует о низкой метаболической сложности.
Измеренные скорости поглощения [28–33] были получены из экспериментов, в которых скорость роста варьировалась в хемостатах с ограничением углерода. Для каждого вида мы нормализовали измеренную скорость роста до так называемой критической скорости роста: скорости роста, при которой начинается производство продуктов перелива. Скорости поглощения были нормализованы относительно скорости поглощения видов при критической скорости роста.До критической скорости роста все микроорганизмы демонстрируют простую пропорциональную зависимость между скоростью роста и скоростью поглощения глюкозы и кислорода. В Разделе 8 мы объясняем, почему эта пропорциональность указывает на использование только одного EFM. После критической скорости роста скорости реакции перестают быть пропорциональными, это явление называется метаболизмом переполнения.
https://doi.org/10.1371/journal.pcbi.1006858.g002
Принцип экстремума: небольшое количество биофизических ограничений обуславливает низкую метаболическую сложность
Принцип экстремума гласит: когда скорость конкретной реакции в метаболической сети максимальна, количество несущих поток EFM не более чем равно количеству ограничений на концентрации ферментов, которые ограничивают целевой поток.В частности, принцип справедлив для реакции клеточного синтеза. Следовательно, если количество активных ограничений невелико, то будет и количество активных EFM с максимальной скоростью роста. Это основа нашего открытия, что максимальная скорость роста требует минимальной метаболической сложности, и это расширяет результат, согласно которому скорость максимизируется одним EFM при общем ограничении белка [24, 25]. Этот более ранний результат не мог объяснить — с точки зрения распределения ресурсов — наборы данных, в которых используются несколько метаболических путей, такие как метаболизм переполнения, метаболические переключатели и экспрессия неиспользованных белков.
Принцип экстремума сохраняется независимо от сложности метаболической сети, то есть ее кинетики и структуры. Сложность метаболизма определяется только количеством активных ограничений; кинетика и структура впоследствии определяют, какие EFM являются оптимальными и выбираются эволюцией — как показано на примере in silico эволюция метаболической регуляции в направлении только одного активного EFM [34]. По этой причине также метаболические модели в масштабе генома, которые содержат все аннотированные метаболические реакции, которые кодирует геном микроорганизма [35], и даже те, которые были изучены с различными ограничениями дополнительных ресурсов [36, 37], ведут себя качественно так же, как и упрощенные основные модели.Таким образом, можно использовать крупнозернистые модели без потери общности, что значительно облегчает наше понимание метаболического поведения.
Используя формализм вектора стоимости, который мы использовали в доказательстве теоремы 1, мы можем изучать метаболизм в пространстве ограничений низкой размерности, а не в пространстве потоков высокой размерности (см. Рис. 3). В случае двух ограничений (также проиллюстрированных на рис. 1) принцип экстремума утверждает, что оба ограниченных пула ферментов всегда могут быть полностью использованы с двумя векторами затрат (EFM), не более.Однако EFM с диагональным вектором затрат может полностью использовать оба пула самостоятельно: следовательно, количество EFM, которые максимизируют поток, также может быть меньше количества активных ограничений. Другой случай, когда оптимальным является только один EFM, — это когда все векторы затрат лежат выше или ниже диагонали. В этом случае есть только одно активное ограничение; другой пул не ограничивает общий возможный поток системы в этих условиях. Мы вывели необходимые и достаточные условия, при которых оптимально использовать ЭФМ в смесях (S1 Приложение, раздел 5).Построение векторов затрат для различных концентраций внутренних метаболитов также показывает, что на длину и направление векторов затрат влияют концентрации метаболитов через кинетику ферментов (показано заштрихованными областями на рис. 1). Мы показываем в разделе 5 приложения S1, что эта зависимость от метаболитов делает гораздо более вероятным использование менее K EFM в системе с K ограничениями, потому что внутренние концентрации могут быть изменены, чтобы сделать векторы затрат диагональными.
Рис. 3. Иллюстрация принципа экстремума.
Принцип экстремума утверждает, что размерность пространства решений определяется количеством ограничений экспрессии ферментов, а не размерностью метаболической сети. Ограничения возникают из-за биофизических ограничений, например, ограниченных возможностей растворителя в клеточных компартментах. Наш формализм вектора затрат, поясненный на рис. 1, позволяет нам анализировать метаболизм в пространстве ограничений низкой размерности, а не в пространстве потоков высокой размерности, которое обычно используется.
https://doi.org/10.1371/journal.pcbi.1006858.g003
Количество ферментативных ограничений можно вывести из экспериментальных данных: Принцип экстремума применяется к избыточному метаболизму
Хорошо известное явление, наблюдаемое у микробов, — это избыточный метаболизм: очевидно расточительное выделение продуктов. Примерами являются аэробное производство этанола дрожжами (эффект Крэбтри), лактата раковыми клетками (Варбург) или ацетата Escherichia coli [4, 38, 39].Начало избыточного метаболизма обычно изучается как функция скорости роста (например, в хемостатах, где скорость роста определяется степенью разведения культуры). До некоторой критической скорости роста клетки полностью дышат, но когда скорость роста увеличивается выше некоторого критического значения, дыхательный поток уменьшается и возникает поток избыточного метаболизма.
