Что такое метаболизм, за что он отвечает
О метаболизме обычные люди и эксперты говорят довольно много. Однако далеко не все понимают, в чём именно заключается его основное предназначение. Многие задумываются над вопросом: Как ускорить метаболизм? Другие же даже понятия не имеют, для чего это делать и как это обстоятельство отразиться на качестве жизни. Давайте разбираться в деталях.
Как метаболизм может влиять на жизнь человека?
Считается, что люди с отличным метаболизмом способны съесть много пищи, быстро её переработать и при этом оставаться стройными и полными сил. Заложники медленного метаболизма должны быть максимально избирательными в части рациона. Им нужно выбирать те продукты, которые не способствуют накоплению жировых отложений, при этом употребляя их небольшими дозами. Именно поэтому каждый заинтересован в улучшении показателей метаболизма.
Что же такое метаболизм?
Метаболизм – это обмен веществ, или все процессы, протекающие в организме, в результате которых еда и напитки, попадающие внутрь, превращаются в живительную энергию, так необходимую всем органам и системам человека.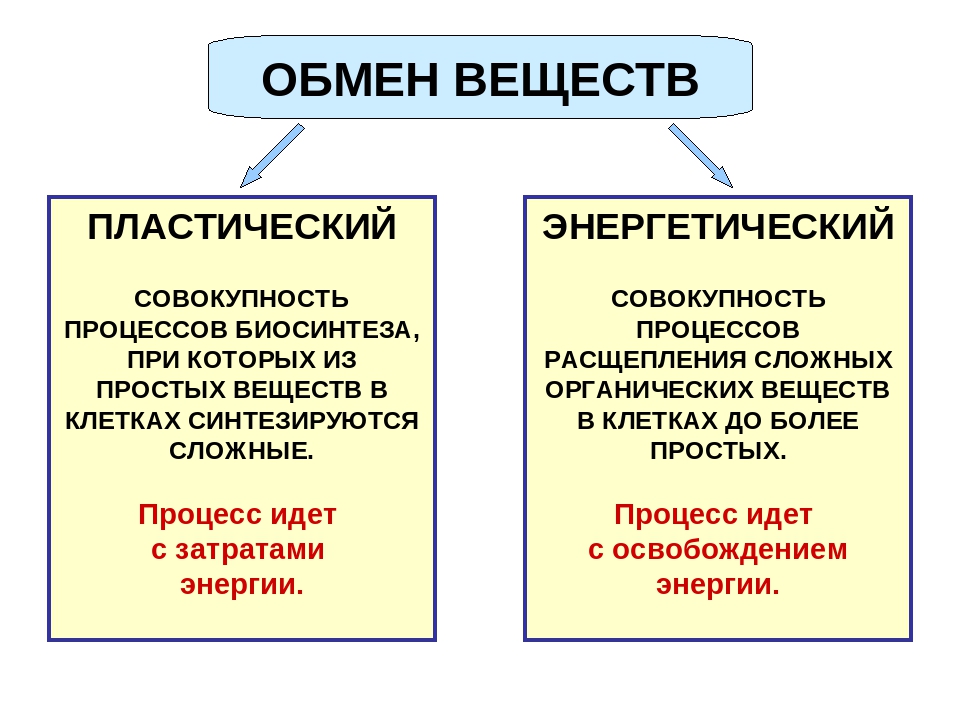
Метаболизм состоит из трёх подгрупп:
- Метаболизм в состоянии полного покоя;
- Метаболизм во время активных движений;
- Термический эффект от употребляемых в пищу продуктов.
Метаболизм в состоянии покоя
Когда человек не двигается, кажется, что все процессы остановились. Однако это не так. Их в организме происходит великое множество. Мозг обрабатывает информацию, сердце качает кровь, лёгкие обогащают клетки кислородом, желудок переваривает пищу, слуховые рецепторы находятся в поиске информации и так далее. Показателем метаболизма являются калории, которые организм затрачивает на банальное поддержание жизнедеятельности.
Потребности среднестатистического человека обычно ограничиваются показателем в 60-70% затрат. На качество метаболизма в состоянии полного покоя могут воздействовать самые различные факторы, к примеру, вес человека и его рост, пол, масса тела без жира. Установлено, что метаболизм выше у тех, у кого больше сухой массы.
Термический эффект от употребляемых в пищу продуктов
Здесь подразумевается количество калорий или энергии, которое расходуется организмом на сам процесс переваривания пищи, а также поэтапное усвоение полезных элементов, поступающих вместе с продуктами. Углеводы, белки и жиры выделяются различным термическим эффектом. Именно поэтому рацион должен быть сбалансированным и вымеренным, исходя из потребностей отдельного индивидуума.
Метаболизм во время активных движений
Здесь говорится о количестве энергии, которую человек дополнительно расходует во время активной ходьбы, бега, физической работы, упражнений и прочей деятельности. Активность бывает тренировочной или нетренировочной. В первом случае затраты энергии в полной мере зависят от характера и стиля упражнений. На нетренировочную активность приходится немало затрат энергии от общего количества. Сюда можно отнести, к примеру, ходьбу и даже банальное поддержание осанки, во время сидения на стуле.
Метаболизм может быть плохим или хорошим. При этом важно понимать, что существует достаточно инструментов, методик и способов изменить ситуацию в лучшую сторону, или наладить работу организма. Однако сделать это будет не так просто. Для нормализации алгоритмов, понадобится время. Консультация с профильным врачом носит обязательный характер.
18+
На правах рекламыЦитохром P450 2C19. Генотипирование по маркеру CYP2C19 G681A
Маркер связан с особенностями метаболизма лекарств в печени. Исследуется для выявления физиологической эффективности применения препаратов: антиагреганты (клопидогрел и его аналоги), ингибиторы протонного насоса (омепразол, лансопразол), антидепрессанты (амитриптилин), противогрибковые (вориконазол), психотропные (диазепам), противоопухолевые (тамоксифен) и др.
Альтернативное обозначение генетического маркера CYP2C19*2.
Метод исследования
Полиморфизм длин рестрикционных фрагментов.
Какой биоматериал можно использовать для исследования?
Венозную кровь, буккальный (щечный) эпителий.
Как правильно подготовиться к исследованию?
Специальной подготовки не требуется.
Название гена
CYP2C19.
OMIM
*124020.
Локализация гена на хромосоме
10q23.33.
Функция гена
Ген CYP2C19 кодирует белок CYP2C19 (S-мефенитоингидроксилаза) – изофермент семейства цитохрома Р450, участвующий в метаболизме ряда лекарственных препаратов в печени.
Генетический маркер CYP2C19 (G681A)
Замена нуклеотида гуанин (G) в пятом экзоне в позиции 681 на аденин (A) последовательности ДНК гена CYP2C19 обозначается как генетический маркер CYP2C19 G681A.
Основной аллель (681G) — CYP2C19*1.
Минорный аллель (681A) — CYP2C19*2.
Возможные генотипы
Встречаемость в популяции
Аллель CYP2C19* 2 встречается у европейцев с частотой 15 %.
Ассоциация маркера с метаболизмом лекарственных препаратов
Исследуется для выявления физиологической эффективности применения следующих препаратов: антиагреганты (клопидогрел и его аналоги), ингибиторы протонного насоса (омепразол, лансопразол), антидепрессанты, противогрибковые (вориконазол) и др.
Общая информация об исследовании
Для достижения эффекта лекарственных препаратов в организме необходима их биоактивация (трансформация в активную форму) в клетках печени (гепатоцитах) системой ферментов цитохрома Р450 (CYP). Гены, кодирующие эти ферменты, полиморфны, причем часто встречаются аллели, кодирующие образование ферментов со сниженной или отсутствующей функцией.
На активность цитохромов, помимо особенностей строения кодирующих их генов, оказывают влияние такие факторы, как возраст, масса тела, образ жизни, вредные привычки, особенности диеты, сопутствующие заболевания, прием лекарственных препаратов. Эти факторы отвечают за формирование индивидуальных особенностей работы ферментов Р450 и определяют характер метаболизма большинства лекарств. Среди цитохромов Р450 чаще всего с пониженной чувствительностью к ряду препаратов связывают полиморфизм гена
Фармакогенетическое исследование рекомендуется перед началом приема ряда препаратов. Например, клопидогрел (торговое название «Плавикс», аналоги — «Зилт» и «Эгитромб») — это антитромбоцитарный препарат, который широко используется для лечения и профилактики рецидива инфаркта миокарда, инсульта и других проявлений ишемической болезни сердца, также назначается пациентам, перенесшим вмешательства на коронарных артериях (коронарное стентирование).
Выраженность антитромбоцитарного эффекта препарата клопидогрел у людей существенно различается.
Существуют аллели CYP2C19, кодирующие образование фермента со сниженной или отсутствующей функцией. Вариант гена, несущий точечную замену G681A, ведет к утрате функции фермента и обозначается как CYP2C19*2. Замена одного нуклеотида влечет за собой образование преждевременного стоп-кодона. Таким образом, получается неактивный (укороченный) фермент CYP2C19. [PubMed: 8195181]. Основной (неизмененный) вариант гена обозначается как CYP2C19*1.
Активный метаболит клопидогрела в норме связывается с тромбоцитами, ингибируя их АДФ-индуцируемую агрегационную активность, блокируя рецептор к АДФ. У носителей аллеля CYP2C19*2 снижен уровень активного метаболита клопидогрела, наблюдается уменьшенное ингибирование агрегации тромбоцитов и, как следствие, повышенный риск нежелательных реакций со стороны сердечно-сосудистой системы и повышение риска тромбоза стента при лечении клопидогрелом, по сравнению с пациентами с нормальной функцией фермента.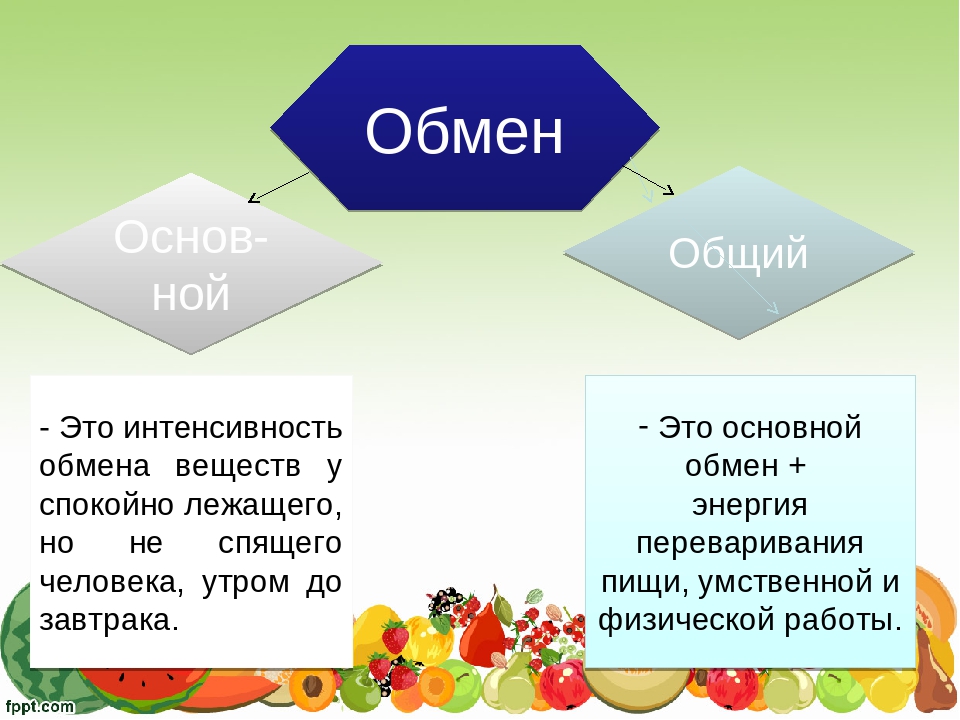
Использование клопидогрела в комбинации с рядом других препаратов, которые подавляют активность фермента (некоторые антидепрессанты, противогрибковые препараты, ингибиторы протонной помпы, применяемые для лечения гастритов и язвы желудка), у носителей аллеля CYP2C19*2, приводит к практически полному отсутствию терапевтического эффекта.
Генетическое исследование поможет определиться с дозировкой препарата клопидогрел, тактикой лечения и не допустить рецидива осложнений сердечно-сосудистых заболеваний.
Интерпретация результатов
На основании генотипа CYP2C19 (G681A) можно выделить три основных фенотипа метаболизатора CYP2C19: «медленный» (отсутствие функции фермента или значительно сниженная функция), «замедленный (промежуточный)» (сниженная функция фермента) и «быстрый» (нормальная функция фермента, терапевтический эффект будет достаточным).
Варианты генотипов:
*1/*1 – нормальная функция фермента;
*1/*2 – сниженная функция фермента;
*2/*2 – значительно сниженная функция фермента или её отсутствие.
На основании выявленного генотипа в комплексе с другими генетическими, анамнестическими, клиническими и лабораторными данными врач может выбрать соответствующую стратегию лечения.
что это? У кого бывает ускоренный метаболизм? Как ускорить метаболизм?
Метаболизм – это все химические процессы, протекающие в человеческом организме. В результате этих процессов белки, жиры, углеводы превращаются в другие вещества и выделяется энергия, которую организм использует для своих нужд.
Например, наша кожа, волосы, ногти регулярно обновляются: для их построения нужны «кирпичики» — белки и жиры, а также «рабочая сила» — энергия. Это и есть метаболизм.
Метаболизм разделяют на два вида:
основной, который происходит постоянно, когда человек находится в состоянии покоя;
дополнительный, связанный с активностью организма, например, физической.

Затраты энергии при основном обмене веществ — это калории, которые уходят на поддержание температуры тела, работы сердца, легких, мозга, органов пищеварения, а также различных реакций в организме. Эти потребности восполняются с помощью питания.
У каждого человека имеется своя норма калорий в день. Она зависит от телосложения, пола, веса, возраста, уровня ежедневной физической активности и состояния щитовидной железы.
Энергетические потребности организма после 21 года начинают снижаться, в среднем на 2%-3% за десятилетие. А потребности детей, наоборот, в два раза выше, чем у взрослых.
Сегодня в интернете существуют калькуляторы расчета метаболизма, которые помогают рассчитать индивидуальное суточное количество калорий, необходимое для поддержания жизнедеятельности в состоянии покоя.
Как был открыт метаболизм
Понятие обмена веществ впервые встречается в работах врача Ибн аль-Нафиса (1213—1288), который писал, что «тело и его части находятся в постоянном состоянии распада и питания, так что оно неизбежно претерпевает постоянные изменения».
Первые контролируемые эксперименты по метаболизму у человека опубликовал другой врач — Санториов 1614 году. Он рассказал, как сам взвесил себя до и после приема пищи, сна, работы, натощак, после питья и выделения мочи, и обнаружил, что большая часть пищи, которую он принял, была утрачена в результате процесса, названного им «незаметным испарением».
В начале ХХ века Эдуард Бухнер открыл ферменты, что позволило еще точнее изучать метаболические реакции. Ферменты – это вещества, которые ускоряют химические реакции в организме человека.
Как происходит обмен веществ
Процесс метаболизма делят на три стадии:
Белки, жиры, углеводы поступают вместе с пищей в организм человека, где они взаимодействуют с веществами, ускоряющими химические реакции – ферментами и распадаются на более простые вещества. Белки распадаются на аминокислоты, углеводы на глюкозу, жиры на глицерин и жирные кислоты.
Все полученные питательные элементы всасываются в кровь и вместе с ней передвигаются по всему организму к тканям, клеткам.
Далее эти элементы еще больше распадаются до конечных продуктов и при этом выделяется энергия, которая нужна для обеспечения слаженной работы всего организма.Побочные продукты, которые организму не нужны, выводятся через пот, кал, мочу, легкие.
 Далее эти элементы еще больше распадаются до конечных продуктов и при этом выделяется энергия, которая нужна для обеспечения слаженной работы всего организма.
Далее эти элементы еще больше распадаются до конечных продуктов и при этом выделяется энергия, которая нужна для обеспечения слаженной работы всего организма.Что влияет на скорость обмена веществ
Обычно, услышав фразы «ускоренный метаболизм» или «медленный метаболизм», люди представляют возможность сохранять стройность без физических нагрузок и ограничений в еде, либо же наоборот легко набирать вес. Однако это не совсем так.
У людей с быстрым метаболизмом на жизненно важные функции, например, работу сердца и лёгких, за одно и то же время тратится больше энергии, чем у обладателей медленного обмена веществ.
Поэтому при одинаковых нагрузках один человек питается булочками, мгновенно сжигая все полученные калории, а другой будет стремительно набирать вес – это значит, что у них разная скорость основного обмена.
Есть два типа факторов, влияющих на метаболизм: статические и динамические.
Статические факторы — это все, на что невозможно повлиять: наследственность, пол, возраст, тип телосложение.
Динамические факторы позволяют немного ускорить или замедлить обмен веществ: масса тела, физические нагрузки, организация рациона питания, психоэмоциональное состояние.
Однако нельзя начать изнурять себя постоянными тренировками и жесткой диетой. Так, при снижении калорийности питания, организм сокращает потребности в энергии, когда он в ней нуждается. Человеку это только вредит — он становится сонным, заторможенным. Эти процессы ускоряются, если при соблюдении диеты выполняются активные тренировки на жиросжигание.
Как ускорить метаболизм?
Для ускорения метаболизма придерживайтесь следующих рекомендаций:
Не истощать себя жесткой диетой. Питаться следует 5-6 раз в день, небольшими порциями.
Не пренебрегать жирами. Часто худеющие отказываются от продуктов, содержащих жир.
Необходимо употреблять в пищу растительные масла, рыбу, орехи.
Важно высыпаться, так как недостаток сна тормозит обмен веществ. Кроме этого вырабатывается кортизол – «гормон стресса», который негативно влияет на организм.
Пить воду. Вода помогает улучшить метаболические процессы, притупляет чувство голода. При недостатке воды у организма появляется новая задача – восполнить водный баланс, а не сжечь жиры.
Употреблять продукты, содержащие йод. За обмен веществ в организме отвечает щитовидная железа. Йод активизирует ее работу. Он содержится в морской капусте, креветках, кальмарах, яйцах, кукурузе, черносливе, тунце.

Неинфекционные заболевания
Общая информация
Неинфекционные заболевания (НИЗ), также известные как хронические заболевания, как правило, имеют продолжительное течение и являются результатом воздействия комбинации генетических, физиологических, экологических и поведенческих факторов.
К основным типам НИЗ относятся сердечно-сосудистые заболевания (такие, как инфаркт и инсульт), раковые заболевания, хронические респираторные заболевания (такие, как хроническая обструктивная болезнь легких и астма) и диабет.
Бремя НИЗ диспропорционально велико в странах с низким и средним уровнем доходов, на которые приходится более трех четвертей случаев смерти от НИЗ во всем мире (32 миллиона).
Кто подвергается риску таких заболеваний?
НИЗ распространены во всех возрастных группах, всех регионах и всех странах. Эти заболевания часто связывают с пожилыми возрастными группами, но фактические данные свидетельствуют о том, что среди всех случаев смерти, связываемых с НИЗ, 17 миллионов регистрируется в возрастной группе от 30 до 69 лет. При этом более 85% этих случаев «преждевременной» смерти имеет место в странах с низким и средним уровнем доходов. Дети, взрослые и пожилые люди — все уязвимы перед факторами риска, способствующими развитию НИЗ, такими как нездоровое питание, недостаточная физическая активность, воздействие табачного дыма или злоупотребление алкоголем.
Развитию этих заболеваний способствуют такие факторы, как быстрая и неорганизованная урбанизация, глобализация нездорового образа жизни и старение населения. Последствия нездорового питания и недостаточной физической активности могут проявляться у отдельных людей в виде повышенного кровяного давления, повышенного содержания глюкозы в крови, повышенного уровня липидов в крови и ожирения.
Последствия нездорового питания и недостаточной физической активности могут проявляться у отдельных людей в виде повышенного кровяного давления, повышенного содержания глюкозы в крови, повышенного уровня липидов в крови и ожирения.
Факторы риска
Поддающиеся изменению поведенческие факторы риска
Поддающиеся изменению формы поведения, такие как употребление табака, отсутствие физической активности, нездоровое питание и вредное употребление алкоголя, — повышают риск развития НИЗ.
- От последствий употребления табака каждый год умирает более 7,2 млн. человек (в том числе от последствий воздействия вторичного табачного дыма), и по прогнозам в предстоящие годы эта цифра заметно вырастет . (1)
- Каждый год от последствий чрезмерного потребления соли/натрия умирает 4,1 млн. человек. (1)
- Из 3,3 млн. случаев смерти в год в результате употребления алкоголя более половины приходятся на долю НИЗ, в том числе раковых заболеваний . (2)
- 1,6 млн. случаев смерти в год можно признать связанными с недостаточным уровнем физической активности . (1)
 случаев смерти в год можно признать связанными с недостаточным уровнем физической активности . (1)
случаев смерти в год можно признать связанными с недостаточным уровнем физической активности . (1)Метаболические факторы риска
Метаболические факторы риска способствуют развитию четырех основных изменений метаболизма, приводящих к повышению риска НИЗ:
- повышенное кровяное давление
- излишний вес/ожирение
- гипергликемия (высокое содержание глюкозы в крови)
- гиперлипидемия (высокое содержание липидов в крови)
Первым метаболическим фактором риска смерти от НИЗ во всем мире является повышенное кровяное давление (с которым связано 19% всех случаев смерти в мире) (1), за которым следуют избыточный вес, ожирение и повышенное содержание глюкозы в крови.
Каковы социально-экономические последствия НИЗ?
НИЗ ставят под угрозу прогресс на пути осуществления Повестки дня в области развития на период до 2030 г., где предусмотрена цель по снижению числа случаев преждевременной смерти от НИЗ на одну треть к 2030 году.