Согласно нашей теории, дополнительное ферментативное ограничение должно было стать активным при критической скорости роста (см. Рис. 2).Ниже критической скорости роста респираторный поток пропорционален скорости роста, что является характеристикой однократного использования EFM (см. S1 Приложение, Раздел 8). Однако, превышая критическую скорость роста, уменьшающийся дыхательный поток и увеличивающийся поток перелива указывают на то, что должны быть активны по крайней мере два EFM и, следовательно, два ограничения. Действительно, все современные модели метаболизма переполнения используют такое дополнительное ограничение, но биофизическая природа первого ограничения (в основном ограничение поглощения) часто сохраняется неявной.Поэтому многие объяснения метаболизма переполнения, по-видимому, имеют только одно ограничение, например, связанное с общим белком [4] или мембранным белком [40], но в рамках нашей теории оптимальное распределение потока с двумя EFMs возможно только при наличии как минимум двух ограничений.
Мы можем получить больше информации о метаболизме переполнения, применив формализм вектора затрат к грубой модели (рис. 4 и приложение S2). Обратите внимание, однако, что эта модель имеет только иллюстративную цель, чтобы показать, что метаболизм переполнения может быть легко объяснен двумя ограничениями экспрессии ферментов.Мы не утверждаем, что наложенные ограничения являются реальными ограничениями; для этого необходимы эксперименты, как мы объясним позже. Модель включает дыхательный путь и ветвь перелива ацетата. Все этапы включают кинетику ферментов, и ограничения накладываются на два пула ферментов: общий цитозольный белок и общий мембранный белок. Мы моделируем метаболизм переполнения как функцию концентрации глюкозы, потому что даже несмотря на то, что экспериментально скорость роста устанавливается степенью разбавления ограниченного глюкозой хемостата, скорость роста всегда коррелирует с доступной концентрацией глюкозы.При низких внеклеточных концентрациях глюкозы все векторы затрат имеют высокую стоимость мембран и лежат выше или в лучшем случае по диагонали (поскольку мембранные ограничения находятся на оси Y): мембранный пул ограничивает поглощение субстрата и, следовательно, способствует эффективному использованию глюкозы посредством дыхания. . Наша базовая модель предсказывает, что по мере увеличения внеклеточной концентрации глюкозы увеличивается и уровень насыщения гликолитических ферментов, так что поток может увеличиваться без изменения уровня белка. Следовательно, в большом диапазоне внешних концентраций субстрата чистое дыхание приводит к максимальной скорости роста за счет полного использования двух доступных пулов ферментов.Однако мембранное ограничение в большей степени ограничивает рост, т.е. ослабление этого ограничения даст больший выигрыш в скорости роста. При высоких концентрациях глюкозы переносчики более насыщены (векторы затрат становятся короче в направлении мембраны), а вектор затрат дыхания становится ниже диагонального: при чистом дыхании пул мембранных белков будет использоваться недостаточно, в то время как цитозольный пул ограничивает дыхание. Лучшая стратегия — меньше дышать и сделать часть цитозольного пула доступной для другого EFM, который может использовать недостаточно используемый пул мембран.Чистый результат состоит в том, что смесь EFM достигает более высоких темпов роста, чем любой из двух EFM по отдельности.
Рис. 4. Формализм вектора затрат дает представление о том, как максимизация скорости роста приводит к избыточному метаболизму.
a ) Основная модель с двумя EFM, которые по отдельности приводят к синтезу клеток (оранжевый: дыхание и синий: переполнение ацетата). Все рассматриваемые реакции имеют ассоциированный фермент, активность которого зависит от кинетических параметров и концентраций метаболитов.Мы варьировали скорость роста, изменяя внешнюю концентрацию субстрата. Учитывая это внешнее условие, скорость роста была оптимизирована при двух ферментативных ограничениях (ограниченный цитозольный фермент Σ e i , цито ≤ 1 и ограниченная площадь мембраны e транспорт ≤ 0,3). b ) Прогнозируемые потоки поглощения субстрата, направленные на дыхание и переток, качественно согласуются с экспериментальными данными (показанными ранее на рис. 2) для нескольких микроорганизмов, масштабированных по скорости роста ( μ крит ) и скорости поглощения. ( q крит ) в начале переполнения [4, 38, 39]. c ) Векторы стоимости (сплошные стрелки) двух EFM до (слева) и после (справа) респироферментативного переключения. Координата x векторов затрат обозначает долю цитозольного объема, которая необходима для получения одной единицы потока от цели с соответствующей EFM. Координата y показывает необходимую долю доступной площади мембраны. Положение векторов затрат показано для оптимизированных концентраций метаболитов; заштрихованные области показывают альтернативные положения векторов затрат при различных концентрациях ферментов и метаболитов.Пунктирные векторы показывают использование EFM в оптимальном решении.
https://doi.org/10.1371/journal.pcbi.1006858.g004
Мы думаем, что многие опубликованные объяснения метаболизма переполнения объединены принципом экстремума. Добавленная стоимость не в том, что она дает еще одну модель, качественно отражающую метаболизм перелива, а в том, что она объясняет, почему опубликованные модели успешны, предлагая всеобъемлющую теорию. Действительно, в разделе 4 приложения S1 мы показываем, что объяснения метаболизма переполнения, предлагаемые другими методами моделирования, налагающими другие ограничения, такие как крупнозернистые модели целых клеток [4, 5] и основанные на ограничениях М-модели в масштабе генома [19, 41–43] математически являются примерами (или упрощениями) одной и той же задачи оптимизации с ограничениями, которую мы изучаем здесь.Таким образом, все их максимизаторы следуют принципу экстремума, а избыточный метаболизм должен быть результатом второго ограничения, которое становится активным. Так называемые ME-модели [36] относятся к немного иному классу математических задач, но начало метаболизма переполнения по-прежнему вызывается дополнительным активным ограничением. Однако, поскольку все вышеприведенные объяснения охватывают явление с разными ограничениями и решают одну и ту же математическую задачу, мы пока не можем сделать вывод о механистической природе ограничений.