Существует тесная связь между бедностью и НИЗ. По прогнозам, стремительный рост заболеваемости НИЗ будет препятствовать реализации инициатив в области сокращения бедности в странах с низким уровнем доходов, в частности, приводя к росту медицинских расходов домохозяйств. Представители уязвимых и незащищенных категорий населения болеют чаще и умирают в более молодом возрасте по сравнению с представителями более благополучных социальных групп, главным образом, в силу своей подверженности более высокому риску воздействия со стороны вредной для здоровья продукции, такой как табак, или склонности к нездоровому питанию, а также ввиду ограниченного доступа к услугам здравоохранения.
По прогнозам, стремительный рост заболеваемости НИЗ будет препятствовать реализации инициатив в области сокращения бедности в странах с низким уровнем доходов, в частности, приводя к росту медицинских расходов домохозяйств. Представители уязвимых и незащищенных категорий населения болеют чаще и умирают в более молодом возрасте по сравнению с представителями более благополучных социальных групп, главным образом, в силу своей подверженности более высокому риску воздействия со стороны вредной для здоровья продукции, такой как табак, или склонности к нездоровому питанию, а также ввиду ограниченного доступа к услугам здравоохранения.
В условиях низкой обеспеченности ресурсами медицинские расходы, связанные с НИЗ, приводят к быстрому истощению ресурсов домохозяйств. Каждый год непомерные издержки, связанные с НИЗ, включая расходы, связанные с нередко продолжительным и дорогостоящим лечением, и последствия потери кормильца, ввергают людей в нищету и мешают развитию.
Профилактика НИЗ и борьба с ними
Важным способом борьбы с НИЗ является целенаправленная деятельность по сокращению воздействия факторов риска развития этих болезней. Есть недорогостоящие решения, которыми правительства и другие заинтересованные стороны могли бы воспользоваться для сокращения воздействия поддающихся изменению факторов риска. При выработке мер политики и расстановке приоритетов важное значение имеет мониторинг прогресса и тенденций в области НИЗ.
Есть недорогостоящие решения, которыми правительства и другие заинтересованные стороны могли бы воспользоваться для сокращения воздействия поддающихся изменению факторов риска. При выработке мер политики и расстановке приоритетов важное значение имеет мониторинг прогресса и тенденций в области НИЗ.
Для снижения негативного воздействия НИЗ на отдельных людей и общество в целом требуется комплексный подход, в реализации которого должны участвовать все сектора, включая здравоохранение, финансы, транспорт, образование, сельское хозяйство, планирование и другие, работая в сотрудничестве друг с другом над сокращением рисков, связанных с НИЗ, и содействуя осуществлению мероприятий по профилактике этих болезней и борьбе с ними.
Инвестиции в более эффективное ведение НИЗ имеют важнейшее значение. Ведение НИЗ включает выявление, скрининг и лечение этих болезней и предоставление доступа к паллиативной помощи для всех нуждающихся. Высокоэффективные основные мероприятия в отношении НИЗ могут осуществляться на базе первичной медико-санитарной помощи, что позволит укрепить раннее выявление и своевременное лечение. Фактические данные показывают, что при условии их своевременного проведения такие мероприятия с экономической точки зрения являются прекрасной инвестицией, поскольку они могут снизить потребность в более дорогостоящем лечении.
Фактические данные показывают, что при условии их своевременного проведения такие мероприятия с экономической точки зрения являются прекрасной инвестицией, поскольку они могут снизить потребность в более дорогостоящем лечении.
Страны с недостаточным охватом медицинским обслуживанием едва ли будут способны обеспечить всеобщий охват основными мероприятиями по лечению и профилактике НИЗ. Такие мероприятия имеют важнейшее значение для достижения поставленной на глобальном уровне цели относительного сокращения риска преждевременной смертности от НИЗ на 25% к 2025 г. и задачи ЦУР по снижению числа случаев преждевременной смерти от НИЗ на одну треть к 2030 году.
Ответные действия ВОЗ
Роль ВОЗ в области общего руководства и координации
В повестке дня в области устойчивого развития на период до 2030 г. НИЗ признаны одним из главных препятствий на пути к устойчивому развитию. В рамках Повестки дня главы государств и правительств взяли на себя обязательство принять на национальном уровне решительные меры с тем, чтобы к 2030 г. сократить на одну треть преждевременную смертность от НИЗ посредством лечебно-профилактической работы (задача 3.4 ЦУР). Эта задача была поставлена по итогам проведения Совещаний высокого уровня Генеральной Ассамблеи ООН по НИЗ в 2011 и 2014 гг., на которых вновь подтверждалась роль ВОЗ в области общего руководства и координации в сфере мониторинга и содействия глобальной работе по борьбе с НИЗ. Генеральная Ассамблея ООН проведет третье Совещание высокого уровня в 2018 г. для анализа хода работы и выработки консенсуса относительно дальнейших действий на период 2018-2030 гг.
сократить на одну треть преждевременную смертность от НИЗ посредством лечебно-профилактической работы (задача 3.4 ЦУР). Эта задача была поставлена по итогам проведения Совещаний высокого уровня Генеральной Ассамблеи ООН по НИЗ в 2011 и 2014 гг., на которых вновь подтверждалась роль ВОЗ в области общего руководства и координации в сфере мониторинга и содействия глобальной работе по борьбе с НИЗ. Генеральная Ассамблея ООН проведет третье Совещание высокого уровня в 2018 г. для анализа хода работы и выработки консенсуса относительно дальнейших действий на период 2018-2030 гг.
Для оказания странам поддержки в их работе на национальном уровне ВОЗ подготовила Глобальный план действий в области профилактики неинфекционных заболеваний и борьбы с ними на 2013-2020 гг., включающий девять глобальных целей, достижение которых окажет наибольшее влияние на глобальные показатели смертности от НИЗ. Эти цели касаются вопросов профилактики и ведения НИЗ.
Справочная литература
(1) GBD 2015 Risk Factors Collaborators. Global, regional, and national comparative risk assessment of 79 behavioural, environmental and occupational, and metabolic risks or clusters of risks, 1990–2015: a systematic analysis for the Global Burden of Disease Study 2015. Lancet, 2016; 388(10053):1659-1724
Global, regional, and national comparative risk assessment of 79 behavioural, environmental and occupational, and metabolic risks or clusters of risks, 1990–2015: a systematic analysis for the Global Burden of Disease Study 2015. Lancet, 2016; 388(10053):1659-1724
Страница не найдена |
Страница не найдена |404. Страница не найдена
Архив за месяц
ПнВтСрЧтПтСбВс
13141516171819
20212223242526
2728293031
12
12
1
3031
12
15161718192021
25262728293031
123
45678910
12
17181920212223
31
2728293031
1
1234
567891011
12
891011121314
11121314151617
28293031
1234
12
12345
6789101112
567891011
12131415161718
19202122232425
3456789
17181920212223
24252627282930
12345
13141516171819
20212223242526
2728293031
15161718192021
22232425262728
2930
Архивы
Метки
Настройки
для слабовидящих
понятий метаболизма | SpringerLink
ChapterПервый онлайн:
Abstract
Хотя группа виталистов первоначально использовала термин «органический» для соединений, производимых только организмами, позже он был использован для соединений углерода.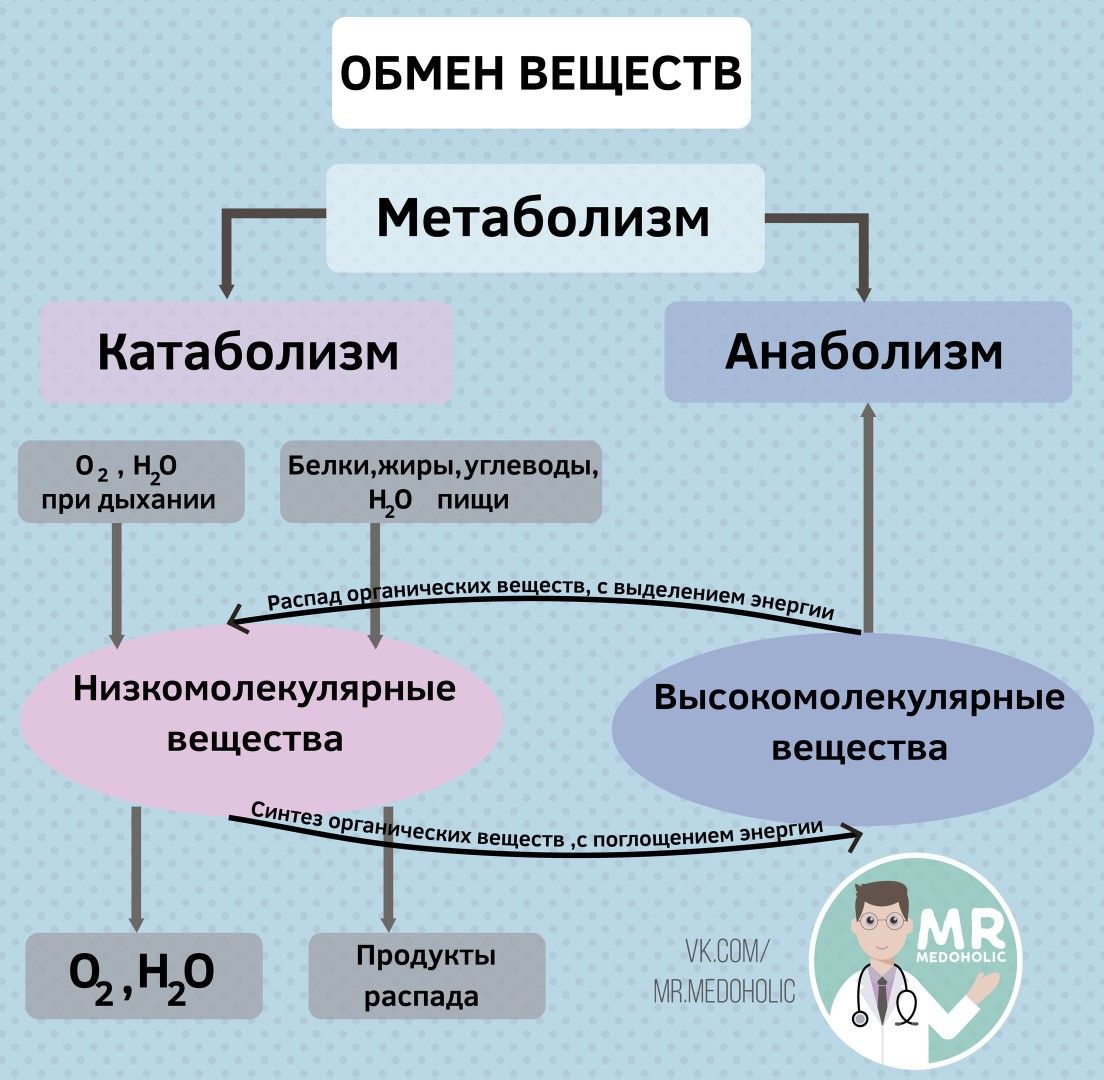 Wohler (1928) обнаружил, что мочевина, которая раньше считалась производимой только в живых существах, также может быть произведена в лаборатории из аммиака и бикарбоната. В 1897 году немецкие химики Эдуард Бюхнер и Ганс Бюхнер продемонстрировали, что ферментация может осуществляться с помощью бесклеточного экстракта дрожжей .Эти наблюдения привели к развитию науки биохимии. В начале двадцатого века, благодаря открытию различных метаболических путей, в биохимии преобладала органическая химия, за которой следовали энзимология и биоэнергетика . Некоторые из аналитических методов, которые сделали возможным изучение биохимии, включали выделение органелл, высокоэффективную жидкостную хроматографию, электрофорез, использование радиоактивных индикаторов, методы трансформации растений с использованием Agrobacterium tumefaciens, подавление генов , прямую генетику, обратную генетику, масс-спектрометрию, и микроматрица ДНК, среди прочего.С помощью вычислительных технологий теперь можно полностью понять взаимосвязь метаболических путей.
Wohler (1928) обнаружил, что мочевина, которая раньше считалась производимой только в живых существах, также может быть произведена в лаборатории из аммиака и бикарбоната. В 1897 году немецкие химики Эдуард Бюхнер и Ганс Бюхнер продемонстрировали, что ферментация может осуществляться с помощью бесклеточного экстракта дрожжей .Эти наблюдения привели к развитию науки биохимии. В начале двадцатого века, благодаря открытию различных метаболических путей, в биохимии преобладала органическая химия, за которой следовали энзимология и биоэнергетика . Некоторые из аналитических методов, которые сделали возможным изучение биохимии, включали выделение органелл, высокоэффективную жидкостную хроматографию, электрофорез, использование радиоактивных индикаторов, методы трансформации растений с использованием Agrobacterium tumefaciens, подавление генов , прямую генетику, обратную генетику, масс-спектрометрию, и микроматрица ДНК, среди прочего.С помощью вычислительных технологий теперь можно полностью понять взаимосвязь метаболических путей.
Ключевые слова
Аллостерическая регуляция Биоэнергетика Кофактор Сопряженные реакции Свободная энергия Гиббса Холофермент Метаболическое каналирование Метаболическая избыточность Константа Михаэлиса-Ментен Редокс-потенциалЭто предварительный просмотр содержимого подписки,
войдите в систему, чтобы проверить доступ.
вопросов с множественным выбором
- 1.
Биоэнергетика относится к:
- (a)
Обмен энергией между клеткой и окружающей средой
- (b)
Наука, которая занимается преобразованием энергии внутри клетки
- (в)
Выделение энергии при химической реакции
- (г)
- (a)
- 2.
Живая клетка является открытой системой, потому что:
- (a)
Она не обменивается энергией или веществом с окружающей средой.
- (b)
Он может обмениваться энергией и веществом с окружающей средой.
- (c)
Он может обмениваться энергией с окружающей средой, но не с веществом.
- (d)
Он может обмениваться веществом, но не энергией, с окружающей средой.
- (a)
- 3.
Согласно второму закону термодинамики, произойдет спонтанная реакция:
- (a)
Когда менее сложные молекулы будут преобразованы в более сложные.
- (б)
Когда происходит поглощение энергии из окружающей среды.
- (c)
Молекулы с более высокой энтропией преобразуются в молекулы с меньшей энтропией.
- (d)
Молекулы с меньшей энтропией превращаются в молекулы с высокой энтропией.
- (a)
- 4.
ΔG клеточной реакции будет отрицательным, если:
- (a)
Продукты реакции имеют меньшую энтропию, чем реагенты.
- (б)
Продукты реакции обладают большей энтропией, чем реагенты.
- (c)
Реакция не является спонтанной.
- (d)
Для протекания реакции требуется подвод энергии.
- (a)
- 5.
Какое из следующих утверждений верно?
- (a)
В живой клетке константа равновесия поддерживается равной 0.
- (b)
Свободная энергия — это полная энергия, присутствующая в молекуле.
- (c)
Свободная энергия — это энергия, изотермически доступная для выполнения работы.
- (d)
ΔG ° ‘ определяется как изменение свободной энергии во время реакции, которая не находится в равновесии.
- (a)
- 6.
Высокоэнергетическая связь (~) АТФ означает:
- (a)
Для образования этой связи требуется энергия.
- (b)
Гидролиз этой связи высвобождает энергию.
- (c)
Продукты гидролиза обладают меньшей энергией, чем сама молекула.
- (г)
Продукты гидролиза обладают большей энергией, чем сама молекула.
- (a)
- 7.
АТФ — это молекула с высокой энергией, потому что:
- (a)
Это нуклеозидтрифосфат.
- (б)
АТФ более резонансно стабилизирован, чем продукты его гидролиза.
- (c)
АТФ присутствует в клетке в виде комплекса Mg-ATP.
- (г)
Продукты его гидролиза стабилизируются за счет резонанса.
- (a)
- 8.
В окислительно-восстановительной реакции движутся электроны:
- (a)
От соединений с более положительным окислительно-восстановительным потенциалом к соединениям с меньшим положительным окислительно-восстановительным потенциалом
- (b)
От соединений с меньшим положительным окислительно-восстановительным потенциалом до более положительного окислительно-восстановительного потенциала
- (c)
От соединений с меньшим отрицательным окислительно-восстановительным потенциалом до соединений с более отрицательным окислительно-восстановительным потенциалом
- (d)
- (a)
- 9.
В молекуле фермента, состоящей из белковой и небелковой структур, белковая часть известна как:
- 10.
Небелковая структура, ковалентно присоединенная к белковой части молекулы фермента, называется:
- 11.
Константа Михаэлиса (K
m) фермента:
- (a)
Концентрация субстрата, при которой фермент полностью насыщен
- (b)
Концентрация субстрата при где V max составляет половину максимального значения
- (c)
Концентрация фермента при V max
- (d)
Концентрация фермента, при которой V max составляет половину максимальной
- (a)



Рекомендуемая дополнительная литература
Jones RL, Ougham H, Thomas H, Waaland S (2013) Молекулярная жизнь растений.Wiley-Blackwell, Chichester, pp 42–70
Google ScholarNelson DL, Cox MM (2017) Принципы биохимии Ленингера, 7-е изд.
Google Scholar W.H. Freeman, New York, pp 495–525Voet DJ, Voet JG, Charlotte WP (2008) Принципы биохимии, 3-е изд. Wiley, Hoboken, pp. 448–484
Google Scholar
 W.H. Freeman, New York, pp 495–525
W.H. Freeman, New York, pp 495–525Информация об авторских правах
© Springer Nature Singapore Pte Ltd. 2018
Авторы и аффилированные лица
- 1.Кафедра ботаникиКирори Мал Колледж, Университет Дели Нью-ДелиИндия
Границы | Проблемы городского метаболизма: устойчивость и благополучие в городах
Концепция городского метаболизма восходит к девятнадцатому веку (Wolman, 1965; Odum, 1996), поскольку города стали двигателем экономического развития и благосостояния людей. Термин городской метаболизм, как подчеркивают Сеспедес Рестрепо и Моралес-Пинсон (2018), «представляет собой концепцию, в которой город использует биологическое понятие, относящееся к внутренним процессам, посредством которых живые организмы поддерживают непрерывный обмен вещества и энергии с окружающей средой. для обеспечения работы, роста и воспроизводства.«Города — это сложные экосистемы, полностью естественные, даже если мы иногда думаем, что созданные человеком структуры не являются природой. Понять города намного легче, если мы осознаем, что они представляют собой естественные экосистемы, подчиняющиеся тем же законам (принцип максимальной мощности Лотки, эволюция Дарвина, естественный отбор), что и все другие части и виды. Хотя города покрывают <2% поверхности Земли, они потребляют около 78% энергии, доступной на планете, к которой необходимо добавить количество материалов и продуктов (продукты питания, строительные минералы, металлы и т. Д.)), которые косвенно также требуют потребления энергии. Эта цифра может быть объяснена с экономической и социальной точек зрения, поскольку города предлагают жителям новые возможности для бизнеса, образования, здравоохранения, безопасности, социальной жизни. Поддержка этой деятельности (и связанных с ней выгод) требует огромных потоков ресурсов, что чаще всего приводит к экологическому стрессу как на местном, так и на глобальном уровне.
для обеспечения работы, роста и воспроизводства.«Города — это сложные экосистемы, полностью естественные, даже если мы иногда думаем, что созданные человеком структуры не являются природой. Понять города намного легче, если мы осознаем, что они представляют собой естественные экосистемы, подчиняющиеся тем же законам (принцип максимальной мощности Лотки, эволюция Дарвина, естественный отбор), что и все другие части и виды. Хотя города покрывают <2% поверхности Земли, они потребляют около 78% энергии, доступной на планете, к которой необходимо добавить количество материалов и продуктов (продукты питания, строительные минералы, металлы и т. Д.)), которые косвенно также требуют потребления энергии. Эта цифра может быть объяснена с экономической и социальной точек зрения, поскольку города предлагают жителям новые возможности для бизнеса, образования, здравоохранения, безопасности, социальной жизни. Поддержка этой деятельности (и связанных с ней выгод) требует огромных потоков ресурсов, что чаще всего приводит к экологическому стрессу как на местном, так и на глобальном уровне. Следовательно, оценка ресурсного метаболизма городской системы, хотя и очень трудная из-за ее сложности, становится все более важной, не только с точки зрения взаимосвязи с окружающей средой как источника или поглотителя, но и с точки зрения внутренней динамики обмена ресурсами между компонентами города. секторы, а также конкуренция между городами за ограниченные ресурсы.За последние несколько лет наблюдается рост интереса к городам и их использованию энергии и материалов — отчасти из-за проблем изменения климата, а отчасти из-за социальных проблем, обусловленных социальными и организационными проблемами, порожденными огромными миграционными потоками из сельской местности в другие. городские районы. Города играют центральную роль в сокращении выбросов CO 2 и борьбе с изменением климата, одним из наиболее серьезных вызовов, с которыми в настоящее время сталкивается наше общество (Zhang et al., 2018).Изменение климата — не единственная проблема, определяемая городами и влияющая на них: заброшенность сельских районов, потребность в воде, отводимой от естественных путей, концентрация городских отходов и потребность в соответствующих способах их удаления, потребность в транспорте.
Следовательно, оценка ресурсного метаболизма городской системы, хотя и очень трудная из-за ее сложности, становится все более важной, не только с точки зрения взаимосвязи с окружающей средой как источника или поглотителя, но и с точки зрения внутренней динамики обмена ресурсами между компонентами города. секторы, а также конкуренция между городами за ограниченные ресурсы.За последние несколько лет наблюдается рост интереса к городам и их использованию энергии и материалов — отчасти из-за проблем изменения климата, а отчасти из-за социальных проблем, обусловленных социальными и организационными проблемами, порожденными огромными миграционными потоками из сельской местности в другие. городские районы. Города играют центральную роль в сокращении выбросов CO 2 и борьбе с изменением климата, одним из наиболее серьезных вызовов, с которыми в настоящее время сталкивается наше общество (Zhang et al., 2018).Изменение климата — не единственная проблема, определяемая городами и влияющая на них: заброшенность сельских районов, потребность в воде, отводимой от естественных путей, концентрация городских отходов и потребность в соответствующих способах их удаления, потребность в транспорте. и связанные с этим ограничения по пространству, энергии и загрязнению воздуха, огромное количество ресурсов для поддержки городского образа жизни (образование, медицинские услуги, цепочка поставок продуктов питания и промышленных товаров и т. д.) не только требуют энергии и вызывают загрязнение, но и являются источником загрязнения. потенциальный источник новой динамики рынка, реорганизации рабочей силы, конфликтов, новых инструментов управления и дорожных карт, спроса на общественные услуги и, наконец, упадка прежних культурных и социальных моделей и их замены новыми (или ничем, что еще хуже).Мониторинг притока и оттока ресурсов и понимание того, как они соотносятся с населением, доступностью ресурсов и пропускной способностью окружающей среды, имеют решающее значение для осознанной и заинтересованной политики устойчивого развития городов и управления. Реорганизация городских структур, связывающая биофизические аспекты с социальными аспектами благополучия сообществ, является приоритетной задачей при разработке политики во всем мире.
и связанные с этим ограничения по пространству, энергии и загрязнению воздуха, огромное количество ресурсов для поддержки городского образа жизни (образование, медицинские услуги, цепочка поставок продуктов питания и промышленных товаров и т. д.) не только требуют энергии и вызывают загрязнение, но и являются источником загрязнения. потенциальный источник новой динамики рынка, реорганизации рабочей силы, конфликтов, новых инструментов управления и дорожных карт, спроса на общественные услуги и, наконец, упадка прежних культурных и социальных моделей и их замены новыми (или ничем, что еще хуже).Мониторинг притока и оттока ресурсов и понимание того, как они соотносятся с населением, доступностью ресурсов и пропускной способностью окружающей среды, имеют решающее значение для осознанной и заинтересованной политики устойчивого развития городов и управления. Реорганизация городских структур, связывающая биофизические аспекты с социальными аспектами благополучия сообществ, является приоритетной задачей при разработке политики во всем мире. Чтобы влиять на метаболизм в городах, нам необходимо лучше понимать отношения между обществами, окружающей средой, массой и потоками энергии (производство и потребление), не недооценивая фактор роста населения.Благополучие, а не богатство или изобилие, будет ключевым словом для будущего городов, а именно осознанных и сбалансированных отношений городского населения с окружающей средой, ресурсами, другими видами, чтобы способствовать справедливому и равноправному доступу к социальным услугам. , улучшение культуры, экономики и здоровья не обязательно связано с увеличением потребления ресурсов на душу населения.
Чтобы влиять на метаболизм в городах, нам необходимо лучше понимать отношения между обществами, окружающей средой, массой и потоками энергии (производство и потребление), не недооценивая фактор роста населения.Благополучие, а не богатство или изобилие, будет ключевым словом для будущего городов, а именно осознанных и сбалансированных отношений городского населения с окружающей средой, ресурсами, другими видами, чтобы способствовать справедливому и равноправному доступу к социальным услугам. , улучшение культуры, экономики и здоровья не обязательно связано с увеличением потребления ресурсов на душу населения.
Распределение человеческого населения по всему миру ясно показывает, что все больше и больше людей переезжают из сельской местности в города, что приводит к росту проблем (и осведомленности о) доступности и использования энергии и ресурсов, городскому развитию и целостности окружающей среды (Cui, 2018) .Согласно отчетам Организации Объединенных Наций, к 2030 году будут существовать 43 мегаполиса (т. е. города с населением более 10 миллионов человек), большинство из которых в развивающихся странах (United Nations, 2018b). Ожидается, что наибольшая часть мирового населения будет проживать в городах или мегаполисах, и ожидаемое значительное изменение стиля жизни, цепочек поставок, моделей управления, общественной организации, производства отходов, образовательных инструментов и структур и многого другого в общей экологической, экономической и социальной динамике, трансформируются в острую необходимость во всесторонних исследованиях устойчивости и благополучия городов.
е. города с населением более 10 миллионов человек), большинство из которых в развивающихся странах (United Nations, 2018b). Ожидается, что наибольшая часть мирового населения будет проживать в городах или мегаполисах, и ожидаемое значительное изменение стиля жизни, цепочек поставок, моделей управления, общественной организации, производства отходов, образовательных инструментов и структур и многого другого в общей экологической, экономической и социальной динамике, трансформируются в острую необходимость во всесторонних исследованиях устойчивости и благополучия городов.
Городские системы и города представляют собой сложные образования со своим собственным метаболизмом, чрезвычайно зависимыми от различных материальных и энергетических ресурсов извне (Currie et al., 2017). Связь воды и энергии в городской системе с учетом моделей потребления воды и энергии в зданиях, жилых секторах и производственных секторах может дать важную информацию для устойчивого городского планирования (Fan et al. , 2019). Метаболизм ресурсов влияет и зависит от того, как городские сообщества самоорганизуются и принимают решения, потребляют и сохраняют, деградируют и перерабатывают ресурсы, совместно используют общественные услуги, проектируют и используют инфраструктуры.Тем не менее, общественный аспект и управление метаболизмом ресурсов все еще плохо изучены. Вторая важная тема — это то, как городские системы соотносятся с окружающими сельскими районами и дикой природой, уважают их, помогают им развиваться и сохранять свои модели целостности (Wei et al., 2015). Наконец, много было написано о замедлении роста активов, населения и потребления, но это очень сильно ограничивалось количественным замедлением, без изучения все еще возможных моделей качественного роста с точки зрения качества работы, безопасности, взаимодействия с сообществом, совместного использования. , счастье, справедливый доступ к ресурсам на всех уровнях и совместные процессы.Это, вероятно, самый сложный, но также и самый важный аспект устойчивости и стабильности городов.
, 2019). Метаболизм ресурсов влияет и зависит от того, как городские сообщества самоорганизуются и принимают решения, потребляют и сохраняют, деградируют и перерабатывают ресурсы, совместно используют общественные услуги, проектируют и используют инфраструктуры.Тем не менее, общественный аспект и управление метаболизмом ресурсов все еще плохо изучены. Вторая важная тема — это то, как городские системы соотносятся с окружающими сельскими районами и дикой природой, уважают их, помогают им развиваться и сохранять свои модели целостности (Wei et al., 2015). Наконец, много было написано о замедлении роста активов, населения и потребления, но это очень сильно ограничивалось количественным замедлением, без изучения все еще возможных моделей качественного роста с точки зрения качества работы, безопасности, взаимодействия с сообществом, совместного использования. , счастье, справедливый доступ к ресурсам на всех уровнях и совместные процессы.Это, вероятно, самый сложный, но также и самый важный аспект устойчивости и стабильности городов. Интегрированное городское управление посредством надлежащего управления и экологической оценки преимуществ и компромиссов в отношении планирования развития, пространственного планирования инфраструктуры, а также соответствующего вспомогательного вмешательства становится фундаментальным (Fan et al., 2019). Вместо того, чтобы просто заниматься городским развитием и сохранением экологии, градостроители всегда должны помнить об аспектах устойчивости, связанных с качеством и типологией метаболизма (Cui, 2018).Проблема не в том, чтобы максимизировать или обеспечить огромное энергоснабжение или максимально возможное количество и разнообразие продуктов питания или самый быстрый транспорт, а в том, чтобы оптимизировать на всех уровнях доступ всего городского населения к услугам, товарам, образованию, отдыху, здоровью. услуг, качества окружающей среды в достаточной степени для обеспечения качества жизни, стабильности, справедливости и уважения.
Интегрированное городское управление посредством надлежащего управления и экологической оценки преимуществ и компромиссов в отношении планирования развития, пространственного планирования инфраструктуры, а также соответствующего вспомогательного вмешательства становится фундаментальным (Fan et al., 2019). Вместо того, чтобы просто заниматься городским развитием и сохранением экологии, градостроители всегда должны помнить об аспектах устойчивости, связанных с качеством и типологией метаболизма (Cui, 2018).Проблема не в том, чтобы максимизировать или обеспечить огромное энергоснабжение или максимально возможное количество и разнообразие продуктов питания или самый быстрый транспорт, а в том, чтобы оптимизировать на всех уровнях доступ всего городского населения к услугам, товарам, образованию, отдыху, здоровью. услуг, качества окружающей среды в достаточной степени для обеспечения качества жизни, стабильности, справедливости и уважения.
Благополучие в городах, также называемое пригодностью для жизни в городе, представляет способность города обеспечивать адекватные условия для процветания и хорошего качества жизни граждан (del Mar Martínez-Bravo et al. , 2019). Доступность ресурсов и их справедливое распределение / распределение, индивидуальное восприятие, различные организации и условия сообщества (например, уровень образования, общественные услуги, наличие свободного времени, готовность зеленых насаждений, среди прочего), экологическая осведомленность, социальное и экономическое развитие это взаимосвязанные вопросы, которые сильно влияют на метаболизм города. Устойчивость и благополучие городских территорий были темой обсуждения как в научном мире (Wei et al., 2015; del Mar Martínez-Bravo et al., 2019), а также политические дебаты (United Nations, 2018a) из-за решающей связи между ростом, эксплуатацией природных ресурсов (использование невозобновляемых источников энергии и материалов) и последствиями для состояния среда.
, 2019). Доступность ресурсов и их справедливое распределение / распределение, индивидуальное восприятие, различные организации и условия сообщества (например, уровень образования, общественные услуги, наличие свободного времени, готовность зеленых насаждений, среди прочего), экологическая осведомленность, социальное и экономическое развитие это взаимосвязанные вопросы, которые сильно влияют на метаболизм города. Устойчивость и благополучие городских территорий были темой обсуждения как в научном мире (Wei et al., 2015; del Mar Martínez-Bravo et al., 2019), а также политические дебаты (United Nations, 2018a) из-за решающей связи между ростом, эксплуатацией природных ресурсов (использование невозобновляемых источников энергии и материалов) и последствиями для состояния среда.
Городские системы широко исследовались с помощью нескольких различных экономических и биофизических подходов. Показатели эффективности для городов, регионов и стран были разработаны на основе хорошо известных методов оценки, иногда интегрированных в специальный инструментарий: воплощенная энергия, анализ материальных потоков (MFA), анализ жизненного цикла (LCA), выбросы CO 2 , затраты / выгода и экономическая отдача (Yong et al. , 2012). Эти индикаторы по отдельности или в комбинации не обязательно обеспечивают полностью адекватную характеристику экологической целостности городской системы и использования ресурсов, поскольку они не были разработаны для оценки целостных систем, замкнутых циклов и характеристик обратной связи, которые являются ключевыми характеристиками устойчивого городского развития. экономика (Yong et al., 2013). Чаще всего в этих подходах не учитываются качество и характеристики потока, а также сложность взаимодействия между природной средой и социально-экономическими системами (Huang et al., 2006). Поэтому очень важно обсудить и глубже понять сложность будущего развития и управления городами, выходя далеко за рамки линейного и одномерного подхода простого измерения городского населения, потребления энергии или ВВП. Связь городского метаболизма со стратегиями политики необходима для измерения и изменения показателей устойчивости городов, развития междисциплинарной практики городского метаболизма (Dijst et al.
, 2012). Эти индикаторы по отдельности или в комбинации не обязательно обеспечивают полностью адекватную характеристику экологической целостности городской системы и использования ресурсов, поскольку они не были разработаны для оценки целостных систем, замкнутых циклов и характеристик обратной связи, которые являются ключевыми характеристиками устойчивого городского развития. экономика (Yong et al., 2013). Чаще всего в этих подходах не учитываются качество и характеристики потока, а также сложность взаимодействия между природной средой и социально-экономическими системами (Huang et al., 2006). Поэтому очень важно обсудить и глубже понять сложность будущего развития и управления городами, выходя далеко за рамки линейного и одномерного подхода простого измерения городского населения, потребления энергии или ВВП. Связь городского метаболизма со стратегиями политики необходима для измерения и изменения показателей устойчивости городов, развития междисциплинарной практики городского метаболизма (Dijst et al. , 2018). На социальное поведение и индивидуальные предпочтения сильно влияет конкуренция за доступные ресурсы, пространство, возможности.Невозможно иметь образованное следующее поколение без вложений в образование; невозможно наслаждаться здоровой пищей, не инвестируя ресурсы в более чистое производство; Невозможно иметь стабильное и счастливое сообщество без обеспечения равного доступа к основным ресурсам. Как следствие, надлежащее и справедливое управление ресурсами, вероятно, повлияет (и будет затронуто) образованием, демократией, стабильностью, чувством принадлежности, сбалансированным взаимодействием с окружающими территориями и населением.
, 2018). На социальное поведение и индивидуальные предпочтения сильно влияет конкуренция за доступные ресурсы, пространство, возможности.Невозможно иметь образованное следующее поколение без вложений в образование; невозможно наслаждаться здоровой пищей, не инвестируя ресурсы в более чистое производство; Невозможно иметь стабильное и счастливое сообщество без обеспечения равного доступа к основным ресурсам. Как следствие, надлежащее и справедливое управление ресурсами, вероятно, повлияет (и будет затронуто) образованием, демократией, стабильностью, чувством принадлежности, сбалансированным взаимодействием с окружающими территориями и населением.
Одной из самых сложных задач ближайшего будущего является разработка новых экологических стратегий и альтернатив ископаемым источникам энергии с целью удовлетворения потребностей и ожиданий нынешнего и будущих поколений на устойчивой основе.Во времена сокращения ресурсной базы более круговые и гибкие модели предоставления ресурсов в сочетании с изменениями образа жизни могут стать основой менее ресурсоемкой городской жизни (Zucaro et al. , 2014). Так как городская территория представляет собой сложную систему, связанную с различными производственно-потребительскими цепочками поставок, исследование экологических, экономических и социальных аспектов с использованием общесистемной основы анализа для обеспечения устойчивости является ключевым вопросом для защиты благосостояния и разработки адекватных политических решений. .Даже если энергоэффективность и внедрение возобновляемых источников энергии, а также модели рециркуляции материалов (круговая экономика), кажется, развиваются быстро и эффективно, многое еще предстоит сделать. Общество представляет собой сложную сеть производственной и потребительской деятельности, связанных друг с другом взаимодействующими цепочками поставок и торговли. Необязательно, что повышение энергоэффективности в одной точке сети (например, за счет рециркуляции или обмена ресурсами или реализации схем замкнутой экономики) приводит к общему повышению эффективности на уровне всей системы; Аналогичным образом, уменьшение одного вида воздействия (истощение энергии, истощение водных ресурсов, потребность в земле, выбросы и т.
, 2014). Так как городская территория представляет собой сложную систему, связанную с различными производственно-потребительскими цепочками поставок, исследование экологических, экономических и социальных аспектов с использованием общесистемной основы анализа для обеспечения устойчивости является ключевым вопросом для защиты благосостояния и разработки адекватных политических решений. .Даже если энергоэффективность и внедрение возобновляемых источников энергии, а также модели рециркуляции материалов (круговая экономика), кажется, развиваются быстро и эффективно, многое еще предстоит сделать. Общество представляет собой сложную сеть производственной и потребительской деятельности, связанных друг с другом взаимодействующими цепочками поставок и торговли. Необязательно, что повышение энергоэффективности в одной точке сети (например, за счет рециркуляции или обмена ресурсами или реализации схем замкнутой экономики) приводит к общему повышению эффективности на уровне всей системы; Аналогичным образом, уменьшение одного вида воздействия (истощение энергии, истощение водных ресурсов, потребность в земле, выбросы и т. д.)) не обязательно приводит к общему снижению воздействия системы. Следовательно, исследования городского метаболизма должны срочно подключиться к разработке решений в области устойчивого развития для социально-экологической динамики, описывающих и понимающих взаимозависимости и динамику городов и экосистем (John et al., 2019).
д.)) не обязательно приводит к общему снижению воздействия системы. Следовательно, исследования городского метаболизма должны срочно подключиться к разработке решений в области устойчивого развития для социально-экологической динамики, описывающих и понимающих взаимозависимости и динамику городов и экосистем (John et al., 2019).
Обсуждение, диалог, совместное управление и соответствующие процессы решения проблем, основанные на подходящей количественной и качественной информации, являются важными аспектами для разработки соответствующего процесса принятия решений для моделей городского метаболизма, которые восстанавливаются как по замыслу, так и по замыслу, чтобы для достижения целей устойчивости и благополучия.
Устойчивое развитие — это то, что произойдет в городах или вряд ли произойдет. Переход от риторики устойчивости к реализации сообщества и благополучия — это то, что не произойдет само по себе и не может быть решено с помощью линейных и детерминированных подходов. Подобно экосистемам, городам требуется оптимизация использования ресурсов во всех секторах и на всех уровнях, взаимосвязь между компонентами и процессами, сбалансированное сотрудничество между городскими и сельскими районами, модели восстановления и рециркуляции остатков и отходов, импульсные стратегии в соответствии с колеблющейся доступностью ресурсов, образовательные программы, ориентированные на более глубокое понимание сложных экологических ограничений, стратегии качественного роста вместо стремления к неограниченному и вряд ли устойчивому количественному росту.
Подобно экосистемам, городам требуется оптимизация использования ресурсов во всех секторах и на всех уровнях, взаимосвязь между компонентами и процессами, сбалансированное сотрудничество между городскими и сельскими районами, модели восстановления и рециркуляции остатков и отходов, импульсные стратегии в соответствии с колеблющейся доступностью ресурсов, образовательные программы, ориентированные на более глубокое понимание сложных экологических ограничений, стратегии качественного роста вместо стремления к неограниченному и вряд ли устойчивому количественному росту.
Авторские взносы
Все перечисленные авторы внесли существенный, прямой и интеллектуальный вклад в работу и одобрили ее к публикации.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Список литературы
Сеспедес Рестрепо, Дж. Д. и Моралес-Пинсон, Т. (2018).Городской метаболизм и устойчивость: прецеденты, генезис и перспективы исследования. Resour. Консерв. Рецикл . 131, 216–224. DOI: 10.1016 / j.resconrec.2017.12.023
Д. и Моралес-Пинсон, Т. (2018).Городской метаболизм и устойчивость: прецеденты, генезис и перспективы исследования. Resour. Консерв. Рецикл . 131, 216–224. DOI: 10.1016 / j.resconrec.2017.12.023
CrossRef Полный текст | Google Scholar
Цуй, X. (2018). Как города могут поддерживать устойчивость: библиометрический анализ городского метаболизма. Ecol. Инд . 93, 704–717. DOI: 10.1016 / j.ecolind.2018.05.056
CrossRef Полный текст | Google Scholar
Карри, П. К., Мусанго, Дж.К., и Мэй, Н. Д. (2017). Городской метаболизм: обзор со ссылкой на Кейптаун. Города 70, 91–110. DOI: 10.1016 / j.cities.2017.06.005
CrossRef Полный текст | Google Scholar
дель Мар Мартинес-Браво, М., Мартинес-дель-Рио, Х. и Антолин-Лопес, Р. (2019). Компромиссы между устойчивостью городов, загрязнением окружающей среды и пригодностью для жизни в европейских городах. J. Clean. Тов . 224, 651–660. DOI: 10.1016 / j.jclepro.2019.03. 110
110
CrossRef Полный текст | Google Scholar
Дийст, М., Worrell, E., Böcker, L., Brunner, P., Davoudi, S., Geertman, S., et al. (2018). Изучение городского метаболизма — к междисциплинарной перспективе. Resour. Консерв. Рецикл . 132, 190–203. DOI: 10.1016 / j.resconrec.2017.09.014
CrossRef Полный текст | Google Scholar
Фань, Дж. Л., Конг, Л. С., Ван, Х. и Чжан, X. (2019). Обзор взаимосвязи воды и энергии с точки зрения городского метаболизма. Ecol. Модель . 392, 128–136. DOI: 10.1016 / j.ecolmodel.2018.11.019
CrossRef Полный текст | Google Scholar
Хуанг, С. Л., Ли, К. Л., и Чен, К. В. (2006). Социально-экономический метаболизм в Тайване: неожиданный синтез против анализа материального потока. Resour. Консерв. Рецикл . 48, 166–196. DOI: 10.1016 / j.resconrec.2006.01.005
CrossRef Полный текст | Google Scholar
Джон Б., Людериц К., Ланг Д. Дж. И фон Верден Х. (2019). На пути к устойчивому метаболизму в городах. от понимания системы к трансформации системы. Ecol. Экон . 157, 402–414. DOI: 10.1016 / j.ecolecon.2018.12.007
от понимания системы к трансформации системы. Ecol. Экон . 157, 402–414. DOI: 10.1016 / j.ecolecon.2018.12.007
CrossRef Полный текст | Google Scholar
Одум, Х. Т. (1996). Экологический учет: принятие решений по чрезвычайным ситуациям и окружающей среде . Нью-Йорк, штат Нью-Йорк: Wiley-Indersciene.
Google Scholar
Организация Объединенных Наций (2018b). Мировые перспективы урбанизации: редакция 2018 г. .
Вэй, Х., Шэнхуэй, К., Яриме, М., Хашимото, С., и Манаги, С. (2015).Улучшение городского метаболизма исследования для устойчивой городской трансформации. Environ. Technol. Иннов . 4, 62–72. DOI: 10.1016 / j.eti.2015.04.004
CrossRef Полный текст | Google Scholar
Юн, Г., Фу, Дж., Саркис, Дж., И Сюэ, Б. (2012). На пути к национальной системе показателей замкнутой экономики в Китае: оценка и критический анализ. J. Clean. Прод . 23, 216. doi: 10.1016 / j.jclepro.2011.07.005
CrossRef Полный текст | Google Scholar
Йонг, Г. , Саркис, Дж., Улгиати, С., и Чжан, П. (2013). Измерение круговой экономики Китая. Наука 339, 1526–1527. DOI: 10.1126 / science.1227059
, Саркис, Дж., Улгиати, С., и Чжан, П. (2013). Измерение круговой экономики Китая. Наука 339, 1526–1527. DOI: 10.1126 / science.1227059
CrossRef Полный текст | Google Scholar
Чжан Ю., Вуа К. и Фатх Б. Д. (2018). Обзор пространственного анализа городского углеродного метаболизма Ecol . Модель . 371, 18–24. DOI: 10.1016 / j.ecolmodel.2018.01.005
CrossRef Полный текст
Зукаро, А., Рипа, М., Меллино, С., Асьоне, М., и Ульгиати, С. (2014). Показатели использования городских ресурсов и экологической результативности. Приложение декомпозиционного анализа. Ecol. Инд . 47, 16–25. DOI: 10.1016 / j.ecolind.2014.04.022
CrossRef Полный текст | Google Scholar
Метаболизм Метаболические пути — концепция, цель и примеры
Введение о том, что такое метаболизм Метаболический путь
Все химические реакции, происходящие внутри клетки, в совокупности называются клеточным метаболизмом.Если бы мы могли заглянуть внутрь любой клетки нашего тела, мы бы увидели, что это серия биохимических реакций, которые превращают молекулы субстрата в серию промежуточных продуктов метаболизма с образованием конечных продуктов. Промежуточный продукт ферментативной реакции известен как метаболит.