Идентичность ферментативных ограничений может быть выявлена экспериментальными изменениями
Мы можем предсказать влияние экспериментальных возмущений на метаболизм с помощью формализма вектора стоимости. Примерами таких нарушений являются экспрессия нефункциональных белков или ингибирование ферментов, что, соответственно, можно интерпретировать как уменьшение ограниченного пула ферментов или увеличение стоимости векторов. Влияние таких возмущений на рост, когда выражены два EFM, было проанализировано в формализме вектора затрат (анализ см. В разделе 6 и 7 Приложения S1).На рис. 5 a –5 d мы прогнозируем (качественный) эффект уменьшения доступной площади в ограниченном пространстве для двух случаев (i) уменьшения обоих пулов ферментов на одинаковую величину; или (ii) сокращение только первого ограниченного пула. Впоследствии мы сравниваем эти предсказания с экспериментами по возмущениям, проведенными Basan et al. [4] (математический анализ см. В SI).
Рис. 5. Прогнозы и экспериментальные результаты изменения размера ограниченных пулов ферментов во время роста с использованием смеси EFM.
На графиках вектора стоимости, панели a ) и b ) красный вектор обозначает оптимальное решение в невозмущенном организме. При экспериментальном возмущении доступная область в пространстве ограничений может измениться, что обозначено заштрихованными серыми областями. Зеленые, синие и серые векторы показывают новые оптимальные решения при нарастающих возмущениях. Прогнозируемое влияние на поток через ацетатную ветвь показано на панелях c и d ). a, c ) Анализ возмущений, которые затягивают оба пула белков с одинаковым количеством, показывает, что поток и скорость роста будут пропорционально уменьшаться, как это наблюдалось экспериментально ( e )) для сверхэкспрессии LacZ на разных источниках углерода (данные Basan и другие.[4]). b, d ) Нарушения, которые сжимают пул ферментов, который в основном используется одним EFM (здесь обозначается CO 2 ), первоначально вызывают увеличение потока через другой EFM в смеси ( Ac ). В конце концов, при более сильных ограничениях этот поток также уменьшается. f ) Такое поведение наблюдается, в том числе, для экспериментов с ингибиторами трансляции с использованием хлорамфеникола (S1 Приложение, раздел 7).
https://doi.org/10.1371/journal.pcbi.1006858.g005
С помощью этого анализа мы предлагаем широко применимый экспериментальный подход для проверки вероятных ограничений, ограничивающих рост.Учитывая возможное ограничение, теория предполагает нарушение размера соответствующего ограниченного пула ферментов, например, путем экспрессии нефункционального белка в этом пуле. Затем влияние этого возмущения на поток через активные EFM можно сравнить с прогнозами, как на рис. 5. Теперь мы можем подтвердить или опровергнуть, действительно ли определенные ограниченные пулы ферментов ограничивают рост. В качестве альтернативы можно ингибировать определенный фермент; однако это создает риск подавления одних EFM в большей степени, чем других, в результате чего результаты потенциально невозможно интерпретировать.
Прогнозы возмущений также можно использовать для повторной интерпретации опубликованных экспериментов. Например, сверхэкспрессия неиспользованного белка LacZ совпадает с нашим предсказанным эффектом равного сокращения двух пулов ферментов (рис. 5, и ). Стоимость создания цитозольного белка LacZ, таким образом, составляет равную долю обоих ограничений. Мы думаем, что это можно объяснить тем, что LacZ можно считать средним белком с точки зрения потребности в ресурсах. Поскольку метаболизм уже настроен на оптимальное использование обоих ограниченных пулов ферментов, всем EFM теперь потребуется большее количество обоих ограниченных пулов ферментов для поддержания скорости роста (векторы затрат удлиняются).Следовательно, дополнительные затраты на синтез уменьшают оба ограниченных пула в одинаковой степени. Как следствие, этот анализ не может принять решение о биологической интерпретации ограничений.
Добавление хлорамфеникола является примером, когда наш анализ показывает, что один ферментативный пул затронут больше, чем другой (рис. 5 f )). Хлорамфеникол подавляет трансляцию, поэтому клетке требуется большее количество рибосом на единицу потока. Это снова увеличивает затраты на синтез белка, тем самым уменьшая оба пула.Однако набор данных показывает, что хлорамфеникол оказывает более доминирующее влияние на первый пул (ось x), чем на второй пул (ось y). Технически это происходит потому, что запрещение трансляции удлиняет векторы затрат всех EFM в направлении x в разной степени. Мы изучаем этот случай в разделе 7 приложения S1 и показываем, что эффекты эквивалентны эффектам изменения размера первого пула ферментов. Это означает, что увеличенное количество рибосом оказывает дополнительный эффект на первый пул, что вполне может быть связано с большим цитозольным объемом, который занимают рибосомы.Это предполагает, что одним из ограниченных пулов является сумма цитозольных белков.