Метаболиты дополнительно модифицируются последовательностью химических реакций, катализируемых ферментами.
Концепция обмена веществ
Весь процесс питания состоит из двух частей. Прием пищи и использование пищи для получения энергии.В каждом живом организме, будь то прокариотическая бактерия или эукариотическая клетка, процесс питания одинаков. Понятие метаболических реакций относится к использованию пищи для получения энергии. Непереваренную пищу нужно утилизировать для переворота. Ключ к метаболизму — это питание, а цель — получение энергии.
Во время метаболизма биомолекулы, присутствующие в пище, используются для извлечения энергии из клетки. Образование другой молекулы вызвано преобразованием одного соединения.Например, белки, полученные из пищи, метаболизируются в аминокислоты, которые позже используются для синтеза другого белка. Метаболизм — это катализируемая ферментами реакция, которая обеспечивает биомолекулы, необходимые клетке для роста, поддержания и восстановления.
Пример метаболических путей:
Глюкоза окисляется с помощью кислорода с целью производства АТФ, аденозинтрифосфата в процессе клеточного дыхания.
Для правильного функционирования жизнедеятельности клеток молекулы АТФ используются клетками животных, поскольку они действуют как основной источник энергии для метаболизма.
Простые способы увеличить метаболизм
Солома содержит белки, которые повышают метаболизм в организме. Это называется термическим эффектом еды. Наибольший рост термического эффекта пищи наблюдается при потреблении белка. Употребление белков предохраняет нас от явления переедания.
Употребление большего количества холодной воды вместо сладких напитков может успешно помочь снизить потребление калорий. Выпивая 17 унций (0.5 литров воды увеличивает метаболизм в состоянии покоя на 10-30 процентов в течение примерно часа.
Поднятие тяжелых предметов помогает тренировать мышцы и бороться со снижением метаболизма.
Больше стойте — долгие часы сидения снижают метаболическую активность.
Употребление перца в пищу увеличивает метаболизм, так как перец содержит капсаицин, который ускоряет метаболизм.
Назначение метаболических путей
Для клеточной активности по выработке энергии.
Для преобразования пищи в строительные блоки для синтеза биомолекул, таких как углеводы, белки, липиды и нуклеиновые кислоты.
Для удаления отходов и токсичных продуктов.
Основные метаболические пути
Есть два основных типа метаболических путей.
Катаболические и анаболические.
Катаболические пути выделяют энергию, которая расщепляет молекулы на более простые молекулы.
Пример катаболического пути —
Цикл Кребса или цикл лимонной кислоты — в этом пути ацетат макроэлементов, таких как молекулы белков, жиров и углеводов, подвергается окислению.Конечный продукт — диоксид углерода.
[Изображение будет загружено в ближайшее время]
Гликолиз. Процесс, при котором организмы и растения накапливают и выделяют глюкозу и другую энергию сахара для образования высокоэнергетической молекулы, известен как аденозинтрифосфат (АТФ). АТФ называют «энергетической валютой жизни», потому что он хранит энергию, необходимую нам для повседневной работы. Катаболический путь помогает создавать энергию для различных видов деятельности.
Анаболические пути
Ферменты, участвующие в анаболическом пути или процессе биосинтеза, нуждаются в энергии для изменения или преобразования молекул в более сложные молекулы или макромолекулы.
Пример. Аминокислоты можно использовать для образования белка, двуокиси углерода и для производства сахара. Кроме того, нуклеиновые кислоты можно использовать для создания новых цепей ДНК.
Press-pulse: новая терапевтическая стратегия для метаболического лечения рака | Питание и обмен веществ
Arens NC, West ID. Пресс-импульс: общая теория массового вымирания? Палеобиология. 2008. 34 (4): 456–71.
Артикул Google ученый
Сейфрид TN, Флорес RE, Poff AM, D’Agostino DP. Рак как нарушение обмена веществ: значение для новой терапии. Канцерогенез. 2014; 35 (3): 515–27.
CAS PubMed Статья Google ученый
Sonnenschein C, Soto AM. Теория соматических мутаций канцерогенеза: почему ее следует отбросить и заменить. Mol Carcinog. 2000. 29 (4): 205–11.
CAS PubMed Статья Google ученый
Ханахан Д., Вайнберг, РА. Признаки рака: следующее поколение. Клетка. 2011; 144 (5): 646–74.
CAS PubMed Статья Google ученый
Сейфрид TN, Huysentruyt LC. О происхождении метастазов рака. Crit Rev Oncog. 2013; 18 (1–2): 43–73.
PubMed PubMed Central Статья Google ученый
Sporn MB. Война с раком. Ланцет.1996. 347 (9012): 1377–81.
CAS PubMed Статья Google ученый
Фидлер И.Дж. Патогенез метастазирования рака: пересмотр гипотезы «семя и почва». Нат Рев Рак. 2003. 3 (6): 453–8.
CAS PubMed Статья Google ученый
Лазебник Ю. Каковы признаки рака? Нат Рев Рак. 2010. 10 (4): 232–3.
CAS PubMed Статья Google ученый
Тарин Д. Взаимодействия клеток и тканей в канцерогенезе и метастазировании и их клиническое значение. Semin Cancer Biol. 2011. 21 (2): 72–82.
CAS PubMed Статья Google ученый
Сигель Р.Л., Миллер К.Д., Джемаль А. Статистика рака, 2017. CA Cancer J Clin. 2017; 67: 7–30.
Сигел Р., Найшадхам Д., Джемаль А. Статистика рака, 2013. CA Cancer J Clin. 2013; 63 (1): 11–30.
PubMed Статья Google ученый
Сейфрид TN. Рак как нарушение обмена веществ: о происхождении, лечении и профилактике рака. Хобокен: Уайли; 2012.
Книга. Google ученый
Martincorena I, Campbell PJ. Соматическая мутация в раковых и нормальных клетках. Наука. 2015; 349 (6255): 1483–9.
CAS PubMed Статья Google ученый
Сейфрид TN. Рак как заболевание митохондриального обмена.Front Cell Dev Biol. 2015; 3: 43.
PubMed PubMed Central Статья Google ученый
Александров Л.Б., Ник-Зайнал С., Клин Д.К., Апарисио С.А., Бехьяти С., Бьянкин А.В., Бигнелл Г.Р., Болли Н., Борг А., Борресен-Дейл А.Л. и др. Сигнатуры мутационных процессов при раке человека. Природа. 2013; 500 (7463): 415–21.
CAS PubMed PubMed Central Статья Google ученый
Фогельштейн Б., Пападопулос Н., Велкулеску В.Е., Чжоу С., Диас-младший Л.А., Кинзлер К.В. Пейзажи генома рака. Наука. 2013. 339 (6127): 1546–58.
CAS PubMed PubMed Central Статья Google ученый
Mazzocca A, Ferraro G, Misciagna G, Carr BI. Системный эволюционный подход к раку: гепатоканцерогенез как парадигма. Мед-гипотезы. 2016; 93: 132–7.
CAS PubMed Статья Google ученый
Bizzarri M, Cucina A. SMT и TOFT: почему и как они являются противоположными и несовместимыми парадигмами. Acta Biotheor. 2016; 64 (3): 221–39.
PubMed Статья Google ученый
Бейкер С.Г. Возникновение теории рака может привести к новым направлениям исследований. J Natl Cancer Inst. 2015; 107 (2).
Wishart DS. Рак — это генетическое заболевание или нарушение обмена веществ? EBioMedicine. 2015; 2 (6): 478–9.
PubMed PubMed Central Статья Google ученый
Бейкер С.Г., Крамер Б.С. Парадоксы канцерогенеза: новые возможности направлений исследований. BMC Рак. 2007; 7: 151.
PubMed PubMed Central Статья CAS Google ученый
Берджио Э., Мильоре Л. К системной парадигме канцерогенеза: связь эпигенетики и генетики. Mol Biol Rep. 2015; 42 (4): 777–90.
CAS PubMed Статья Google ученый
Soto AM, Sonnenschein C. Является ли системная биология многообещающим подходом к разрешению противоречий в исследованиях рака? Cancer Cell Int. 2012; 12 (1): 12.
PubMed PubMed Central Статья Google ученый
Braun AC. О происхождении раковых клеток. Am Sci. 1970. 58 (3): 307–20.
CAS PubMed Google ученый
Ник-Зайнал С., Дэвис Х., Стааф Дж., Рамакришна М., Глодзик Д., Зоу Х, Мартинкорена И., Александров Л.Б., Мартин С., Клин Д.К. и др.Пейзаж соматических мутаций в 560 полногеномных последовательностях рака груди. Природа. 2016; 534 (7605): 47–54.
CAS PubMed PubMed Central Статья Google ученый
Stratton MR. Изучение геномов раковых клеток: прогресс и перспективы. Наука. 2011. 331 (6024): 1553–8.
CAS PubMed Статья Google ученый
Cooke SL, Shlien A, Marshall J, Pipinikas CP, Martincorena I, Tubio JM, Li Y, Menzies A, Mudie L, Ramakrishna M, et al.Обработанные псевдогены приобретаются соматически во время развития рака. Nat Commun. 2014; 5: 3644.
PubMed PubMed Central Статья CAS Google ученый
Бартесаги С., Грациано В., Галавотти С., Энрикес Н.В., Беттс Дж., Саксена Дж., Миньери В., Дели А., Карлссон А., Мартинс Л. М. и др. Ингибирование окислительного метаболизма приводит к генетической инактивации и трансформации p53 в нервных стволовых клетках. Proc Natl Acad Sci U S A.2015; 112 (4): 1059–64.
CAS PubMed PubMed Central Статья Google ученый
Пачини Н., Борзиани Ф. Онкостатически-цитопротективный эффект мелатонина и других биоактивных молекул: общая цель митохондриального дыхания. Int J Mol Sci. 2016; 17 (3): 341.
PubMed PubMed Central Статья Google ученый
Ким А. Митохондрии в энергетическом обмене рака: виновники или сторонние наблюдатели? Toxicol Res.2015; 31 (4): 323–30.
CAS PubMed PubMed Central Статья Google ученый
Варбург О. О происхождении раковых клеток. Наука. 1956; 123 (3191): 309–14.
CAS PubMed Статья Google ученый
Варбург О. О нарушении дыхания в раковых клетках. Наука. 1956; 124: 269–70.
CAS PubMed Google ученый
Путиньяни Л., Раффа С., Песколидо Р., Аймати Л., Синьор Ф, Торриси М. Р., Грамматико П. Изменение уровней экспрессии системы окислительного фосфорилирования (OXPHOS) в митохондриях клеток рака молочной железы. Лечение рака груди Res. 2008. 110 (3): 439–52.
CAS PubMed Статья Google ученый
Dienel GA, Cruz NF. Аэробный гликолиз во время активации мозга: адренергическая регуляция и влияние норадреналина на метаболизм астроцитов.J Neurochem. 2016; 138 (1): 14–52.
CAS PubMed Статья Google ученый
Ракер Э. История эффекта Пастера и его патобиология. Mol Cell Biochem. 1974. 5 (1–2): 17–23.
CAS PubMed Статья Google ученый
Варбург О. Метаболизм опухолей. Нью-Йорк: Ричард Р. Смит; 1931.
Google ученый
Сейфрид TN. Варбургский спор. В: Рак как заболевание обмена веществ: происхождение, лечение и профилактика рака. edn. Хобокен: Уайли; 2012. с. 107–17.
Глава Google ученый
Зу XL, Гуппи М. Метаболизм рака: факты, фантастика и вымысел. Biochem Biophys Res Commun. 2004. 313 (3): 459–65.
CAS PubMed Статья Google ученый
Коппенол WH, Bounds PL, Dang CV. Вклад Отто Варбурга в современные концепции метаболизма рака. Нат Рев Рак. 2011. 11 (5): 325–37.
CAS PubMed Статья Google ученый
Poff AM, Ari C, Seyfried TN, D’Agostino DP. Кетогенная диета и гипербарическая кислородная терапия продлевают выживаемость мышей с системным метастатическим раком. PLoS One. 2013; 8 (6): e65522.
CAS PubMed PubMed Central Статья Google ученый
Кибиш М.А., Хан Х, Ченг Х., Сейфрид Теннесси. Ростовая среда in vitro вызывает аномалии липидомных и электронных транспортных цепей в митохондриях из неканцерогенных астроцитов и опухолей головного мозга. ASN Neuro. 2009; 1 (3): e00011.
PubMed PubMed Central Статья CAS Google ученый
Диас-Руис Р., Ригуле М., Девин А. Эффекты Варбурга и Крэбтри: о происхождении энергетического метаболизма раковых клеток и подавления глюкозы дрожжами.Biochim Biophys Acta. 2011; 1807 (6): 568–76.
CAS PubMed Статья Google ученый
Лезнев Е.И., Попова И.И., Лавровская В.П., Евтодиенко Ю.В. Сравнение скорости потребления кислорода в минимально трансформированных клетках BALB / 3 T3 и трансформированных вирусом клетках 3T3B-SV40. Биохимия (Москва). 2013. 78 (8): 904–8.
CAS Статья Google ученый
Hall A, Meyle KD, Lange MK, Klima M, Sanderhoff M, Dahl C, Abildgaard C, Thorup K, Moghimi SM, Jensen PB, et al.Дисфункциональное окислительное фосфорилирование делает клетки злокачественной меланомы зависимыми от гликолиза, управляемого онкогеном V600EBRAF. Oncotarget. 2013; 4 (4): 584–99.
PubMed PubMed Central Статья Google ученый
Сейфрид Т.Н. Нормально ли дыхание раковых клеток? В: Рак как заболевание обмена веществ: происхождение, лечение и профилактика рака. edn. Хобокен: Уайли; 2012. с. 119–32.
Глава Google ученый
Хочачка П.В., Сомеро Г.Н. Биохимическая адаптация: механизм и процесс физиологической эволюции. Нью-Йорк: Oxford Press; 2002.
Google ученый
Ramanathan A, Wang C, Schreiber SL. Пертурбационное профилирование клеточной модели онкогенеза с использованием метаболических измерений. Proc Natl Acad Sci U S. A. 2005; 102 (17): 5992–7.
CAS PubMed PubMed Central Статья Google ученый
Arcos JC, Тисон MJ, Gosch HH, Fabian JA. Последовательные изменения в транспорте электронов внутренней и внешней мембраны митохондрий и в контроле дыхания во время кормления аминоазокрасителями; стабильность фосфорилирования. Корреляция с изменениями набухания-сокращения и порогом туморогенеза. Cancer Res. 1969. 29 (6): 1298–305.
CAS PubMed Google ученый
Suarez RK, Lighton JR, Brown GS, Mathieu-Costello O.Митохондриальное дыхание в летных мышцах колибри. Proc Natl Acad Sci U S. A. 1991; 88 (11): 4870–3.
CAS PubMed PubMed Central Статья Google ученый
Burk D, Schade AL. О нарушении дыхания в раковых клетках. Наука. 1956; 124 (3215): 270–2.
CAS PubMed Google ученый
Smith AE, Kenyon DH. Объединяющая концепция канцерогенеза и его терапевтических последствий.Онкология. 1973; 27 (5): 459–79.
CAS PubMed Статья Google ученый
Colowick SP. Состояние теории гликолиза и дыхания в опухолях Варбурга. Q Rev Biol. 1961; 36: 273–6.
Артикул Google ученый
Ху И, Лу В., Чен Дж, Ван П., Чен З, Чжоу Ю., Огасавара М., Трачутхэм Д., Фенг Л., Пеликано Х и др. Трансформация K-ras (G12V) приводит к дисфункции митохондрий и метаболическому переключению с окислительного фосфорилирования на гликолиз.Cell Res. 2012. 22 (2): 399–412.
CAS PubMed Статья Google ученый
Куэзва Дж. М., Чен Дж., Алонсо А. М., Исидоро А., Мисек Д. Е., Ханаш С. М., Бир Д. Г.. Биоэнергетическая характеристика аденокарциномы легких является молекулярным маркером диагностики и прогноза рака. Канцерогенез. 2004. 25 (7): 1157–63.
CAS PubMed Статья Google ученый
Ferreira LM. Метаболизм рака: эффект Варбурга сегодня. Опыт Мол Патол. 2010. 89 (3): 372–80.
CAS PubMed Статья Google ученый
Сейфрид TN, Shelton LM. Рак как нарушение обмена веществ. Нутр Метаб (Лондон). 2010; 7 (1): 7.
Артикул CAS Google ученый
Poff AM, Ari C, Arnold P, Seyfried TN, D’Agostino DP. Добавление кетонов снижает жизнеспособность опухолевых клеток и продлевает выживаемость мышей с метастатическим раком.Int J Cancer. 2014; 135 (7): 1711–20.
CAS PubMed PubMed Central Статья Google ученый
Pedersen PL. Варбург, я и гексокиназа 2: многочисленные открытия ключевых молекулярных событий, лежащих в основе одного из наиболее распространенных фенотипов рака, «эффекта Варбурга», то есть повышенного гликолиза в присутствии кислорода. J Bioenerg Biomembr. 2007. 39 (3): 211–22.
CAS PubMed Статья Google ученый
Варбург О. Пересмотренные лекции Линдау: первопричина рака и профилактика — части 1 и 2. В: Линдау Б.Д., редактор. Встреча лауреатов Нобелевской премии. Боденское озеро: К. Трильч; 1969. с. 1–9. http://www.hopeforcancer.com/OxyPlus.htm.
Google ученый
Ракер Э. Биоэнергетика и проблема опухолевого роста. Am Sci. 1972; 60 (1): 56–63.
CAS PubMed Google ученый
Вайнхаус С. Гипотеза Варбурга пятьдесят лет спустя. Z Krebsforsch Klin Onkol Cancer Res Clin Oncol. 1976. 87 (2): 115–26.
CAS PubMed Статья Google ученый
Марин-Валенсия I, Ян К., Машимо Т., Чо С., Пэк Х., Ян XL, Раджагопалан К.Н., Мэдди М., Вемиредди В., Чжао З. и др. Анализ метаболизма опухолей показывает окисление митохондриальной глюкозы в генетически разнообразных глиобластомах человека в мозге мышей in vivo.Cell Metab. 2012. 15 (6): 827–37.
CAS PubMed PubMed Central Статья Google ученый
Сейфрид Т.Н. Нарушение функции дыхания в раковых клетках. В: Рак как заболевание обмена веществ: происхождение, лечение и профилактика рака. edn. Хобокен: Уайли; 2012. с. 73–105.
Глава Google ученый
Lichtor T, Dohrmann GJ. Респираторные паттерны при опухолях головного мозга человека.Нейрохирургия. 1986. 19 (6): 896–9.
CAS PubMed Статья Google ученый
Сейфрид Т.Н., Мукерджи П. Ориентация на энергетический метаболизм при раке мозга: обзор и гипотеза. Нутр Метаб (Лондон). 2005; 2:30.
Артикул CAS Google ученый
Vander Heiden MG, Cantley LC, Thompson CB. Понимание эффекта Варбурга: метаболические потребности пролиферации клеток.Наука. 2009. 324 (5930): 1029–33.
CAS PubMed PubMed Central Статья Google ученый
Cuezva JM, Krajewska M, de Heredia ML, Krajewski S, Santamaria G, Kim H, Zapata JM, Marusawa H, Chamorro M, Reed JC. Биоэнергетическая подпись рака: маркер прогрессирования опухоли. Cancer Res. 2002. 62 (22): 6674–81.
CAS PubMed Google ученый
Pedersen PL. Митохондрии опухолей и биоэнергетика раковых клеток. Prog Exp Tumor Res. 1978; 22: 190–274.
CAS PubMed Статья Google ученый
Мортон Р., Каннингем С., Шут Р., Уэйт М., Миллер Н., Моррис ХП. Изменение функции митохондрий и липидного состава при гепатоме Моррис 7777. Cancer Res. 1976; 36 (9 ч.1): 3246–54.
CAS PubMed Google ученый
Schild L, Lendeckel U, Gardemann A, Wiswedel I, Schmidt CA, Wolke C, Walther R, Grabarczyk P, Busemann C. Состав молекулярных видов кардиолипина коррелирует с пролиферацией лимфоцитов. Exp Biol Med. 2012. 237 (4): 372–9.
CAS Статья Google ученый
Sapandowski A, Stope M, Evert K, Evert M, Zimmermann U, Peter D, Page I, Burchardt M, Schild L. Состав кардиолипина коррелирует с пролиферацией клеток рака простаты.Mol Cell Biochem. 2015; 410 (1–2): 175–85.
CAS PubMed Статья Google ученый
Canuto RA, Biocca ME, Muzio G, Dianzani MU. Жирнокислотный состав фосфолипидов в митохондриях и микросомах во время канцерогенеза диэтилнитрозамина в печени крыс. Cell Biochem Funct. 1989. 7 (1): 11–9.
CAS PubMed Статья Google ученый
Кибиш М.А., Хан Х, Ченг Х, Чуанг Дж.Х., Сейфрид Теннесси. Кардиолипин и аномалии цепи транспорта электронов в митохондриях опухолей головного мозга мышей: липидомные данные, подтверждающие теорию рака Варбурга. J Lipid Res. 2008. 49 (12): 2545–56.
CAS PubMed PubMed Central Статья Google ученый
Пейта Л., Ярнуен К., Пино М., Гимараес С., де Баррос Дж. П. П., Шевалье С., Дюма Дж. Ф., Майо Ф., Хэтч Дж. М., Лойер П. и др.Пониженное содержание кардиолипина снижает емкость дыхательной цепи и увеличивает синтез АТФ в клетках HepaRG человека. Biochim Biophys Acta. 2016; 4: 443–53.
Артикул CAS Google ученый
Kiebish MA, Han X, Cheng H, Seyfried TN. Митохондриальный липидом и изменения цепи переноса электронов в неметастатических и метастатических опухолях головного мозга мышей. J Neurochem. 2008; 104 Приложение 1: 37–8.
Google ученый
Claypool SM, Келер CM. Сложность кардиолипина в здоровье и болезни. Trends Biochem Sci. 2012; 37 (1): 32–41.
CAS PubMed Статья Google ученый
Ren M, Phoon CK, Schlame M. Метаболизм и функция митохондриального кардиолипина. Prog Lipid Res. 2014; 55: 1–16.
CAS PubMed Статья Google ученый
Чинопулос К.В какую сторону поворачивается цикл лимонной кислоты во время гипоксии? Решающая роль комплекса альфа-кетоглутаратдегидрогеназы. J Neurosci Res. 2013. 91 (8): 1030–43.
CAS PubMed Статья Google ученый
Peiris-Pages M, Martinez-Outschoorn UE, Pestell RG, Sotgia F, Lisanti MP. Метаболизм раковых стволовых клеток. Рак молочной железы Res. 2016; 18 (1): 55.
PubMed PubMed Central Статья Google ученый
Дейтон Р.Ф., Ле Бихан Т., Мартин С.Ф., Герт А.М., МакКуллох М., Эдгар Дж. М., Керр Л. Е., Уиттл И. Р., МакКаллох Дж. Взаимодействия между митохондриальными белками, измененными в глиобластоме. J Neuro-Oncol. 2014. 118 (2): 247–56.
CAS Статья Google ученый
Arismendi-Morillo GJ, Castellano-Ramirez AV. Ультраструктурная митохондриальная патология в астроцитарных опухолях человека: потенциальные последствия терапевтических стратегий. J Electron Microsc (Токио).2008. 57 (1): 33–9.
Артикул Google ученый
Schmitt S, Schulz S, Schropp EM, Eberhagen C, Simmons A, Beisker W., Aichler M, Zischka H. Зачем сравнивать абсолютное количество митохондрий. Митохондрия. 2014; 19 Pt A: 113–23.
PubMed Статья CAS Google ученый
Verschoor ML, Ungard R, Harbottle A, Jakupciak JP, Parr RL, Singh G.Митохондрии и рак: прошлое, настоящее и будущее. Biomed Res Int. 2013; 2013: 612369.
CAS PubMed PubMed Central Статья Google ученый
Сринивасан С., Гуха М., Донг Д.В., Уилан К.А., Рутел Г., Утикадо Ю., Нацуго С.С., Накагава Х., Авадхани Н.Г. Нарушение функции цитохром-с-оксидазы вызывает эффект Варбурга и метаболическое перепрограммирование. Онкоген. 2015; 35: 1585–95.
Sriskanthadevan S, Jeyaraju DV, Chung TE, Prabha S, Xu W., Skrtic M, Jhas B, Hurren R, Gronda M, Wang X, et al.Клетки AML обладают низкой резервной способностью в дыхательной цепи, что делает их восприимчивыми к окислительному метаболическому стрессу. Кровь. 2015; 125 (13): 2120–30.
CAS PubMed PubMed Central Статья Google ученый
Levine AJ, Puzio-Kuter AM. Контроль метаболического переключателя при раке онкогенами и генами-супрессорами опухолей. Наука. 2010. 330 (6009): 1340–4.
CAS PubMed Статья Google ученый
Kaipparettu BA, Ma Y, Park JH, Lee TL, Zhang Y, Yotnda P, Creighton CJ, Chan WY, Wong LJ. Перекрестные помехи от незлокачественных митохондрий могут подавлять опухолевые свойства метастатических клеток, подавляя онкогенные пути. PLoS One. 2013; 8 (5): e61747.
CAS PubMed PubMed Central Статья Google ученый
Сейфрид Т.Н. Митохондрии: лучший супрессор опухолей. В: Рак как заболевание обмена веществ: происхождение, лечение и профилактика рака.edn. Хобокен: Уайли; 2012. с. 195–205.
Глава Google ученый
Kloc M, Li XC, Ghobrial RM. Ответственны ли макрофаги за метастазирование рака? J Immuno Biol. 2016; 1: 1.
Google ученый
Павелек Ю.М., Чакраборти А.К. Слияние опухолевых клеток с клетками костного мозга: объединяющее объяснение метастазов. Нат Рев Рак. 2008. 8 (5): 377–86.
CAS PubMed Статья Google ученый
Бастида-Руис Д., Ван Хусен К., Коэн М.: Темная сторона слияния клеток. Int J Mol Sci. 2016, 17 (5). DOI: 10.3390 / ijms17050638
Сейфрид TN. Митохондриальная респираторная дисфункция и внехромосомное происхождение рака. В: Рак как заболевание обмена веществ: происхождение, лечение и профилактика рака. edn. Хобокен: Уайли; 2012 г.п. 253–9.
Глава Google ученый
Nemeth B, Doczi J, Csete D, Kacso G, Ravasz D, Adams D, Kiss G, Nagy AM, Horvath G, Tretter L, et al. Отмена митохондриального фосфорилирования на уровне субстрата итаконовой кислотой, продуцируемой LPS-индуцированной экспрессией Irg1 в клетках мышиного клона макрофагов. FASEB J. 2016; 30 (1): 286–300.
CAS PubMed Статья Google ученый
Сейфрид TN. Является ли митохондриальная ферментация глутамина недостающим звеном в метаболической теории рака? В: Рак как заболевание обмена веществ: происхождение, лечение и профилактика рака. edn. Хобокен: Уайли; 2012. с. 133–44.
Глава Google ученый
Чинопулос С., Геренсер А.А., Манди М., Матэ К., Торочик Б., Докзи Дж., Туриак Л., Кисс Г., Конрад С., Вайда С. и др. Прямое действие адениннуклеотидной транслоказы во время обращения F0F1-АТФазы: критическая роль фосфорилирования на уровне субстрата матрикса.FASEB J. 2010; 24 (7): 2405–16.
CAS PubMed PubMed Central Статья Google ученый
Филлипс Д., Апонте А.М., Французский SA, Chess DJ, Balaban RS. Сукцинил-КоА-синтетаза является фосфатной мишенью для активации митохондриального метаболизма. Биохимия. 2009. 48 (30): 7140–9.
CAS PubMed PubMed Central Статья Google ученый
Schwimmer C, Lefebvre-Legendre L, Rak M, Devin A, Slonimski PP, di Rago JP, Rigoulet M. Повышение фосфорилирования митохондриального уровня субстрата может спасти респираторный рост дрожжей с дефицитом АТФ-синтазы. J Biol Chem. 2005. 280 (35): 30751–9.
CAS PubMed Статья Google ученый
Kiss G, Konrad C, Pour-Ghaz I, Mansour JJ, Nemeth B, Starkov AA, Adam-Vizi V, Chinopoulos C. Митохондриальные диафоразы как доноры NAD (+) для сегментов цикла лимонной кислоты, которые поддерживают фосфорилирование на уровне субстрата с образованием АТФ во время респираторного ингибирования.FASEB J. 2014; 28 (4): 1682–97.
CAS PubMed PubMed Central Статья Google ученый
Ньюсхолм Е.А., Совет М. Применение логики метаболического контроля для использования топлива и его значение в опухолевых клетках. Adv Enzyme Regul. 1991; 31: 225–46.
CAS PubMed Статья Google ученый
DeBerardinis RJ, Cheng T. Q: различные функции глутамина в метаболизме, клеточной биологии и раке.Онкоген. 2010. 29 (3): 313–24.
CAS PubMed Статья Google ученый
Юнева М. Обнаружение «ахиллесовой пяты» рака: роль метаболизма глюкозы и глутамина в выживании трансформированных клеток. Клеточный цикл. 2008. 7 (14): 2083–9.
CAS PubMed Статья Google ученый
Медина MA. Глютамин и рак. J Nutr. 2001; 131 (9 доп.): 2539–2542S.обсуждение 2550С-2531С.
Google ученый
Хуанг В., Чой В., Чен Й, Чжан К., Дэн Х, Хе В., Ши Ю. Предполагаемая роль глутамина в росте раковых клеток за счет кислотной устойчивости. Cell Res. 2013; 23 (5): 724–7.
CAS PubMed PubMed Central Статья Google ученый
Накашима Р.А., Пагги М.Г., Педерсен ПЛ. Вклад гликолиза и окислительного фосфорилирования в продукцию аденозин-5′-трифосфата в клетках гепатомы AS-30D.Cancer Res. 1984. 44 (12 Pt 1): 5702–6.
CAS PubMed Google ученый
Ta NL, Сейфрид Теннесси. Влияние сыворотки и гипоксии на включение [(14) C] -D-глюкозы или [(14) C] -L-глутамина в липиды и лактат в клетках глиобластомы мыши. Липиды. 2015; 50 (12): 1167–84.
CAS PubMed Статья Google ученый
Portais JC, Voisin P, Merle M, Canioni P.Метаболизм глюкозы и глутамина в клетках глиомы С6 изучен методом ЯМР углерода 13. Биохимия. 1996. 78 (3): 155–64.
CAS PubMed Статья Google ученый
Скотт Д.А., Ричардсон А.Д., Филипп Ф.В., Кнутцен, Калифорния, Чианг Г.Г., Ронаи З.А., Остерман А.Л., Смит Дж.В. Сравнительное профилирование метаболического потока клеточных линий меланомы: за пределами эффекта Варбурга. J Biol Chem. 2011. 286 (49): 42626–34.
CAS PubMed PubMed Central Статья Google ученый
Shelton LM, Huysentruyt LC, Сейфрид Теннесси. Нацеливание на глутамин подавляет системные метастазы в модели опухоли мышей VM-M3. Int J Cancer. 2010. 127 (10): 2478–85.
CAS PubMed PubMed Central Статья Google ученый
Писаренко О.И., Соломатина Е.С., Иванов В.Е., Студнева И.М., Капелко В.И., Смирнов В.Н. О механизме повышенного образования АТФ в гипоксическом миокарде под действием глутаминовой кислоты. Basic Res Cardiol.1985. 80 (2): 126–34.
CAS PubMed Статья Google ученый
Weinberg JM, Venkatachalam MA, Roeser NF, Nissim I. Митохондриальная дисфункция во время гипоксии / реоксигенации и ее коррекция анаэробным метаболизмом промежуточных продуктов цикла лимонной кислоты. Proc Natl Acad Sci U S. A. 2000; 97 (6): 2826–31.
CAS PubMed PubMed Central Статья Google ученый
Tannahill GM, Curtis AM, Adamik J, Palsson-McDermott EM, McGettrick AF, Goel G, Frezza C, Bernard NJ, Kelly B, Foley NH, et al. Сукцинат является воспалительным сигналом, который индуцирует IL-1beta через HIF-1alpha. Природа. 2013. 496 (7444): 238–42.
CAS PubMed PubMed Central Статья Google ученый
Хочачка П.В., Оуэн Т.Г., Аллен Дж.Ф., Уиттоу Г.К. Множественные конечные продукты анаэробиоза у ныряющих позвоночных.Comp Biochem Physiol B. 1975; 50 (1): 17–22.
CAS PubMed Google ученый
Кинг А., Селак М.А., Готлиб Э. Сукцинатдегидрогеназа и фумаратгидратаза: связь митохондриальной дисфункции и рака. Онкоген. 2006. 25 (34): 4675–82.
CAS PubMed Статья Google ученый
Marsh J, Mukherjee P, Seyfried TN. Akt-зависимые проапоптотические эффекты ограничения диеты на поздней стадии лечения астроцитомы мыши с дефицитом фосфатазы и гомолога тензина / комплекса туберозного склероза 2.Clin Cancer Res. 2008. 14 (23): 7751–62.
CAS PubMed Статья Google ученый
Semenza GL. HIF-1 опосредует эффект Варбурга при светлоклеточном раке почек. J Bioenerg Biomembr. 2007. 39 (3): 231–4.
CAS PubMed Статья Google ученый
Чжан Х., Гао П., Фукуда Р., Кумар Г., Кришнамачари Б., Зеллер К.И., Данг К.В., Семенза Г.Л.HIF-1 ингибирует митохондриальный биогенез и клеточное дыхание при почечно-клеточной карциноме с дефицитом VHL путем репрессии активности C-MYC. Раковая клетка. 2007. 11 (5): 407–20.
CAS PubMed Статья Google ученый
Comerford SA, Huang Z, Du X, Wang Y, Cai L, Witkiewicz AK, Walters H, Tantawy MN, Fu A, Manning HC, et al. Ацетатная зависимость опухолей. Клетка. 2014. 159 (7): 1591–602.
CAS PubMed PubMed Central Статья Google ученый
Хосиос AM, Вандер Хайден MG. Метаболизм ацетата в раковых клетках. Рак и метаболизм. 2014; 2 (1): 27.
Артикул Google ученый
Ballard FJ. Поставка и использование ацетата у млекопитающих. Am J Clin Nutr. 1972: 25 (8): 773–9.
CAS PubMed Google ученый
Jaworski DM, Namboodiri AM, Moffett JR. Ацетат как метаболический и эпигенетический модификатор терапии рака.J Cell Biochem. 2015; 117: 574–88.
Huysentruyt LC, Сейфрид Теннесси. Перспективы мезенхимального происхождения метастатического рака. Раковые метастазы Rev.2010; 29 (4): 695–707.
PubMed PubMed Central Статья Google ученый
Pawelek JM. Слияние опухолевых клеток как источник миелоидных признаков рака. Ланцет Онкол. 2005; 6 (12): 988–93.
CAS PubMed Статья Google ученый
Ерш MR, Перт CB. Мелкоклеточный рак легкого: антигены, специфичные для макрофагов, предполагают происхождение гемопоэтических стволовых клеток. Наука. 1984. 225 (4666): 1034–6.
CAS PubMed Статья Google ученый
Пауэлл А.Е., Андерсон Е.С., Дэвис П.С., Шелк А.Д., Пелц С., Импей С., Вонг М.Х. Слияние эпителиальных клеток кишечника и макрофагов в контексте рака приводит к ядерному репрограммированию. Cancer Res. 2011; 71 (4): 1497–505.
CAS PubMed PubMed Central Статья Google ученый
Yeh MH, Chang YH, Tsai YC, Chen SL, Huang TS, Chiu JF, Ch’ang HJ. Макрофаги, полученные из костного мозга, сливаются со стромальными клетками кишечника и способствуют развитию хронического фиброза после облучения. Радиотренажер Oncol. 2016; 119 (2): 250–8.
CAS PubMed Статья Google ученый
Abodief WT, Dey P, Al-Hattab O.Клеточный каннибализм при протоковом раке груди. Цитопатология. 2006. 17 (5): 304–5.
CAS PubMed Статья Google ученый
Фейс С. Каннибализм: способ подпитаться метастатическими опухолями. Cancer Lett. 2007. 258 (2): 155–64.
CAS PubMed Статья Google ученый
Луджини Л., Матаррезе П., Тинари А., Лозупоне Ф., Федеричи С., Иесси Е., Джентиле М., Лучиани Ф., Пармиани Дж., Риволтини Л. и др.Каннибализм живых лимфоцитов метастатическими, но не первичными клетками меланомы человека. Cancer Res. 2006. 66 (7): 3629–38.
CAS PubMed Статья Google ученый
Матаррезе П., Чиарло Л., Тинари А., Пьячентини М., Малорни В. Ксено-каннибализм как обострение самоканнибализма: возможная плодотворная стратегия выживания раковых клеток. Curr Pharm Des. 2008. 14 (3): 245–52.
CAS PubMed Статья Google ученый
Гупта К., Дей П. Клеточный каннибализм: диагностический маркер злокачественности. Diagn Cytopathol. 2003. 28 (2): 86–7.
PubMed Статья Google ученый
Кодзима С., Секин Х., Фукуи И., Охшима Х. Клиническое значение «каннибализма» в цитологии мочи при раке мочевого пузыря. Acta Cytol. 1998. 42 (6): 1365–9.
CAS PubMed Статья Google ученый
Shelton LM. Ориентация на энергетический метаболизм при раке мозга. Честнат-Хилл: Бостонский колледж; 2010.
Google ученый
Камфорст Дж. Дж., Нофал М., Коммиссо С., Хакетт С. Р., Лу В., Грабока Е., Вандер Хайден М. Г., Миллер Г., Дребин Дж. А., Бар-Саги Д. и др. Опухоли рака поджелудочной железы человека бедны питательными веществами, и опухолевые клетки активно поглощают внеклеточный белок. Cancer Res. 2015; 75 (3): 544–53.
CAS PubMed PubMed Central Статья Google ученый
Лу Дж, Шарма Л.К., Бай Ю. Влияние мутаций митохондриальной ДНК и митохондриальной дисфункции в онкогенезе. Cell Res. 2009. 19 (7): 802–15.
CAS PubMed PubMed Central Статья Google ученый
Yang D, Wang MT, Tang Y, Chen Y, Jiang H, Jones TT, Rao K, Brewer GJ, Singh KK, Nie D. Нарушение митохондриального дыхания в фибробластах мыши онкогенным H-RAS (Q61L ). Cancer Biol Ther. 2010. 9 (2): 122–33.
CAS PubMed PubMed Central Статья Google ученый
Smiraglia DJ, Kulawiec M, Bistulfi GL, Gupta SG, Singh KK. Новая роль митохондрий в регуляции эпигенетической модификации ядра. Cancer Biol Ther. 2008. 7 (8): 1182–90.
CAS PubMed PubMed Central Статья Google ученый
Делсайт Р.Л., Расмуссен Л.Дж., Расмуссен А.К., Кален А., Госвами П.С., Сингх К.К.Нарушение митохондрий сопровождается нарушением окислительной репарации ДНК в ядре. Мутагенез. 2003. 18 (6): 497–503.
CAS PubMed Статья Google ученый
Кулавец М., Сафина А., Десуки М.М., Стилл И., Мацуи С.И., Бакин А., Сингх К.К. Опухолевая трансформация эпителиальных клеток молочной железы человека, вызванная истощением митохондриальной ДНК. Cancer Biol Ther. 2008. 7 (11): 1732–43.
CAS PubMed PubMed Central Статья Google ученый
Расмуссен А.К., Чаттерджи А., Расмуссен Л.Дж., Сингх К.К. Митохондриально-опосредованный фенотип ядерных мутаторов у Saccharomyces cerevisiae. Nucleic Acids Res. 2003. 31 (14): 3909–17.
CAS PubMed PubMed Central Статья Google ученый
Чандра Д., Сингх К.К. Генетическое понимание дефекта OXPHOS и его роли в развитии рака. Biochim Biophys Acta. 2011; 1807 (6): 620–5.
CAS PubMed Статья Google ученый
Veatch JR, McMurray MA, Nelson ZW, Gottschling DE. Дисфункция митохондрий приводит к нестабильности ядерного генома из-за дефекта кластера железо-сера. Клетка. 2009. 137 (7): 1247–58.
PubMed PubMed Central Статья Google ученый
Сампер Э., Николлс Д.Г., Мелов С. Митохондриальный окислительный стресс вызывает хромосомную нестабильность эмбриональных фибробластов мыши. Ячейка старения. 2003. 2 (5): 277–85.
CAS PubMed Статья Google ученый
Seoane M, Mosquera-Miguel A, Gonzalez T, Fraga M, Salas A, Costoya JA. Митохондриальный геном — это «генетическое убежище» во время онкогенного процесса. PLoS One. 2011; 6 (8): e23327.
CAS PubMed PubMed Central Статья Google ученый
Minocherhomji S, Tollefsbol TO, Singh KK. Митохондриальная регуляция эпигенетики и ее роль в заболеваниях человека. Эпигенетика. 2012; 7 (4): 326–34.
CAS PubMed PubMed Central Статья Google ученый
Veech RL. Терапевтическое значение кетоновых тел: эффекты кетоновых тел при патологических состояниях: кетоз, кетогенная диета, окислительно-восстановительные состояния, инсулинорезистентность и митохондриальный метаболизм. Простагландины Leukot Essent Fatty Acids. 2004. 70 (3): 309–19.
CAS PubMed Статья Google ученый
Sabharwal SS, Schumacker PT. Митохондриальные АФК при раке: инициаторы, усилители или ахиллесова пята? Нат Рев Рак.2014. 14 (11): 709–21.
CAS PubMed PubMed Central Статья Google ученый
Клауниг Ю.Е., Камендулис Л.М., Хочевар Б.А. Окислительный стресс и окислительное повреждение в канцерогенезе. Toxicol Pathol. 2010. 38 (1): 96–109.
CAS PubMed Статья Google ученый
Сент-Дьердьи А. Живое состояние и рак. Proc Natl Acad Sci U S A.1977; 74 (7): 2844–7.
CAS PubMed PubMed Central Статья Google ученый
Кэрнс Дж. Происхождение рака у человека. Природа. 1981. 289 (5796): 353–7.
CAS PubMed Статья Google ученый
Мукерджи С. Император всех болезней: биография рака (страницы 285, 303, 333, 342). Нью-Йорк: Скрибнер; 2010.
Google ученый
Поттс Р. Экологические гипотезы эволюции гомининов. Am J Phys Anthropol. 1998; Дополнение 27: 93–136.
CAS PubMed Статья Google ученый
Поттс Р. Происхождение человечества: последствия экологической нестабильности. Нью-Йорк: William Morrow & Co., Inc.; 1996.