Наш кинетический подход, основанный на ограничениях, обеспечивает новое биологическое понимание
Недостаточное использование ферментов, по-видимому, противоречит оптимальному распределению ресурсов. Например, Goel et al. [44] изучали переключение L. lactis со смешанно-кислотной ферментации на гомолактическую. Поскольку они обнаружили постоянную экспрессию белка в зависимости от скорости роста, они пришли к выводу, что этот метаболический переход не может быть объяснен соображениями стоимости белка.Однако на рис. 6 a ) мы показываем, что кинетическая модель, которая включает в себя различные силы ингибирования продуктом АТФ на пути ферментации, может привести к экспериментально наблюдаемому поведению, когда распределение белка оптимизировано. В нашей модели насыщение ферментов гомолактической ферментации быстро увеличивается с увеличением скорости роста, в то время как насыщение ферментов смешанной кислотной ферментации немного снижается из-за повышенного ингибирования продуктом АТФ. Таким образом, метаболический поток может быть перераспределен без изменения распределения белков (подробности приведены в приложении S3).Другой пример — экспрессия больших фракций недостаточно используемых белков E. coli при низких скоростях роста [45]. Это также согласуется с оптимальным распределением ресурсов, если рассматривать кинетику ферментов, так что их насыщение реагентами варьируется. В этих двух примерах недостаточное использование белков, таким образом, используется как показатель того, что микроорганизмы не оптимально распределяют свои ресурсы. Здесь мы показали, что эти предполагаемые контрпримеры могут фактически соответствовать оптимальному распределению ресурсов, если рассматривать кинетическую модель, включая, таким образом, различные концентрации метаболитов и насыщения ферментов.
Рис. 6. Недостаточное использование ферментов и совместное потребление можно понять с помощью нашего кинетического подхода, основанного на ограничениях.
a ) Показаны модели метаболического переключателя L. lactis (пунктирные линии) вместе с экспериментальными данными из [44]. Прогнозы потока для обоих путей выражаются как доля от общего потока через оба пути. Концентрации ферментов нормализуются до концентраций при скорости роста 0,15 и затем масштабируются по логарифмической шкале.Модель воспроизводит переключение от смешанно-кислотной к гомолактической ферментации при постоянных концентрациях фермента, так как она учитывает кинетику фермента. Подробности этой модели описаны в Приложении S3. Чтобы получить идеальное соответствие с данными, следует задействовать более крупную модель, но это выходит за рамки данной статьи. Мы подчеркиваем, что концентрации белка могут оставаться постоянными при изменении использования пути. b ) Показан пример метаболической сети с EFM, которые используют либо сукцинат, либо ксилозу (оранжевые и синие кружки соответственно), и EFM (зеленые кружки), в котором используются два источника углерода.Серые квадраты обозначают продукты, которые необходимы для роста клеток. EFM совместного потребления может синтезировать один клеточный компонент с сукцинатом, а другой — с ксилозой. Таким образом, реакция, соединяющая верхнюю и нижнюю части сети, становится несущественной. Это приводит к возможному снижению затрат на белок и, следовательно, к преимуществу в скорости роста. Мы действительно измерили увеличение скорости роста при совместном потреблении сукцината и ксилозы, как показано на вставке, где разные биологические реплики обозначены разными точками.Результаты других проверенных комбинаций можно найти в Приложении S4.
https://doi.org/10.1371/journal.pcbi.1006858.g006
При наличии нескольких источников углерода микроорганизмы могут потреблять их одновременно [46–48]. Мы экспериментально подтвердили, что E. coli потребляет вместе источники углерода только тогда, когда это увеличивает скорость его роста (Приложение S4). Однако пока неясно, почему совместное потребление может быть выгодным. Были созданы модели оптимизации, которые показывают одновременное поглощение субстрата [47, 48], но подход Hermsen et al.[47] является механистическим и не обеспечивает фундаментальной причины, а Beg et al. [48] утверждают, что «клетки, предпочтительно использующие более эффективный источник углерода, перерастут те, которые позволяют одновременно использовать другие источники углерода». Aidelberg et al. [46] заявляют, что подходы к оптимизации с одной целью не могут объяснить совместное потребление. Однако мы показываем, что существуют сопотребляющие EFM (приложение S4), которые снижают затраты на ресурсы на единицу скорости роста, что приводит к более высоким темпам роста. Эти новые EFM существуют, когда каждая подложка создает другой набор предшественников (см. Рис. 6 b ) для иллюстрации).Следовательно, совместное потребление может стать благоприятным, когда реакции, связывающие источник углерода с удаленным предшественником, больше не являются необходимыми. Следуя этим рассуждениям, можно было бы ожидать наибольшего улучшения роста, если бы субстраты попадали в метаболическую сеть далеко друг от друга. Действительно, мы, как и другие [47], наблюдали наибольшее улучшение роста, когда субстраты с низким содержанием гликолита сочетаются с субстратами с высоким содержанием гликолита.