Google ученый
Поттс Р. Сложность адаптации в эволюции человека.В: Гудман М., Моффат А.С., редакторы. Исследование происхождения человека. edn. Кембридж: Американская академия искусств и наук; 2002. с. 33–57.
Google ученый
Сейфрид Т.Н. Ничто в биологии рака не имеет смысла, кроме как в свете эволюции. В: Рак как заболевание обмена веществ: происхождение, лечение и профилактика рака. edn. Хобокен: Уайли; 2012. с. 261–75.
Глава Google ученый
Дарвин К. О происхождении видов путем естественного отбора или о сохранении привилегированных рас в борьбе за жизнь. Лондон: Джон Мерри; 1859.
Google ученый
Моисеева О., Бурдо В., Ру А., Дешен-Симар Х, Фербейр Г. Дисфункция митохондрий способствует старению, индуцированному онкогенами. Mol Cell Biol. 2009. 29 (16): 4495–507.
CAS PubMed PubMed Central Статья Google ученый
de Groof AJ, te Lindert MM, van Dommelen MM, Wu M, Willemse M, Smift AL, Winer M, Oerlemans F, Pluk H, Fransen JA, et al. Повышенная активность OXPHOS предшествует увеличению скорости гликолиза в фибробластах, трансформированных H-RasV12 / E1A, которые развивают фенотип Варбурга. Молочный рак. 2009; 8: 54.
PubMed PubMed Central Статья CAS Google ученый
Матоба С., Кан Дж. Г., Патино В. Д., Врагг А., Бём М., Гаврилова О., Херли П. Дж., Бунц Ф., Хван П. М..p53 регулирует митохондриальное дыхание. Наука. 2006. 312 (5780): 1650–3.
CAS PubMed Статья Google ученый
Galmiche A, Fueller J. RAF киназы и митохондрии. Biochim Biophys Acta. 2007. 1773 (8): 1256–62.
CAS PubMed Статья Google ученый
Kerr EM, Gaude E, Turrell FK, Frezza C, Martins CP. Число копий мутанта Kras определяет метаболическое перепрограммирование и терапевтическую восприимчивость.Природа. 2016; 531 (7592): 110–3.
CAS PubMed PubMed Central Статья Google ученый
Grabacka M, Pierzchalska M, Reiss K. Активированные пролифератором пероксисомы рецепторы альфа-лиганды как противораковые препараты, направленные на митохондриальный метаболизм. Curr Pharm Biotechnol. 2013; 14: 342–56.
Eales KL, Hollinshead KE, Tennant DA. Гипоксия и метаболическая адаптация раковых клеток. Онкогенез.2016; 5: e190.
CAS PubMed PubMed Central Статья Google ученый
Xu RH, Pelicano H, Zhou Y, Carew JS, Feng L, Bhalla KN, Keating MJ, Huang P. Ингибирование гликолиза в раковых клетках: новая стратегия преодоления лекарственной устойчивости, связанной с митохондриальным респираторным дефектом и гипоксия. Cancer Res. 2005. 65 (2): 613–21.
CAS PubMed Google ученый
Хенсли, Коннектикут, Вости, АТ, ДеБерардини, Р.Дж. Глютамин и рак: клеточная биология, физиология и клинические возможности. J Clin Invest. 2013. 123 (9): 3678–84.
CAS PubMed PubMed Central Статья Google ученый
Рожок А.И., ДеГрегори Дж. К эволюционной модели рака: рассмотрение механизмов, которые управляют судьбой соматических мутаций. Proc Natl Acad Sci U S. A. 2015; 112 (29): 8914–21.
CAS PubMed PubMed Central Статья Google ученый
Мукерджи П., Малруни Т.Дж., Марш Дж., Блэр Д., Чилийский Т.С., Сейфрид Теннесси. Дифференциальные эффекты энергетического стресса на фосфорилирование AMPK и апоптоз в экспериментальной опухоли головного мозга и нормальном мозге. Молочный рак. 2008; 7: 37.
PubMed PubMed Central Статья CAS Google ученый
Мукерджи П., Сотников А.В., Мангиан Х.Дж., Чжоу Дж.Р., Висек В.Дж., Клинтон С.К. Потребление энергии и рост опухоли простаты, ангиогенез и экспрессия фактора роста эндотелия сосудов.J Natl Cancer Inst. 1999. 91 (6): 512–23.
CAS PubMed Статья Google ученый
Небелинг Л.С., Миральди Ф., Шурин С.Б., Лернер Э. Влияние кетогенной диеты на метаболизм опухоли и нутритивный статус у педиатрических онкологических пациентов: два клинических случая. J Am Coll Nutr. 1995. 14 (2): 202–8.
CAS PubMed Статья Google ученый
Zuccoli G, Marcello N, Pisanello A, Servadei F, Vaccaro S, Mukherjee P, Seyfried TN.Метаболическое управление мультиформной глиобластомой с использованием стандартной терапии вместе с ограниченной кетогенной диетой: отчет о болезни. Нутр Метаб (Лондон). 2010; 7 (1): 33.
Артикул CAS Google ученый
Мукерджи П., Эль-Аббади М.М., Касперзик Дж.Л., Ранес М.К., Сейфрид Т.Н. Ограничение диеты снижает ангиогенез и рост на модели ортотопической опухоли мозга мышей. Br J Рак. 2002. 86 (10): 1615–21.
CAS PubMed PubMed Central Статья Google ученый
Mukherjee P, Abate LE, Сейфрид Теннесси. Антиангиогенные и проапоптотические эффекты ограничения питания на экспериментальные опухоли головного мозга мышей и человека. Clin Cancer Res. 2004. 10 (16): 5622–9.
CAS PubMed Статья Google ученый
Сейфрид Т.Н., Сандерсон Т.М., Эль-Аббади М.М., Макгоуэн Р., Мукерджи П. Роль глюкозы и кетоновых тел в метаболическом контроле экспериментального рака мозга. Br J Рак. 2003. 89 (7): 1375–82.
CAS PubMed PubMed Central Статья Google ученый
Сейфрид Т.Н., Мукерджи П. Антиангиогенные и проапоптотические эффекты ограничения диеты при экспериментальном раке мозга: роль глюкозы и кетоновых тел. В: Медоуз Г.Г., редактор. Интеграция / взаимодействие онкологического роста. Том 15. 2-е изд. Нью-Йорк: Клувер; 2005. с. 259–70.
Глава Google ученый
Чжоу В., Мукерджи П., Кибиш М.А., Маркис В. Т., Мантис Дж. Г., Сейфрид Т. Н.. Кетогенная диета с ограничением калорийности, эффективная альтернативная терапия злокачественного рака мозга. Нутр Метаб (Лондон). 2007; 4: 5.
Артикул CAS Google ученый
Сото AM, Sonnenschein C. Теория соматической мутации рака: растущие проблемы с парадигмой? Биологические исследования. 2004. 26 (10): 1097–107.
CAS PubMed Статья Google ученый
Nowell ПК. Клональная эволюция популяций опухолевых клеток. Наука. 1976 г., 194 (4260): 23–8.
CAS PubMed Статья Google ученый
Кэхилл-младший GF. Топливный обмен при голодании. Анну Рев Нутр. 2006; 26: 1–22.
CAS PubMed Статья Google ученый
Маги Б.А., Потезный Н., Рофе А.М., Коньерс РА. Подавление кетоновыми телами роста злокачественных клеток.Aust J Exp Biol Med Sci. 1979; 57 (5): 529–39.
CAS PubMed Статья Google ученый
Скиннер Р., Трухильо А., Ма Х, Байерле Е.А. Кетоновые тела подавляют жизнеспособность клеток нейробластомы человека. J Pediatr Surg. 2009. 44 (1): 212–6. обсуждение 216.
PubMed Статья Google ученый
Maurer GD, Brucker DP, Baehr O, Harter PN, Hattingen E, Walenta S, Mueller-Klieser W, Steinbach JP, Rieger J.Дифференциальное использование кетоновых тел нейронами и линиями клеток глиомы: обоснование кетогенной диеты в качестве экспериментальной терапии глиомы. BMC Рак. 2011; 11 (1): 315.
CAS PubMed PubMed Central Статья Google ученый
Chang HT, Olson LK, Schwartz KA. Профили экспрессии кетолитических и гликолитических ферментов в злокачественных глиомах: значение для кетогенной диетотерапии. Нутр Метаб. 2013; 10 (1): 47.
CAS Статья Google ученый
Малруни Т.Дж., Марш Дж., Уриц И., Сейфрид Т.Н., Мукерджи П. Влияние ограничения калорий на конститутивную экспрессию NF-kappaB в экспериментальной астроцитоме мыши. PLoS One. 2011; 6 (3): e18085.
CAS PubMed PubMed Central Статья Google ученый
Абдельвахаб М.Г., Фентон К.Э., Преул М.С., Ро Дж. М., Линч А., Стаффорд П., Шек А.С. Кетогенная диета является эффективным дополнением к лучевой терапии для лечения злокачественной глиомы.PLoS One. 2012; 7 (5): e36197.
PubMed PubMed Central Статья Google ученый
Martuscello RT, Vedam-Mai V, McCarthy DJ, Schmoll ME, Jundi MA, Louviere CD, Griffith BG, Skinner CL, Suslov O, Deleyrolle LP, et al. Диета с добавлением жиров и низким содержанием углеводов для лечения глиобластомы. Clin Cancer Res. 2015; 22: 2482–95.
Винсент М. Рак: де-репрессия стандартной программы выживания, общей для всех клеток?: Взгляд на природу рака из истории жизни.BioEssays. 2012. 34 (1): 72–82.
CAS PubMed Статья Google ученый
Сервантес-Мадрид Д., Ромеро И., Дуэнас-Гонсалес А. Возрождение лонидамина и 6-диазо-5-оксо-L-норлейцина для использования в комбинации для метаболической терапии рака. Biomed Res Int. 2015; 2015: 6
.
PubMed PubMed Central Статья CAS Google ученый
Фриман Дж. М., Коссофф Э. Х.Кетоз и кетогенная диета, 2010: достижения в лечении эпилепсии и других расстройств. Adv Pediatr. 2010. 57 (1): 315–29.
PubMed Статья Google ученый
Kossoff EH, Hartman AL. Кетогенные диеты: новые достижения в терапии, основанной на метаболизме. Curr Opin Neurol. 2012; 25 (2): 173.
CAS PubMed PubMed Central Статья Google ученый
Meidenbauer JJ, Mukherjee P, Seyfried TN. Калькулятор кетонового индекса глюкозы: простой инструмент для отслеживания терапевтической эффективности метаболического лечения рака мозга. Нутр Метаб (Лондон). 2015; 12: 12.
Артикул CAS Google ученый
Poff AM, Ward N, Seyfried TN, Arnold P, D’Agostino DP. Нетоксическое метаболическое управление метастатическим раком у мышей VM: новая комбинация кетогенной диеты, добавок кетонов и гипербарической кислородной терапии.PLoS One. 2015; 10 (6): e0127407.
CAS PubMed PubMed Central Статья Google ученый
Берт М.Э., Горшбот С.М., Бреннан М.Ф. Контролируемое проспективное рандомизированное исследование, оценивающее метаболические эффекты энтерального и парентерального питания у онкологического пациента. Рак. 1982. 49 (6): 1092–105.
CAS PubMed Статья Google ученый
Кэмпбелл ТС. Диетический белок, факторы роста и рак. Am J Clin Nutr. 2007; 85 (6): 1667.
CAS PubMed Google ученый
Лу З., Се Дж., Ву Дж., Шен Дж., Коллинз Р., Чен В., Кан Х, Ло М, Цзоу Й, Хуанг Л. Дж. И др. Голодание избирательно блокирует развитие острого лимфобластного лейкоза за счет активации рецепторов лептина. Природа. 2017; 23: 79–90.
Цзян Ю.С., Ван Фр. Ограничение калорийности уменьшает отек и продлевает выживаемость на модели глиомы мышей.J Neuro-Oncol. 2013. 114 (1): 25–32.
Артикул Google ученый
Тисдейл М.Дж., Бреннан, РА. Сравнение длинноцепочечных триглицеридов и среднецепочечных триглицеридов при потере веса и размере опухоли на модели кахексии. Br J Рак. 1988. 58 (5): 580–3.
CAS PubMed PubMed Central Статья Google ученый
Tisdale MJ, Brennan RA, Fearon KC.Уменьшение потери веса и размера опухоли на модели кахексии с помощью диеты с высоким содержанием жиров. Br J Рак. 1987. 56 (1): 39–43.
CAS PubMed PubMed Central Статья Google ученый
Люсье Д.М., Вульф Е.С., Джонсон Дж.Л., Брукс К.С., Блаттман Дж. Н., Шек А.С. Повышенный иммунитет на мышиной модели злокачественной глиомы опосредуется терапевтической кетогенной диетой. BMC Рак. 2016; 16: 310.
PubMed PubMed Central Статья Google ученый
Шукла С.К., Гебрегиворгис Т., Пурохит В., Чайка Н.В., Гунда В., Радхакришнан П., Мехла К., Пипинос II, Пауэрс Р., Ю Ф и др. Перепрограммирование метаболизма, вызванное кетоновыми телами, уменьшает кахексию рака поджелудочной железы. Метаболизм рака. 2014; 2: 18.
PubMed PubMed Central Статья Google ученый
Morscher RJ, Aminzadeh-Gohari S, Feichtinger RG, Mayr JA, Lang R, Neureiter D, Sperl W., Kofler B. Ингибирование роста опухоли нейробластомы с помощью кетогенной диеты и / или ограничение калорийности CD1-Nu Модель мыши.PLoS One. 2015; 10 (6): e0129802.
PubMed PubMed Central Статья CAS Google ученый
Morscher RJ, Aminzadeh-Gohari S, Hauser-Kronberger C, Feichtinger RG, Sperl W., Kofler B. Комбинация метрономного циклофосфамида и диетического вмешательства подавляет рост нейробластомы на мышиной модели CD1-nu. Oncotarget. 2016; 7 (13): 17060–73.
PubMed PubMed Central Google ученый
Аллен Б.Г., Бхатия С.К., Буатти Д.М., Брандт К.Э., Линдхольм К.Э., Баттон А.М., Шведа Л.И., Смит Б.Дж., Шпиц Д.Р., Фатх М.А. Кетогенные диеты усиливают окислительный стресс и реакцию на радиохимиотерапию ксенотрансплантатов рака легких. Clin Cancer Res. 2013. 19 (14): 3905–13.
CAS PubMed PubMed Central Статья Google ученый
Mavropoulos JC, Buschemeyer 3rd WC, Tewari AK, Rokhfeld D, Pollak M, Zhao Y, Febbo PG, Cohen P, Hwang D, Devi G, et al.Влияние изменения содержания углеводов и жиров в рационе на выживаемость в модели ксенотрансплантата рака предстательной железы LNCaP на мышах. Рак Prev Res (Phila). 2009. 2 (6): 557–65.
CAS Статья Google ученый
Ким Х.С., Маско Е.М., Поултон С.Л., Кеннеди К.М., Пиццо С.В., Дьюхерст М.В., Фридленд С.Дж. Ограничение углеводов и ингибирование транспортера лактата в мышиной модели ксенотрансплантата рака простаты человека. BJU Int. 2012. 110 (7): 1062–9.
CAS PubMed PubMed Central Статья Google ученый
Lv M, Zhu X, Wang H, Wang F, Guan W. Роли ограничения калорий, кетогенной диеты и прерывистого голодания во время инициации, прогрессирования и метастазирования рака на животных моделях: систематический обзор и метаанализ . PLoS One. 2014; 9 (12): e115147.
PubMed PubMed Central Статья CAS Google ученый
Zhuang Y, Chan DK, Haugrud AB, Miskimins WK. Механизмы, с помощью которых низкий уровень глюкозы усиливает цитотоксичность метформина для раковых клеток как in vitro, так и in vivo. PLoS One. 2014; 9 (9): e108444.
PubMed PubMed Central Статья CAS Google ученый
Hao GW, Chen YS, He DM, Wang HY, Wu GH, Zhang B. Рост клеток рака толстой кишки человека у голых мышей задерживается кетогенной диетой с жирными кислотами омега-3 или без них и среднецепочечной триглицериды.Азиатский Pac J Cancer Prev. 2015; 16 (5): 2061–8.
PubMed Статья Google ученый
Maroon JC, Сейфрид TN, Donohue JP, Bost J. Роль метаболической терапии в лечении мультиформной глиобластомы. Surg Neurol Int. 2015; 6: 61.
PubMed PubMed Central Статья Google ученый
Ригер Дж., Бахр О., Маурер Г.Д., Хаттинген Э., Франц К., Брукер Д., Валента С., Каммерер Ю., Кой Дж. Ф., Веллер М. и др.ERGO: пилотное исследование кетогенной диеты при рецидивирующей глиобластоме. Int J Oncol. 2014; 44 (6): 1843–52.
CAS PubMed PubMed Central Google ученый
Клемент Р.Дж. Ограничение калорий или углеводов? Кетогенная диета как еще один вариант поддерживающего лечения рака. Онколог. 2013; 18 (9): 1056.
PubMed PubMed Central Статья Google ученый
Klement RJ. Ограничение углеводов для борьбы с раком головы и шеи — это реально? Рак биол мед. 2014. 11 (3): 145–61.
PubMed PubMed Central Google ученый
Тан-Шалаби Дж. Л., Каррик Дж., Эдингер К., Дженовезе Д., Лиман А. Д., Пассеро В. А., Шах РБ. Модифицированная диета Аткинса при запущенных злокачественных новообразованиях — окончательные результаты исследования безопасности и осуществимости в системе здравоохранения Питтсбурга по делам ветеранов. Нутр Метаб (Лондон).2016; 13:52.
Артикул Google ученый
Шмидт М., Пфетцер Н., Шваб М., Штраус И., Каммерер У. Влияние кетогенной диеты на качество жизни 16 пациентов с запущенным раком: пилотное испытание. Нутр Метаб. 2011; 8 (1): 54.
CAS Статья Google ученый
Champ CE, Палмер JD, Volek JS, Werner-Wasik M, Andrews DW, Evans JJ, Glass J, Kim L, Shi W.Нацеливание на метаболизм с помощью кетогенной диеты при лечении мультиформной глиобластомы. J Neuro-Oncol. 2014. 117 (1): 125–31.
CAS Статья Google ученый
Champ CE, Mishra MV, Showalter TN, Ohri N, Dicker AP, Simone NL. Диетические рекомендации во время и после лечения рака: постоянно противоречивы? Nutr Cancer. 2013; 65 (3): 430–9.
CAS PubMed Статья Google ученый
Fine EJ, Сегал-Исааксон CJ, Feinman RD, Herszkopf S, Romano MC, Tomuta N, Bontempo AF, Negassa A, Sparano JA. Нацеленность на ингибирование инсулина в качестве метаболической терапии при запущенном раке: пилотное исследование безопасности и осуществимости диет с участием 10 пациентов. Питание. 2012. 28 (10): 1028–35.
CAS PubMed Статья Google ученый
Шварц К., Чанг Х.Т., Николай М., Перниконе Дж., Ри С., Олсон К., Курниали П.С., Хорд Н.Г., Ноэль М.Лечение пациентов с глиомой с помощью кетогенной диеты: отчет о двух случаях, получавших одобренный IRB протокол кетогенной диеты с ограничением энергии, и обзор литературы. Метаболизм рака. 2015; 3: 3.
PubMed PubMed Central Статья Google ученый
Клемент Р.Дж., Суини Р.А. Влияние кетогенной диеты во время лучевой терапии на состав тела: I. Первоначальный клинический опыт с шестью проспективно изученными пациентами.BMC Res Notes. 2016; 9: 143.
PubMed PubMed Central Статья CAS Google ученый
Фриман Дж. М., Коссофф Э. Х., Фриман Дж. Б., Келли М. Т.. Кетогенная диета: лечение детей и других больных эпилепсией. 4-е изд. Нью-Йорк: Демос; 2007.
Google ученый
Mantis JG, Centeno NA, Todorova MT, McGowan R, Seyfried TN. Управление многофакторной идиопатической эпилепсией у мышей EL с ограничением калорийности и кетогенной диетой: роль глюкозы и кетоновых тел.Нутр Метаб (Лондон). 2004; 1 (1): 11.
Артикул CAS Google ученый
Кэхилл-младший Г.Ф., Вич Р.Л. Кетокислоты? Хорошее лекарство? Trans Am Clin Climatol Assoc. 2003. 114: 149–61. обсуждение 162–143.
PubMed PubMed Central Google ученый
Фейн Э. Дж., Фейнман РД. Инсулин, ограничение углеводов, метаболический синдром и рак. Эксперт Rev Endocrinol Metab.2015; 10: 15–24.
Артикул CAS Google ученый
Сато К., Кашивайя Й., Кеон, Калифорния, Цучия Н., Кинг М.Т., Радда Г.К., Чанс Б., Кларк К., Вич Р.Л. Инсулин, кетоновые тела и митохондриальная трансдукция энергии. Фасеб Дж. 1995; 9 (8): 651–658.
CAS PubMed Google ученый
VanItallie TB, Nufert TH. Кетоны: гадкий утенок метаболизма. Nutr Rev.2003. 61 (10): 327–41.
PubMed Статья Google ученый
Veech RL, Chance B, Kashiwaya Y, Lardy HA, Cahill Jr GF. Кетоновые тела, потенциальное терапевтическое использование. МСБМБ Жизнь. 2001. 51 (4): 241–7.
CAS PubMed Статья Google ученый
Chance B, Sies H, Boveris A. Метаболизм гидропероксида в органах млекопитающих. Physiol Rev.1979; 59 (3): 527–605.
CAS PubMed Google ученый
Fine EJ, Miller A, Quadros EV, Sequeira JM, Feinman RD. Ацетоацетат снижает рост и концентрацию АТФ в линиях раковых клеток, которые чрезмерно экспрессируют разобщающий белок 2. Cancer Cell Int. 2009; 9: 14.