Некоторые микробные стратегии, по-видимому, снижают скорость роста, такие как упреждающая экспрессия стрессовых белков [39] и альтернативных переносчиков питательных веществ [49], а также избыточная емкость рибосом [50].То, что эти стратегии все еще выбирались эволюцией, часто приписывают преимуществам приспособленности в динамических условиях. Однако в нашем подходе, основанном на ограничениях, эти типы поведения не обязательно должны приводить к снижению темпов роста. Некоторые из белковых пулов могут быть использованы не полностью, и экспрессия белков может тогда принести незначительные затраты или вообще не потребовать их. Например, наш анализ избыточного метаболизма показывает, что один из ограниченных пулов ферментов используется недостаточно при низких скоростях роста. Этот недостаточно используемый пул может вместить белки, которые могут оказаться благоприятными для будущих условий.Например, предположим, что микроорганизм сталкивается с цитозольным и мембранным ограничением, но предположим, что только мембранное ограничение активно при низких скоростях роста. Неиспользованная цитозольная емкость может быть использована для других целей. Единственная активность ограничения мембраны при низких скоростях роста действительно объясняет, почему O’Brien et al. наблюдали, что E. coli имеет «ограниченную питательными веществами» [36] область роста при медленном росте.
Обсуждение
Принцип экстремума, который мы вывели и проиллюстрировал в этой работе, предсказывает эволюционное направление в коротком временном масштабе, определяя оптимальные уровни экспрессии ферментов.В данный момент принцип экстремума предсказывает, что ресурсы перераспределяются между наиболее эффективными ферментами за счет других, менее активных на единицу фермента: эволюция уменьшает количество активных EFM. В более длительном масштабе времени кинетические параметры и стехиометрия сети могут эволюционировать, тем самым изменяя фенотипический потенциал: эволюция изменяет векторы затрат. В этой новой настройке принцип экстремума снова предсказывает минимальную сложность, хотя выбранные EFM и поток через эти EFM могут измениться.Наша теория предсказывает, что микроорганизм, выбранный для максимальной скорости роста, в статических условиях будет экспрессировать только небольшое количество EFM, и, следовательно, его метаболизм низкоразмерен. Это вполне могло быть объяснением простых линейных соотношений, которые показывают многие экспериментально измеренные зависимости [10–12]. Эта простота может также объяснить, как только небольшое количество метаболитов или белков («главных регуляторов», таких как CcpA или Crp), по-видимому, регулируют (центральный) метаболизм [51].
Понимание того, что размерность метаболизма ограничена числом активных ограничений, применимо к более ранним подходам к моделированию, в которых использовались принципы распределения ресурсов.Кроме того, мы показываем, что те же принципы справедливы и для нелинейных моделей, которые включают кинетику ферментов и, следовательно, зависимости от метаболитов. Модель кинетического саморепликатора, представленная Molenaar et al. [5], например, показывает не смешанные стратегии, а резкое переключение между дыханием и брожением, свидетельство единственного активного ограничения. В самом деле, хотя ограничение на мембранный белок было включено, размер клеток можно было свободно регулировать, чтобы облегчить это ограничение. Во многих исследованиях со стехиометрическими моделями в масштабе генома встречаются смешанные стратегии.Во всех этих исследованиях поток поглощения глюкозы был ограничен (первое ограничение) в сочетании с некоторой линейной комбинацией потоков, которая отражает (второе) ограничение, которое было в центре внимания исследования (емкость растворителя, осмотическое давление [19, 52], протеом пределы [4, 42], мембрана [20, 40]). Также в так называемых моделях ME (метаболизма и экспрессии) [36] и их вариантах [53] скорость роста фиксирована, а поглощение питательных веществ сведено к минимуму. Опять же, в этих моделях наблюдается переполнение, когда возникает дополнительное ограничение (общий протеом).
Даже несмотря на то, что максимизация скорости роста при постоянных условиях может иногда быть довольно грубым приближением давления отбора, мы ожидаем, что принцип экстремума предоставит «эволюционную стрелу времени». Когда условия часто меняются, в игру могут вступить другие аспекты, и приспособленность будет отражаться средней скоростью роста в среде, то есть геометрической скоростью роста [1]. Справедливы ли принципы экстремума для максимизации геометрической скорости роста — открытый вопрос для будущих теоретических работ.
Даже в статических условиях наша теория основана на предположении, что скорость метаболизма максимальна. В принципе, эта скорость не обязательно должна быть скоростью клеточного синтеза, но может быть другой метаболической реакцией. Это могло бы произойти, например, в случае специализации на многоклеточных организмах. Однако мы не знаем, достаточно ли в этих случаях селективное давление, чтобы максимизировать эту скорость. Более того, даже микроорганизмы не всегда оптимально настроены, поскольку было показано, что титрование ArcA может увеличить скорость роста E.coli на гликолитических субстратах [54]. Действительно, принцип экстремума не описывает метаболизм, если никакая скорость не максимизирована, и, таким образом, наша теория не описывает все неоптимальные точки в ландшафте фитнеса. Тем не менее, принцип, который характеризует пики и показывает направление роста в каждой точке ландшафта, по-прежнему может служить отличным ориентиром.
Успех методов моделирования на основе ограничений предполагает, что биофизические ограничения действительно формируют метаболизм микробов.Однако большинство ограничений, используемых в литературе, постулируются и остаются не подтвержденными. Кроме того, налагаемые ограничения часто не могут быть напрямую выведены из физиологии микроорганизмов. Наша теория предлагает механистический путь вперед для будущих методов моделирования на основе ограничений. Наша теория предполагает, что ограничение должно быть наложено для каждого клеточного компартмента с ограниченной способностью растворять белки. Поскольку количество компартментов у прокариот обычно меньше, чем у эукариот, из-за отсутствия органелл метаболическое поведение прокариот, как правило, проще.
Крупномасштабные кинетические модели еще не используются для изучения оптимального метаболизма. Максимизация скорости роста в таких моделях быстро становится вычислительно невыполнимой, потому что все концентрации метаболитов и ферментов должны быть настроены. Наши результаты могут служить руководством для решения этих больших нелинейных задач оптимизации. Скажем, в модели есть K ограничений, принцип экстремума гарантирует, что оптимум должен быть найден среди конических комбинаций K EFM. Этот факт уже использовался в случае одного ограничения в сети среднего размера [55]: EFM можно было оптимизировать отдельно (что является строго выпуклой задачей [56]), и была выбрана задача с наибольшей скоростью роста.Однако сомнительно, что эта вычислительная осуществимость может быть распространена на модели с большим количеством ограничений. С двумя ограничениями все пары EFM уже должны быть рассмотрены, и максимизация скорости в двух EFM при двух ограничениях больше не является выпуклой.
Принцип экстремума — это нулевая гипотеза о ходе определенного эволюционного процесса [57]. Это имеет прямое оперативное значение для стратегий эволюционной инженерии, когда может быть желательно увеличение или уменьшение сложности микробного метаболизма, например, в промышленной биотехнологии, когда преследуется совместное потребление различных сахаров из гидролизатов биомассы, или если предотвращается избыточный метаболизм во время гетерологичного метаболизма. производится попытка производства белка.Возможно, когда были определены ограничения, ограничивающие рост интересующего микроорганизма, их можно было бы нарушить, чтобы направить эволюцию в предпочтительном направлении.
Заключение
Наша теория предполагает, что метаболизм имеет лишь несколько рабочих степеней свободы. Смещая перспективу максимизации скорости от всей метаболической сети к ее представлению в формализме вектора затрат, мы уменьшили проблему до ее основных измерений, равных количеству ограничивающих рост биофизических ограничений.Вместе с принципом экстремума эта работа обеспечивает всеобъемлющую, молекулярную, основанную на ограничениях, перспективу микробного метаболизма.
карточек метаболизма | Quizlet
Клетки организма используют запас макроэлементов в крови, поддерживаемый адекватным потреблением с пищей, для клеточного метаболизма. Попадая в клетки организма, различные макроэлементы могут подвергаться различным анаболическим или катаболическим реакциям, в зависимости от потребностей и функций клетки организма и общих потребностей организма в любой момент времени.Различные метаболические процессы для каждого из макроэлементов:
Углеводы:
Гликолиз:
Катаболическая реакция, при которой из глюкозы образуется пировиноградная кислота. Это трехфазный процесс, в результате которого из одной молекулы глюкозы образуются две молекулы пировиноградной кислоты. Он требует двух молекул АТФ, но генерирует четыре — для чистого образования двух молекул АТФ, а также генерирует два кофермента НАДН.
Пировиноградная кислота может затем подвергнуться двум процессам:
1.Аэробный путь — в присутствии кислорода превращается в ацетил-КоА, чтобы войти в цикл лимонной кислоты
2. Анаэробный путь — в отсутствие кислорода превращается в молочную кислоту, которая может использоваться в качестве источника энергии в ограниченных количествах.
Гликогенез:
Избыточная глюкоза, которая не требуется для выработки АТФ, может превращаться в гликоген в печени и скелетных мышцах. Гликоген — это форма хранения глюкозы, состоящая из длинных цепочек молекул глюкозы.
Гликогенолиз:
Когда в крови недостаточно глюкозы для клеток организма, хранящаяся глюкоза (гликоген) может расщепляться с высвобождением глюкозы.Это происходит в печени, которая расщепляет гликоген на отдельные молекулы глюкозы, которые высвобождаются в кровь (гликоген в скелетных мышцах зарезервирован только для клеток скелетных мышц, а не возвращается в общую циркуляцию).
Липиды:
Триглицериды в крови расщепляются липопротеинлипазой на глицерин и жирные кислоты, которые легко проникают в клетки организма.
Глицерин может быть преобразован в молекулу пировиноградной кислоты, а жирные кислоты могут быть преобразованы непосредственно в ацетил-КоА.Таким образом, любой компонент триглицерида может войти в цикл лимонной кислоты для производства АТФ.
Липогенез: избыток глицерина и жирных кислот поглощается жировыми клетками и превращается в триглицериды для хранения в жировой ткани.
Липолиз: когда жирные кислоты и глицерин необходимы для выработки АТФ, триглицериды в жировой ткани расщепляются на эти компоненты, которые высвобождаются в кровь для обеспечения клеток этим пищевым топливом.
Белки:
Синтез белка:
Большинство аминокислот, потребляемых клетками организма, перестраиваются в белковые структуры, которые становятся частью клетки или транспортируются из клетки с функциональной способностью (например,грамм. ферменты).
Трансаминирование и дезаминирование:
Когда аминокислоты должны использоваться для выработки АТФ, они должны быть преобразованы в модифицированную кетокислоту, которая затем может войти в цикл лимонной кислоты. Это происходит посредством трансаминирования с последующим дезаминированием:
= Трансаминирование: аминогруппа аминокислоты переносится на кетокислоту из цикла лимонной кислоты с образованием глутаминовой кислоты. Оставшаяся аминокислота теперь представляет собой другую кетокислоту, которая может быть модифицирована с образованием пировиноградной кислоты, ацетил-КоА или других кетокислот цикла лимонной кислоты (например,грамм. щавелевоуксусная кислота).