PubMed PubMed Central Статья CAS Google ученый
Ciraolo ST, Previs SF, Fernandez CA, Agarwal KC, David F, Koshy J, Lucas D, Tammaro A, Stevens MP, Tserng KY, et al.Модель экстремальной гипогликемии у собак, приготовленная кетотиком с (R, S) -1,3-бутандиол-ацетоацетатными эфирами. Am J Phys. 1995; 269 (1, часть 1): E67–75.
CAS Google ученый
Шанс Б, редактор. Энергетические функции митохондрий. Нью-Йорк: академический; 1963.
Google ученый
Симадзу Т., Хирши М.Д., Ньюман Дж., Хе В., Сиракава К., Ле Моан Н., Грютер К.А., Лим Х., Сондерс Л.Р., Стивенс Р.Д. и др.Подавление окислительного стресса с помощью бета-гидроксибутирата, эндогенного ингибитора гистондеацетилазы. Наука. 2013. 339 (6116): 211–4.
CAS PubMed Статья Google ученый
West AC, Johnstone RW. Новые и появляющиеся ингибиторы HDAC для лечения рака. J Clin Invest. 2014; 124 (1): 30–9.
CAS PubMed PubMed Central Статья Google ученый
Юм Й., Нгуен К. Ю., Грант Р. В., Голдберг Е. Л., Бодогай М., Ким Д., Д’Агостино Д., Планавский Н., Лупфер С., Каннеганти Т. Д. и др. Кетоновый метаболит бета-гидроксибутират блокирует воспалительное заболевание, опосредованное воспалением NLRP3. Nat Med. 2015; 21 (3): 263–9.
CAS PubMed PubMed Central Google ученый
Kossoff EH, Zupec-Kania BA, Amark PE, Ballaban-Gil KR, Christina Bergqvist AG, Blackford R, Buchhalter JR, Caraballo RH, Helen Cross J, Dahlin MG, et al.Оптимальное клиническое ведение детей, получающих кетогенную диету: рекомендации Международной исследовательской группы по кетогенной диете. Эпилепсия. 2009. 50 (2): 304–17.
PubMed Статья Google ученый
Чан ХД, Бу ХД, Ли ХД, Мин ХЙ, Ли ХЙ. Хронический стресс способствует онкогенезу легких, способствуя экзоцитозу IGF2 в эпителиальных клетках легких. Cancer Res. 2016; 76 (22): 6607–19.
CAS PubMed Статья Google ученый
Feng Z, Liu L, Zhang C, Zheng T, Wang J, Lin M, Zhao Y, Wang X, Levine AJ, Hu W. Хронический стресс сдерживания ослабляет функцию p53 и способствует онкогенезу. Proc Natl Acad Sci U S. A. 2012; 109 (18): 7013–8.
CAS PubMed PubMed Central Статья Google ученый
Раш С.Е., Шарма М. Снижение стресса на основе осознанности как вмешательство по управлению стрессом для лечения рака: систематический обзор. J Evid Based Complementary Altern Med.2014; 19: 271–86.
Lopes-Junior LC, Bomfim EO, Nascimento LC, Nunes MD, Pereira-da-Silva G, Lima RA. Немедикаментозные вмешательства для управления усталостью и психологическим стрессом у детей и подростков с онкологическими заболеваниями: интегративный обзор. Eur J Cancer Care (англ.). 2016; 25 (6): 921–35.
CAS Статья Google ученый
Брэдт Дж., Дилео К., Мэджилл Л., Тиг А. Музыкальные вмешательства для улучшения психологических и физических результатов у онкологических больных.Кокрановская база данных Syst Rev.2016; 8: CD006911.
Google ученый
Левин Г.Т., Гринвуд К.М., Сингх Ф., Цой Д., Ньютон РУ. Упражнения улучшают физическую функцию и психическое здоровье выживших после рака мозга: два исследовательских примера. Integr Cancer Ther. 2016; 15 (2): 190–6.
PubMed Статья Google ученый
Ари С., Ковач З., Юхас Дж., Мурдун С., Голдхаген С.Р., Кутник А.М., Пофф А.М., Кесл С.Л., Д’Агостино Д.П.Добавки с экзогенными кетонами уменьшают поведение, связанное с тревогой, у крыс Sprague – Dawley и Wistar Albino Glaxo / Rijswijk. Front Mol Neurosci. 2016; 9: 137.
PubMed PubMed Central Статья Google ученый
Meynet O, Ricci JE. Ограничение калорийности и рак: молекулярные механизмы и клинические последствия. Тенденции Мол Мед. 2014. 20 (8): 419–27.
CAS PubMed Статья Google ученый
De Lorenzo MS, Baljinnyam E, Vatner DE, Abarzua P, Vatner SF, Rabson AB. Ограничение калорийности снижает рост опухолей молочной железы и метастазов. Канцерогенез. 2011; 32 (9): 1381–7.
PubMed PubMed Central Статья CAS Google ученый
Лонго В.Д., Маттсон М.П. Голодание: молекулярные механизмы и клиническое применение. Cell Metab. 2014; 19 (2): 181–92.
CAS PubMed PubMed Central Статья Google ученый
Аль-Вахаб З., Теббе С., Чхина Дж., Дар С.А., Моррис Р.Т., Али-Фехми Р., Гири С., Мункара А.Р., Раттан Р. Энергетический баланс пищи модулирует прогрессирование и метастазирование рака яичников. Oncotarget. 2014; 5 (15): 6063–75.
PubMed PubMed Central Статья Google ученый
Сафди Ф.М., Дорфф Т., Куинн Д., Фонтана Л., Вэй М., Ли С., Коэн П., Лонго В.Д. Голодание и лечение рака у людей: отчет о серии случаев. Старение (Олбани, штат Нью-Йорк).2009; 1 (12): 988–1007.
Артикул Google ученый
Раффагелло Л., Ли С., Сафди FM, Вей М., Мадиа Ф., Бьянки Дж., Лонго В.Д. Дифференциальная устойчивость к стрессу, зависящая от голода, защищает нормальные, но не раковые клетки от высокодозной химиотерапии. Proc Natl Acad Sci U S. A. 2008; 105 (24): 8215–20.
CAS PubMed PubMed Central Статья Google ученый
Раффагелло Л., Сафди Ф, Бьянки Дж., Дорфф Т., Фонтана Л., Лонго В.Д. Голодание и дифференциальная химиотерапевтическая защита у пациентов. Клеточный цикл. 2010. 9 (22): 4474–6.
CAS PubMed PubMed Central Статья Google ученый
Marsh J, Mukherjee P, Seyfried TN. Синергия лекарств и диеты для лечения злокачественной астроцитомы у мышей: 2-дезокси-D-глюкоза и ограниченная кетогенная диета. Нутр Метаб (Лондон). 2008; 5: 33.
Артикул CAS Google ученый
Williams DS, Cash A, Hamadani L, Diemer T. Добавление оксалоацетата увеличивает продолжительность жизни Caenorhabditis elegans через AMPK / FOXO-зависимый путь. Ячейка старения. 2009. 8 (6): 765–8.
CAS PubMed PubMed Central Статья Google ученый
Фарах И.О. Дифференциальная модуляция внутриклеточной энергетики в клетках A549 и MRC-5. Biomed Sci Instrum. 2007; 43: 110–5.
CAS PubMed Google ученый
Pelicano H, Martin DS, Xu RH, Huang P. Ингибирование гликолиза для противоопухолевого лечения. Онкоген. 2006. 25 (34): 4633–46.
CAS PubMed Статья Google ученый
Питтер К.Л., Тамагно И., Алиханян К., Хосни-Ахмед А., Паттуэлл С.С., Доннола С., Дай К., Озава Т., Чанг М., Чан Т.А. и др. Кортикостероиды снижают выживаемость при глиобластоме. Головной мозг. 2016; 139 (Pt 5): 1458–71.
PubMed Статья Google ученый
Сейфрид Т.Н., Флорес Р., Пофф А.М., Д’Агостино Д.П., Мукерджи П. Метаболическая терапия: новая парадигма лечения злокачественного рака мозга. Cancer Lett. 2015; 356 (2 Pt A): 289–300.
CAS PubMed Статья Google ученый
Сейфрид Т.Н., Шелтон Л.М., Мукерджи П. Повышает ли существующий стандарт лечения энергетический метаболизм глиобластомы? Ланцет Онкол. 2010. 11 (9): 811–3.
PubMed Статья Google ученый
Moen I, Stuhr LE. Гипербарическая кислородная терапия и рак — обзор. Target Oncol. 2012. 7 (4): 233–42.
PubMed PubMed Central Статья Google ученый
Кохши К., Беппу Т., Танака К., Огава К., Иноуэ О, Кукита И., Кларк РЭ. Возможные роли гипербарической оксигенации в лечении опухолей головного мозга. Ухм. 2013. 40 (4): 351–62.
Google ученый
Poff AM, Kernagis D, D’Agostino DP. Гипербарическая среда: кислород и повреждение клеток против защиты. Комп Физиология. 2017; 7 (январь 2017): 213–34.
Google ученый
D’Agostino DP, Colomb Jr DG, Dean JB. Влияние гипербарических газов на мембранную наноструктуру и функцию нейронов. J Appl Physiol. 2009; 106 (3): 996–1003.
PubMed Статья Google ученый
Ma Y, Chapman J, Levine M, Polireddy K, Drisko J, Chen Q. Высокие дозы парентерального аскорбата повышают химиочувствительность рака яичников и снижают токсичность химиотерапии. Sci Transl Med. 2014; 6 (222): 222ra218.
Артикул CAS Google ученый
Мичелакис Э.Д., Сутендра Дж., Дромпарис П., Вебстер Л., Хароми А., Нивен Э., Магуайр С., Гаммер Т.Л., Макки Дж. Р., Фултон Д. и др. Метаболическая модуляция глиобластомы дихлорацетатом.Sci Transl Med. 2010; 2 (31): 31ра34.
CAS PubMed Статья Google ученый
Кокс П.Дж., Кирк Т., Эшмор Т., Уиллертон К., Эванс Р., Смит А., Мюррей А.Дж., Стаббс Б., Вест Дж., МакЛюр С.В. и др. Пищевой кетоз влияет на предпочтение топлива и, следовательно, на выносливость у спортсменов. Cell Metab. 2016; 24 (2): 256–68.
CAS PubMed Статья Google ученый
Мюррей AJ, Knight NS, Cole MA, Cochlin LE, Carter E, Tchabanenko K, Pichulik T, Gulston MK, Atherton HJ, Schroeder MA и др. Новая кетоновая диета улучшает физические и когнитивные способности. FASEB J. 2016; 30 (12): 4021–32.
CAS PubMed PubMed Central Статья Google ученый
Wise DR, DeBerardinis RJ, Mancuso A, Sayed N, Zhang XY, Pfeiffer HK, Nissim I., Daikhin E, Yudkoff M, McMahon SB, et al. Myc регулирует программу транскрипции, которая стимулирует митохондриальный глутаминолиз и приводит к зависимости от глутамина.Proc Natl Acad Sci U S. A. 2008; 105 (48): 18782–7.
CAS PubMed PubMed Central Статья Google ученый
Reitzer LJ, Wice BM, Kennell D. Доказательства того, что глутамин, а не сахар, является основным источником энергии для культивируемых клеток HeLa. J Biol Chem. 1979; 254 (8): 2669–76.
CAS PubMed Google ученый
Dang CV. Глутаминолиз: снабжение раковых клеток углеродом, азотом или и тем, и другим? Клеточный цикл.2010. 9 (19): 3884–6.
CAS PubMed Статья Google ученый
Venneti S, Dunphy MP, Zhang H, Pitter KL, Zanzonico P, Campos C, Carlin SD, La Rocca G, Lyashchenko S, Ploessl K, et al. ПЭТ-визуализация на основе глутамина способствует усиленной метаболической оценке глиом in vivo. Sci Transl Med. 2015; 7 (274): 274ra217.
Артикул CAS Google ученый
Mueller C, Al-Batran S, Jaeger E, Schmidt B, Bausch M, Unger C, Sethuraman N. Исследование фазы IIa ПЭГ-илированной глутаминазы (PEG-PGA) плюс 6-диазо-5-оксо-L-норлейцин (DON ) у пациентов с далеко зашедшими рефрактерными солидными опухолями. J Clin Oncol. 2008; 26: 2533. В: ASCO.
CAS Google ученый
Чакрабарти Г., Мур З. Р., Ло Х, Ильчева М., Али А., Паданад М., Чжоу И., Се Ю., Бирма С., Скаглиони П. П. и др. Нацеленность на метаболизм глутамина повышает чувствительность рака поджелудочной железы к метаболической катастрофе, вызванной PARP, вызванной β-лапахоном.Рак и метаболизм. 2015; 3: 12.
Артикул Google ученый
Матес Дж. М., Сегура Дж. А., Кампос-Сандовал Дж. А., Лобо С., Алонсо Л., Алонсо Ф. Дж., Маркес Дж. Гомеостаз глутамина и митохондриальная динамика. Int J Biochem Cell Biol. 2009. 41 (10): 2051–61.
CAS PubMed Статья Google ученый
Михалак КП, Мацковска-Кедзиора А, Соболевский Б., Возняк П.Ключевые роли глутаминовых путей в перепрограммировании метаболизма рака. Oxid Med Cell Longev. 2015; 2015: 964321.
PubMed PubMed Central Статья Google ученый
Huysentruyt LC, Mukherjee P, Banerjee D, Shelton LM, Seyfried TN. Метастатические раковые клетки со свойствами макрофагов: данные новой модели опухоли мышей. Int J Cancer. 2008. 123 (1): 73–84.
CAS PubMed Статья Google ученый
Shelton LM, Mukherjee P, Huysentruyt LC, Urits I, Rosenberg JA, Seyfried TN. Новая доклиническая модель in vivo на мышах для роста и инвазии злокачественных опухолей головного мозга. J Neurooncol. 2010. 99 (2): 165–76.
CAS PubMed Статья Google ученый
Huysentruyt LC, Shelton LM, Сейфрид Теннесси. Влияние метотрексата и цисплатина на прогрессирование опухоли и выживаемость в модели системного метастатического рака у мышей VM.Int J Cancer. 2010. 126 (1): 65–72.
CAS PubMed Статья Google ученый
Гамильтон Дж. Д., Рапп М., Шнейдерхан Т., Сабель М., Хейман А., Шерер А., Кропил П., Будах В., Гербер П., Кречмар Ю. и др. Мультиформные метастазы глиобластомы вне ЦНС: три истории болезни и возможные механизмы побега. J Clin Oncol. 2014; 32 (22): e80–84.
PubMed Статья Google ученый
Hoffman HJ, Даффнер ПК. Экстраневральные метастазы опухолей центральной нервной системы. Рак. 1985; 56 (7 доп.): 1778–82.
CAS PubMed Статья Google ученый
Xu M, Wang Y, Xu J, Yao Y, Yu WX, Zhong P. Расширенные методы лечения экстраневральных метастазов глиобластомы, подтвержденные с помощью анализа OncoScan. World Neurosurg. 2016; 90: 698 e697–11.
Артикул Google ученый
Ясухара Т., Тамия Т., Мегуро Т., Итикава Т., Сато Й., Дате I, Накашима Н., Омото Т. Глиобластома с метастазами в селезенку — отчет о клиническом случае. Neurol Med Chir (Токио). 2003. 43 (9): 452–6.
Артикул Google ученый
Kalokhe G, Grimm SA, Chandler JP, Helenowski I., Rademaker A, Raizer JJ. Метастатическая глиобластома: презентации случаев и обзор литературы. J Neurooncol. 2012; 107 (1): 21–7.
PubMed Статья Google ученый
Huysentruyt LC, Akgoc Z, Сейфрид TN. Гипотеза: присутствуют ли неопластические макрофаги / микроглия в мультиформной глиобластоме? АСН нейро. 2011; 3 (4): AN20110011.
Артикул Google ученый
Ньюсхолм П. Почему метаболизм L-глутамина важен для клеток иммунной системы в состоянии здоровья, после травм, хирургических вмешательств или инфекций? J Nutr. 2001; 131 (9 доп.): 2515–2522S. обсуждение 2523С-2514С.
Google ученый
Shelton LM, Huysentruyt LC, Mukherjee P, Seyfried TN. Ограничение калорийности как противоинвазивная терапия злокачественного рака мозга у мышей VM. АСН нейро. 2010; 2 (3): e00038.
PubMed PubMed Central Статья CAS Google ученый
Сейфрид TN. Метаболическое лечение рака. В: Рак как заболевание обмена веществ: происхождение, лечение и профилактика рака. edn. Хобокен: Уайли; 2012. с. 291–354.
Глава Google ученый
Арисменди-Морилло Г. Электронная микроскопия морфологии митохондриальной сети при раке человека. Int J Biochem Cell Biol. 2009. 41 (10): 2062–8.
CAS PubMed Статья Google ученый
Cogliati S, Frezza C, Soriano ME, Varanita T, Quintana-Cabrera R, Corrado M, Cipolat S, Costa V, Casarin A, Gomes LC, et al.Форма крист митохондрий определяет сборку суперкомплексов дыхательной цепи и эффективность дыхания. Клетка. 2013; 155 (1): 160–71.
CAS PubMed PubMed Central Статья Google ученый
О выставке | МЕТАБОЛИЗМ: ГОРОД БУДУЩЕГО — Мечты и видения реконструкции в Японии прошлого и настоящего
Первая ретроспектива метаболизма, движения, рожденного видением архитекторов
В 1960-е годы группа японских архитекторов мечтала о городах будущего и выдвинула новые захватывающие идеи.Видения Курокавы Кишо, Кикутакэ Киёнори, Маки Фумихико и других архитекторов, попавших под влияние Танге Кэндзо, породили архитектурное движение, получившее название «метаболизм». Название, взятое из биологической концепции, произошло от образа архитектуры и городов, которые разделяли способность живых организмов продолжать расти, воспроизводиться и трансформироваться в ответ на окружающую среду. Их идеи были великолепны и удивительны, с такими концепциями, как морские города, охватывающие Токийский залив, и города, соединенные шоссе в небе, где автомобили проезжают между группами высотных зданий.
Обмен веществ возник в то время, когда Япония оправилась от разрушительной войны и вступила в период быстрого экономического роста. Люди чувствовали, что создание идеальных городов — это способ построить лучшие сообщества. Эта выставка — первая выставка, которая всесторонне исследует метаболизм. Сейчас Японии предстоит принять важные решения относительно своего будущего. Это идеальное время, чтобы узнать о движении «Метаболизм» и открыть для себя некоторые из его многочисленных намеков на архитектуру и города.
Выставка состоит из четырех разделов, плюс Metabolism Lounge.
Раздел 1: Рождение метаболизма
Метаболизм — архитектурное движение, зародившееся в Японии, но оказавшее влияние на весь мир. Танге Кензо, который думал об архитектуре в масштабе городов, оказал особенно сильное влияние на зарождение метаболизма. В этом разделе исследуется план реконструкции Танге? Генеральный план Хиросимы и Хиросима Мирный центр Комплекс ? Который является четкой отправной точкой для послевоенной японской архитектуры и городского дизайна. Он также прослеживает изменения в японском городском дизайне военного и послевоенного времени, уделяя особое внимание идеям городов будущего, изложенным в 1960 году в манифесте «МЕТАБОЛИЗМ / 1960 — Предложения для нового урбанизма».
Раздел 2: Эра метаболизма
Идеи метаболизма выросли из нереализованных планов футуристических городов, а некоторые приняли форму экспериментальной архитектуры. В этом разделе рассматривается широкий спектр мероприятий, от концептуальных планов города до общественной архитектуры и жилищного строительства, в том числе A Plan for Tokyo? 1960 , в котором излагаются новаторские идеи Танге Кензо и его коллег по морскому городу, охватывающему Токийский залив, Nakagin. Здание Capsule Tower Building , которое стало символом метаболизма, демонстрируя городское жилье в виде съемных капсул, и Syowa Station , антарктическая база, которую можно было собрать на месте без специальных навыков с использованием заводских компонентов, предшественника сегодняшнего дня. сборный корпус.
Раздел 3: Из космоса в окружающую среду
Помимо архитектуры и городского планирования, компания Metabolism глубоко вошла в дизайн и искусство через ключевое слово «окружающая среда». Одним из результатов этого участия стала выставка «От космоса к окружающей среде» в 1966 году. Другой пример — выставка Osaka Expo ’70, которая объединила множество разных жанров в мероприятиях на тему «окружающая среда». В этом разделе представлены работы художника Ямагути Кацухиро, графического дизайнера Авазу Киёси и других участников выставки «Из космоса в окружающую среду.«Он также знакомит с выставкой Expo ’70 с точки зрения городского планирования и архитектуры.
Раздел 4: Глобальный метаболизм
Примерно со времени выставки Expo ’70 Tange Kenzo и другие архитекторы Metabolism начали работать на международном уровне. С тех пор они создали проекты по всему миру, в которых идеи метаболизма претворяются в жизнь во многих различных формах. Здесь мы исследуем основные проекты в масштабах города, в том числе Генеральный план Танге Кензо по реконструкции центра города Скопье в Македонии и Республиканский политехнический институт Маки Фумихико .Включая нереализованные проекты и проекты, которые все еще находятся в стадии реализации, мы пытаемся проверить, в какой степени метаболизм способствовал росту городов по всему миру, и особенно в Азии.
Metabolism Lounge — актуальные идеи о моделях городов будущего
Принимая различные формы, мышление и идеи метаболизма все еще живы в архитектуре и городском дизайне сегодня. Зал Metabolism Lounge включает экспонаты, демонстрирующие подразделения по оказанию помощи при стихийных бедствиях Экуана Кенджи (временные конструкции QS72, разработанные GK Sekkei), которые использовались в больницах после землетрясения в Восточной Японии, а также исследования проекта GREEN FLOAT корпорации Shimizu, который унаследовал многие из его подходов от Metabolism .В Lounge также есть много книг, связанных с метаболизмом и городским дизайном, и будут представлены видеоролики о симпозиумах, проводимых в период выставок, что делает его идеальным местом для размышлений о будущем архитектуры и городов, созданных идеями метаболизма.