= Дезаминирование: аминогруппа глутаминовой кислоты удаляется в виде аммиака (Nh4) и объединяется с диоксидом углерода с образованием мочевины и воды. Впоследствии мочевина выводится из организма с мочой.
каждого макроэлемента, способного превращаться либо в пировиноградную кислоту, либо в ацетил-КоА, некоторые из макроэлементов могут в конечном итоге превращаться друг в друга!
Глюкоза, однажды расщепленная на пировиноградную кислоту или ацетил-КоА, может превращаться в глицерин и жирные кислоты с образованием триглицеридов.Это обычный процесс, когда в крови присутствует избыток глюкозы, особенно потому, что организм может хранить больше жира (триглицеридов) в жировой ткани, чем гликогена.
Аналогичным образом, жиры и белки могут при необходимости превращаться в глюкозу в процессе, известном как глюконеогенез. Когда для клеток организма недостаточно глюкозы, триглицериды и аминокислоты могут использоваться для выработки АТФ, но также могут быть преобразованы в глюкозу (путем объединения двух пировиноградных кислот для образования молекулы глюкозы).Глюконеогенез происходит в печени и является важным процессом для поддержания адекватного уровня глюкозы в крови.
Меняется ли метаболизм у двух людей?
Скорость обмена веществ в покое
Одно исследование [1] отметило, что одно стандартное отклонение дисперсии для скорости метаболизма в покое (сколько калорий сжигается при жизни) составляло 5-8%; это означает, что 1 стандартное отклонение населения (68%) было в пределах 6-8% от средней скорости метаболизма. В дополнение к этому, 2 стандартных отклонения населения (96%) находились в пределах 10–16% от среднего значения для населения. [1]
Если рассматривать это на практике и предполагать, что средний расход составляет 2000 ккал в день, 68% населения попадает в диапазон 1840–2160 ккал в день, в то время как 96% населения находится в диапазоне 1680–2320 ккал в день. Сравнение кого-либо с процентилем 5 -го или ниже с кем-то с процентилем 95 -го или выше дало бы разницу в 600 ккал в день, и вероятность этого (сравнение себя с другом) равна 0.50%, если предположить, что два совершенно случайных человека.
Чтобы дать представление о калориях, 200 ккал (разница в скорости обмена веществ примерно у половины населения) примерно эквивалентны 2 столовым ложкам арахисового масла, одному поптарт (упаковка из двух по 400 ккал) или половине большого ломтика. пиццы. Орео составляет около 70 ккал, а плитка шоколада — от 150 до 270 ккал в зависимости от марки.
Скорость метаболизма действительно различается, и технически может иметь большое отклонение.Однако со статистической точки зрения маловероятно, что это отклонение применимо к вам. Большинство людей живут в диапазоне 200-300 ккал друг от друга и не обладают сильно различающейся скоростью метаболизма.
Скорость обмена веществ и упражнения
Упражнения и физическая активность достоверно увеличивают скорость метаболизма, и в вышеупомянутом исследовании отмечается, что 1 стандартное отклонение в отношении этого составляет 1-2%. [1] Если предположить, что кто-то тренируется 300 ккал (в среднем), истинное значение естественных затрат калорий может варьироваться от 294–306 ккал (68% населения) или 288–312 ккал (96%). [1] Хотя общий расход энергии, по-видимому, является самым сильным коррелятом, предсказывающим потерю жира в результате упражнений, [2] следует отметить, что изменения в безжировой массе (мышцах) могут в дальнейшем влиять на скорость метаболизма. Скорость метаболизма тесно связана с мышечной массой (одна увеличивается вместе с другой) [3] , и процесс гипертрофии скелетных мышц (рост мышц, которые вы можете сокращать) требует высоких калорий. [4]
Эта «низкая» корреляция, по-видимому, применима к аэробным упражнениям (кардиоупражнениям, которые можно поддерживать с постоянной скоростью в течение длительного периода времени). [1] Тренировки с отягощениями (поднятие тяжестей) или высокоинтенсивные интервальные кардиоупражнения (спринт или любое другое упражнение, которое нельзя поддерживать в течение длительного времени и, следовательно, должны выполняться с перерывами) имеют так называемое EPOC, потребление кислорода после тренировки, которое является повышение скорости обмена веществ после тренировки.
Хотя стандартная тренировка с отягощениями (в которой участники не были принуждены к истощению) имеет относительно низкое значение EPOC, например 19 дополнительных калорий через час после тренировки, [5] упражнения более высокой интенсивности, такие как протокол «Табата» ( 170% VO2 max в течение 20 секунд с 10-секундным перерывом; повторение 8 раз, всего 4 минуты) связаны с большим потреблением EPOC и кислорода. [6] [7] В этих анаэробных упражнениях отмеченная выше корреляция имеет тенденцию к нарушению, и отклонение 1-2%, вероятно, не применяется.
Физические упражнения вызывают повышение скорости метаболизма у всех людей в силу естественного характера физического движения. Менее вариативны базовые аэробные движения, такие как ходьба или бег трусцой. Низкая дисперсия может не соблюдаться для упражнений высокой интенсивности
Активная жизнь
Новое исследование, в котором изучались 11 японских мужчин в условиях жизни, но в метаболической камере для измерения скорости метаболизма с помощью тепла, было обнаружено, что после измерения всех участников в дни «низкой», «умеренной» и «высокой» интенсивности, это одно и то же. люди, у которых средний уровень метаболизма 2228 +/- 143 ккал во время низкой активности увеличили дневные расходы до 2816 +/- 197 ккал (среднее увеличение на 588 ккал) во время умеренной физической активности и до 2813 +/- 163 ккал во время высокой активности. [8] Увеличение скорости метаболизма за счет умеренно энергичного активного образа жизни (в основном, «упорная» работа, но с трудом потеет и не теряет дыхание) может свести на нет разницу в скорости метаболизма, обнаруживаемую у большинства (68% +) население (с использованием чисел из первого процитированного исследования). [1]
Умеренно активный образ жизни может быть достаточным, чтобы свести на нет присущие различия в метаболизме для многих людей, не находящихся на крайних точках метаболической колоколообразной кривой (то есть тех, у кого вряд ли будет диагностирован гипотиреоз; низкий уровень метаболизма)
Как Влияет ли метаболизм на ваши шансы на потерю веса?
Здоровый образ жизни может улучшить ваш метаболизм, и наоборот
Основные моменты
- Сжигание большего количества калорий, чем вы потребляете, может помочь вам похудеть
- Здоровый обмен веществ может ускорить процесс похудения
- Метаболизм во многом зависит от вашего образа жизни
Процесс, с помощью которого ваше тело превращает пищу в энергию, известен как метаболизм.Здоровый обмен веществ — необходимое условие для похудания. Если у вас слабый метаболизм, ваша диета и режим упражнений будут давать результаты медленнее. Метаболизм — это сложный биохимический процесс, во время которого калории, содержащиеся в пище и напитках, соединяются с кислородом и высвобождают энергию, которая важна для правильного функционирования организма. Даже когда вы отдыхаете, вашему телу нужна энергия для выполнения таких действий, как кровообращение, дыхание, баланс гормонов, а также рост и восстановление клеток.
Метаболизм и потеря веса
Это обычная тенденция обвинять метаболизм в увеличении веса. Однако только в случае медицинских проблем, таких как недостаточная активность щитовидной железы, вы набираете вес из-за слабого обмена веществ.
Увеличение веса является результатом вашей диеты, генов, образа жизни, сна, стресса и физической активности. Вы набираете вес, когда сжигаете меньше калорий, чем потребляете. Для похудения важно, чтобы вы либо ели меньше калорий, а сжигали больше за счет физической активности, либо делали и то, и другое.Хотя может быть правдой, что некоторые люди могут похудеть быстрее и легче, чем другие, люди обычно худеют, когда сжигают больше калорий, чем едят.
Также прочтите: 8 лучших трав, которые сжигают жир и ускоряют метаболизм
Мы спрашиваем тренера по стилю жизни Люка Коутиньо о связи между метаболизмом и потерей веса. Он согласен с тем, что метаболизм действительно стал «модным словом» в области похудания. «В то время как некоторые люди переходят от одной колонны к другой в поисках способов повысить метаболизм, некоторые благодарят свои гены», — говорит он.
Далее он добавляет, что метаболизм — это не что иное, как скорость, с которой ваше тело сжигает калории. «Наши митохондрии — крошечные клеточные двигатели, которые создают энергию для тела — являются центральной частью нашего метаболизма и пищеварения. То, насколько быстро и эффективно митохондрии могут превращать питательные вещества, такие как глюкоза, в энергию, отражает метаболическое здоровье. Если ваши митохондрии неэффективны, вы — хранят глюкозу в виде жира. Это то, что делает вялый метаболизм », — объясняет Люк.
Ваш режим питания и уровень активности могут определять ваш метаболизм.
Фото предоставлено: iStock
Также прочтите: 7 напитков, повышающих метаболизм, чтобы тонизировать ваше тело
Образ жизни играет огромную роль в определении вашего метаболизма.Правильные изменения в образе жизни могут улучшить ваш метаболизм, а неправильные — парализовать его.
«От вашего режима питания и привычек до уровня активности и того, как вы тренируетесь (упражнения), потребление воды, привычки сна и даже качество мыслей, которые приходят вам в голову, оказывают огромное влияние на ваш метаболизм», — говорит Люк DoctorNDTV.
Время от времени человек может придерживаться здоровой диеты, регулярно заниматься спортом и быть в курсе того, чтобы делать правильные упражнения каждый день, но при этом не терять вес.Стресс может быть главной причиной этого. Стресс может вызвать у вас усталость и неспособность выполнять некоторые изнурительные упражнения. Ваше тело может не реагировать на вашу диету и режим упражнений должным образом, когда вы находитесь в состоянии стресса.
Люк далее добавляет, что прием пищи каждые 2-3 часа, отказ от завтрака или тяжелых тренировок не обязательно улучшат ваш метаболизм. «Они скорее вредны, чем полезны, когда дело касается обмена веществ и похудания. Нам нужна смена парадигмы, чтобы похудеть правильно, и это ОБРАЗ ЖИЗНИ», — подчеркивает он.
Также прочтите: 5 напитков, которые ускоряют ваш метаболизм и помогают мгновенно сжечь жир на животе
(Люк Коутиньо, тренер по холистическому образу жизни — интегративная медицина)
Отказ от ответственности: этот контент, включая советы, предоставляет только общую информацию. Это никоим образом не заменяет квалифицированное медицинское заключение. Для получения дополнительной информации всегда консультируйтесь со специалистом или вашим лечащим врачом. NDTV не несет ответственности за эту информацию.
Ожидание ответа для загрузки.