* В связи с мероприятиями салон Metabolism Lounge не будет работать в субботу, 17 сентября /
чт, 6 октября / пн, 17 октября / чт, 27 октября — вт, 8 ноября / сб, 19 ноября / пт, 25 ноября / Чт, 1 декабря / вт, 6 декабря / сб, 10 декабря / чт, 15 декабря
Модель реального мира для визуализации и совместного создания здоровых городов — Природа городов
Благодаря картам и диаграммам, созданным гражданами, основанным на реальных условиях и построенным вокруг целостной структуры, возникающие закономерности позволяют жителям и планировщикам задавать вопросы, которые могут привести как к местным, так и к региональным экологическим улучшениям.
Подобно человеческому телу, города — это живые, постоянно развивающиеся организмы. Точно так же, как диета, упражнения, сон или смех могут рассматриваться как индикаторы нашего личного физического и эмоционального благополучия, способы, которыми товары, вода, пассажиры или еда перемещаются в городской экосистеме, определяют здоровье и устойчивость города в более крупном регионе. и глобальные природные системы. Чем больше у нас знаний о том, какие ресурсы поступают в нашу систему, как эти ресурсы используются и что происходит с любым выходом, который организму не нужно поддерживать, тем больше у нас шансов вести сбалансированный и здоровый образ жизни.Хотя разобраться в тонкостях метаболизма нашего собственного тела — непростая задача, целостная оценка чего-то столь же сложного, как современный промышленный город со всеми его физическими и культурными микрокосмами, может показаться сложной задачей. Однако, если мы посмотрим на города через призму метаболизма, как и на наши тела, в центре внимания окажется структура, с помощью которой можно успешно моделировать потоки городских систем. Городской метаболизм, используемый для анализа функционирования городских территорий с точки зрения использования ресурсов и базовой инфраструктуры, помогает нам понять взаимосвязь между деятельностью человека и окружающей средой.
Прикладной городской метаболизм
Городской метаболизм как концепция не нова. Еще в 19 веке Карл Маркс и Фридрих Энгельс признали, что человеческая деятельность изменяет биофизические процессы, анализируя динамические внутренние отношения между людьми и природой. Только спустя более ста лет впервые была формально разработана более целостная оценка анатомии города. В статье 2007 года, озаглавленной «Изменяющийся метаболизм в городах», Кристофер Кеннеди и группа инженеров-строителей из Университета Торонто определили городской метаболизм как «совокупность технических и социально-экономических процессов, происходящих в городах и приводящих к росту. , производство энергии и ликвидация отходов ».
Городской метаболизм формируется и протекает. Графика: Ecocity Builders С тех пор анализ городского метаболизма превратился из академического упражнения в практическое применение. Например, городские метаболические информационные системы (UMIS), прикладная методология, впервые разработанная канадским исследователем, автором и специалистом по системному дизайну доктором Себастьяном Моффаттом, представляет собой стандартизированную структуру «от источника к раковине» для лучшего понимания и анализа городских систем в процессе их обработки. через застроенную среду во времени и пространстве.Ключом к такому отслеживанию и визуализации материального потока, составляющего городской метаболизм, являются диаграммы MetaFlow, также известные как диаграммы Санки. В рамках проекта Eco2 Cities 2010: Экологические города как экономические города Моффатт начал разработку диаграмм для целых городских систем. Стремясь предложить общую перспективу, исследовательская группа провела тематические исследования в городах по всей Азии, которые помогли бы продемонстрировать текущий поток в городах и предложить понимание того, как эти потоки могут быть лучше замкнуты, чтобы избежать такого большого количества отходов и утечек.Диаграмма MetaFlow энергетической системы Цзиньцзы, Шанхай, показывает несоответствие между текущей системой (слева) и сценарием для усовершенствованной системы (справа). Источник: разработка автора (Себастьян Моффатт) с приблизительными данными, предоставленными профессором Цзиньшенг Ли, Университет Тонги, Шанхай. Не так давно доктор Филип Мэнсфилд из Graphical Memes создал ряд диаграмм MetaFlow для города Ванкувер, Британская Колумбия, на основе данных, предоставленных Директор BCIT по устойчивому развитию, доктор Дженни Мур.Энергетическая диаграмма, например, показывает очень типичную современную централизованную систему с небольшими объемами электроэнергии из местных источников, небольшим разнообразием энергии и минимальным каскадированием, что означает очень низкую эффективность, рециркуляцию или двойное использование. Большая часть энергии Ванкувера в основном поступает из невозобновляемых источников (кроме гидроэнергетики) и попадает в воздух после того, как использовалась для одной цели одновременно. Ванкувер, Британская Колумбия Диаграммы MetaFlow для энергии (слева) и еды (справа). Предоставлено: д-р Филип Мэнсфилд / Graphical Memes Диаграмма MetaFlow Ванкувера для пищевых продуктов показывает намного больше каскадов вверху, указывая на более разнообразные типы продуктов питания, чем типы энергии, с местными фермами, поставляющими видимую часть различных продуктов.Фрукты и овощи составляют значительную часть общего потока органических материалов. Напротив, раковины в нижней части диаграммы имеют гораздо менее текстурированную структуру, при этом большая часть пищевых отходов Ванкувера (которые обычно составляют около 50% всего потока отходов) проходят через перегрузочные станции и мусоросжигательные заводы перед тем, как отправиться на свалку или попасть в мусорную корзину. воздух. Можно рассмотреть множество стратегий для зацикливания и каскадирования этих потоков, то есть для создания более связанной пищевой сети в городе.Например, если бы пищевые отходы компостировались в качестве почвы, почву можно было бы использовать на месте для сельского хозяйства или озеленения, чтобы у города было меньше потребности в транспортировке материалов грузовиками и приобретении земли для свалки.По словам доктора Моффатта, эти диаграммы лучше тысячи круговых диаграмм. Но что, если бы мы могли еще глубже изучить метаболизм города, глядя на потоки ресурсов через каждого района ? Тот факт, что в настоящее время блок-схемы для большинства городов мира будут похожи на Ванкуверские — линейные и централизованные, оставляя мало места для локализованных и адаптируемых способов более эффективного использования как природных, так и человеческих ресурсов — показывает, что существует реальная потребность в инструментах, которые позволяют общинам лучше понимать свои районы и определять области, в которых можно было бы применять больше циклов и каскадов по мере того, как системы становятся более экологичными.
Партнерский городской метаболизм
Введите методологию, разработанную, чтобы дать гражданам возможность планировать свои собственные районы и стать участниками преобразования своих сообществ в более устойчивые, справедливые и экологически здоровые поселения: Совместное участие Городской метаболизм. В этом подходе повышенное внимание уделяется переходу от нисходящего к восходящему подходу к учету городской окружающей среды с целью сбора данных, которые недоступны в обычных базах данных, при одновременном продвижении трансдисциплинарного подхода, при котором осуществляется совместное проектирование с общества, а не для общества , и гарантировать, что оценка не является разовым мероприятием.
Впервые апробированная в рамках инициативы Urbinsight Global Data Initiative Ecocity Builders в Каире и Касабланке, модель недавно была успешно внедрена в Куско, Перу, и Медельине, Колумбия. Эти две демонстрации, созданные по заказу Управления географических городов США (2C) и представленные в блогах и сериалах видео в рамках Глобальной инициативы ООН по окружающей среде для ресурсоэффективных городов, предлагают реальные примеры того, как инструменты и методы городского метаболизма могут использоваться менеджерами и практиками для перехода своих городов к большей устойчивости и ресурсоэффективности.То есть, основываясь на результатах совместных оценок городского метаболизма Куско и Медельина, городские власти, местные университеты и общественные группы в этих городах теперь работают вместе над разработкой Планов устойчивого развития соседства.
Куско: совместный подход к городскому метаболизму
Сосредоточено на столичном районе в городе Куско и его окрестностях на юге Перу, местные жители, городские власти и студенты-исследователи из Universidad Alas Peruanas участвовали в многогранном и многоэтапном подходе к участию в городских условиях. экологический учет с 2016 года.Исторические кварталы центральной части города, где расположено несколько небольших исследовательских зон, все чаще ощущают потребность в такого рода подробном учете условий на местах, чтобы найти и реализовать целостное решение их проблемы с мусором.
Участники, состоящие из преподавателей, студентов, местных должностных лиц, архитекторов и проектировщиков, были впервые ознакомлены с техническими процессами, взаимодействием с общественностью и планированием, лежащими в основе метода совместного городского метаболизма, во время семинаров EcoCompass, проводимых Ecocity Builders на местах команда внедрения.После изучения тонкостей создания платформы динамического картографирования, которая визуализирует несколько типов данных, они отправились в ознакомительные поездки, прежде чем провести круглый стол с лидерами сообщества, которые изложили изменения, которые они хотели видеть в своих районах: более эффективное управление отходами, более чистая окружающая среда и более здоровая пища.
Исходя из этих потребностей, студенческие отряды продолжили проводить аудит материалов в районе. Они собирали данные о потреблении и отходах от жителей, которые участвовали в сортировке твердых отходов, взвешивании материалов по типам и анализе композитов.Затем команда создала подробные виды архетипов кварталов исторических районов, что дает важную информацию о решениях для районов со схожими характеристиками и проблемами. Поскольку почти 50% бытовых отходов оказались органическими, команда решила исследовать методы создания модулей домашнего компостирования, которые они в конечном итоге совместно разработали с помощью членов сообщества и опробовали в четырех сообществах в домах участвующих соседей.
Диаграммы метаболизма в районе, которые были созданы для визуализации собранных данных, стали предметами обсуждения, которые помогают сообществу, а также городским планировщикам принимать обоснованные решения о том, как перенаправить потоки с линейных на круговые.Члены сообщества обнаружили, что для понимания городского метаболизма необходимо понимать происхождение потоков ресурсов, их распределение в пределах города и способы использования ресурсов. Эти идеи позволяют людям создавать осознанные изменения в их уникальном городском контексте, что в настоящее время закреплено в ДНК города как часть следующего этапа проекта: создания Планов устойчивого развития района.
Медельин: Совместное создание более надежных систем городских знаний
В феврале 2013 года Urban Land Institute выбрал Медельин как самый инновационный город в мире из-за его достижений в политике, образовании и социальном развитии.Хотя Медельин был назван самым инновационным городом и является примером городского планирования для всего мира, город по-прежнему сталкивается с серьезным экономическим неравенством. Богатство в основном концентрируется вокруг центра города и экспоненциально убывает на окружающие холмы.
На этапе I проекта Ecocity Builders / 2C Medellín местные планировщики, коммунальные предприятия, ученые и местные некоммерческие организации объединились с членами сообщества Comuna 8, одного из таких уязвимых районов на склоне холма за пределами центра города, состоящего из нескольких малообеспеченных формальных территорий. и неформальные кварталы, чтобы применить методы местного участия к городскому метаболизму.Команда сосредоточила свои данные на сборе и картировании на управлении отходами, материальных потоках в домашние хозяйства и через них, а также на взглядах граждан на методы обращения с отходами в их сообществах в нескольких кварталах в Comuna 8, приоритет, выраженный лидерами сообществ.
После сбора данных участники смогли применить свои тренинги, полученные на семинарах, с помощью приложения Urbinsight metaflow, чтобы превратить вводимые ими коллективные данные в блок-схемы городской метаболической информационной системы.Эти визуализации оказались важны не только для исследователей в плане оптимизации и интерпретации данных о масштабах домохозяйств и участков, но и для жителей Комуны 8, чтобы понять и улучшить свой собственный поток отходов, а для города — в понимании потребностей его жителей.
Диаграммы метапотоков воды, материалов и энергии для Медельина, любезно предоставленные Ecocity Builders. Для фазы II проекта команда определила, что следующим приоритетным типом района должен быть район центра города со смешанным доходом. Многие кварталы этого типа расположены в центре города среди более богатых деловых районов, но муниципалитет часто не замечает их.В результате члены сообществ в этих районах часто имеют более низкий заработок, высокий уровень безработицы, меньшее количество услуг по сбору мусора, меньшее количество услуг в области безопасности или здравоохранения и повышенное загрязнение воздуха.По рекомендации отдела планирования Universidad EAFIT взял на себя ведущую роль в качестве основного академического партнера, который предложил Бостон, район, охарактеризованный как предполагаемый архетип (центр города со смешанным доходом), в качестве рекомендуемой области исследования. Low Carbon City, всемирно признанная некоммерческая организация из Медельина, имеющая прочные связи с бостонским сообществом, присоединилась в качестве местного партнера.
После сбора данных участники разделились на исследовательские группы на основе категорий материалов, которые оказались наиболее распространенными в потоке отходов: пластик, стекло, металл, бумага, органические, опасные отходы и масла, а также смешанные отходы. Используя полевые данные, они подготовили отчеты и презентации о действиях, которые граждане могут предпринять для снижения спроса, повышения эффективности, а также повторного использования / перепрофилирования и переработки материалов дома и в окрестностях, чтобы уменьшить количество отходов в общественных местах.
Во время двух «мапатонов» участники завершили анализ своих аудитов отходов и материалов и подвергли геообработке исходные данные для создания геопространственных слоев для дальнейшего анализа и разработки карт. Затем они работали с командой, чтобы интегрировать слои муниципальных данных с исходными слоями данных о районе, чтобы создать оригинальные карты, которые они представили членам сообщества. На заключительном общественном мероприятии каждая группа представила интерактивные образовательные мероприятия, связанные с их типом тематики ресурсов.Они разработали плакаты и короткие видеоролики, чтобы «рассказать историю» городского метаболизма и роли горожан в ресурсных циклах.
Образец материалов, подготовленных партнером сообщества Low Carbon City для местных семинаров. С момента их сотрудничества в 2017 году участники курса и Low Carbon City поддерживали прочную связь с офисом планирования, который анализировал все данные курса, чтобы быть официально признанным муниципалитет и включены в их портал открытых данных для планирования и анализа.Заинтригованные потенциальным использованием методологии оценки городского метаболизма, специалисты отдела планирования попросили вместе с командой Ecocity Builders / 2C провести общегородской сбор данных для завершения всех доступных исследований городского метаболизма.Сейчас проект вступил в следующую фазу, во время которой партнеры снова объединяются для создания Плана устойчивого соседства. План, основанный на начальных и текущих оценках метаболизма в городах, методах и совместных процессах, предназначен для адаптации и воспроизведения и основан на восходящем подходе Ecocity Builders к развитию экогорода, что подчеркивается оценками городских метаболических информационных систем (UMIS) по соседству. шкала.
Куда мы хотим пойти и почему?
Более устойчивое и устойчивое будущее городов может быть достигнуто за счет использования методов и подходов городского метаболизма в непрерывном и повторяющемся процессе. Здоровый городской обмен веществ, как и здоровая экосистема или организм, работает лучше всего, когда его часто контролируют и постоянно настраивают.
Кроме того, поскольку именно люди определяют спрос на ресурсы, которые формируют метаболизм города, крайне важно, чтобы гражданам было разрешено взвешивать и участвовать в значимой форме.Получение такой информации из первых рук неоценимо не только из-за ранее неизвестных точек данных, которые она предоставляет, но и потому, что личное участие создает новую осведомленность и побуждает сообщество заниматься поиском решений существующих проблем.
Благодаря картам и диаграммам, созданным гражданами, основанным на реальных условиях и структурированным вокруг целостной структуры, возникающие закономерности позволяют жителям и планировщикам задавать вопросы, которые могут привести как к местным, так и к региональным экологическим улучшениям.Что мы можем сделать, чтобы люди могли обходиться дождевой водой? Как городу избежать утечки воды и энергии? Как мы создаем материальные петли, которые решают как проблемы с местными отходами, так и сокращают глобальные выбросы углерода? Как нам оставаться в пределах несущей способности Земли?
Или, как замечает Себастьян Моффат, «мы можем использовать эти диаграммы, чтобы рассказать историю того, куда мы хотим пойти и почему!»
Свен Эберлейн
Окленд
О природе городов
Header Image : Энергетический след транспорта UMIS в Ванкувере.
О писателе:
Свен Эберлейн
Свен — журналист по решениям из Сан-Франциско и системный мыслитель, приверженный развитию экологически здоровых городов и пригодной для жизни планеты. В настоящее время он является издателем и редактором журнала The Art of the Green New Deal, журнала нового поколения о смене творческой культуры.
Промежуточный обмен
Агиланиу, Х., Дюрье, Дж., И Диллин, А. (2005). Метаболизм, синтез убихинона и долголетие.Genes Dev. 19 , 2399–2406. Абстрактный Статья
Андерсон, Г.Л. (1978). Ответы Caenorhabditis elegans (Nematoda: Rhabdiditae) на термический стресс и кислородное голодание. Жестяная банка. J. Zool. 56 , 1786–1791. Статья
Андерсон, Г.Л., Дузенбери Д. (1977). Критическое давление кислорода Caenorhabditis elegans . J. Nematol. 9 , 253–254.
Антеби, А. (2007). Генетика старения Caenorhabditis elegans . PLoS Genetics 3 , e129, 1565–1571. Абстрактный Статья
Апфельд, Дж., О’Коннор, Г., Макдонаг, Т., ДиСтефано, П.С., и Кертис, Р. (2004). AMP-активированная протеинкиназа aak-2 связывает уровни энергии и инсулиноподобные сигналы с продолжительностью жизни у C. elegans . Genes Dev. 18 , 3004–3009. Абстрактный Статья
Аткинсон, Х.Дж. (1980). Дыхание у нематод. У нематод как биологических моделей (т.2), Б. Цукерманн, изд. (Нью-Йорк: Academic Press), стр. 101–142.
Ауэрон Ф. и Ротштейн М. (1974). Биохимия нематод — XIII. Пероксисомы в свободноживущей нематоде, Turbatrix aceti . Комп. Биохим. Physiol. 49B , 261–271. Абстрактный
Бекман, К.Б., Эймс Б.Н. (1998). Созревает свободнорадикальная теория старения. Physiol. Ред. 78 , 547–581. Абстрактный
Бирнби Д.А., Линк Е.М., Гласные, Дж. Дж., Тиан, Х., Колакурчио, П. Л., Томас, Дж. Х. (2000). Трансмембранная гуанилилциклаза (DAF-11) и hsp90 (DAF-21) регулируют общий набор хемосенсорных поведений у Caenorhabditis elegans .Генетика 155 , 85–104. Абстрактный
Бишоп, Н.А. и Гуаренте, Л. (2007). Два нейрона опосредуют индуцированное ограничением диеты долголетие у C. elegans . Природа 447 , 545–550. Абстрактный Статья
Блэкстер, М.(1998). Caenorhabditis elegans — нематода. Наука 282 , 2041–2046. Абстрактный Статья
Болановски М.А., Рассел Р.Л., Якобсон Л.А. (1981). Количественные показатели старения нематоды Caenorhabditis elegans . I. Популяционные и продольные исследования двух поведенческих параметров. Мех. Aging Dev. 15 , 279–295.Абстрактный Статья
Болла, Р. (1980). Энергетический обмен нематод. В книге «Нематоды как биологические модели» (том 2) Б.М. Цукерманн, изд. (Нью-Йорк: Academic Press), стр. 165–192.
Бракман, Б.П., Хаутхуф, К., Де Вриз, А., и Ванфлетерен, Дж. Р. (1999). Очевидное разъединение производства энергии и потребление долгоживущими мутантами Clk Caenorhabditis elegans . Curr. Биол. 9 , 493–496. Абстрактный Статья
Бракман, Б.П., Хаутхуф, К., Де Вриз, А., и Ванфлетерен, Дж. Р. (2002a). Анализ метаболической активности при старении Caenorhabditis elegans .Мех. Aging Dev. 123 , 105–119. Абстрактный Статья
Braeckman, B.P., Houthoofd, K., and Vanfleteren, J.R. (2002b). Оценка метаболической активности при старении Caenorhabditis elegans : концепции и противоречия. Ячейка старения 1 , 82–88. Абстрактный Статья
Бракман, Б.П., Хоутхуфд, К., Брис, К., Ленертс, И., Де Вриз, А., Ван Эйген, С., Раес, Х., и Ванфлетерен, Дж. Р. (2002c). У мутантов Clk не наблюдается снижения энергетического обмена. Мех. Aging Dev. 123 , 1447–1456. Абстрактный Статья
Бракман Б.П., Хаутофд К. и Ванфлетерен Дж.Р. (2003). Энергетический обмен, антиоксидантная защита и старение у Caenorhabditis elegans .В журнале Topics in Current Genetics (Vol. 3) Model systems in Aging, T. Nyström и H.D. Осевач, ред. (Берлин: Springer-Verlag), С. 99–144.
Бракман Б.П., Ванфлетерен Дж.Р. (2007). Генетический контроль долголетия у C. elegans . . Exp. Геронтол. 42 , 90–98. Абстрактный Статья
Бранд, М.Д. (2000). Разъединение, чтобы выжить? Роль митохондриальной неэффективности в старении. Exp. Геронтол. 35 , 811–820. Абстрактный Статья
Браницки Р., Шибата Ю., Фенг Дж. И Хекими С. (2001). Фенотипический и супрессорный анализ дефекации у мутантов clk-1 показывает, что реакция на изменения температуры является активным процессом у Caenorhabditis elegans .Генетика 159 , 997–1006. Абстрактный
Бреннер, С. (1974). Генетика Caenorhabditis elegans . Генетика 77 , 71–94. Абстрактный
Брис, К., Ванфлетерен, Дж. Р., Бракман, Б. П. (2007). Проверка теории старения, основанной на скорости жизни / окислительном повреждении нематода модель Caenorhabditis elegans .Exp. Геронтол. 42 , 845–851. Абстрактный Статья
Будовская Ю.В., Ву К., Саутворт Л.К., Цзян М., Тедеско П., Джонсон Т.Е. и Ким С.К. (2008). Цепь транскрипции GATA elt-3 / elt-5 / elt-6 направляет старение в C. elegans . Ячейка 134 , 291-303. Абстрактный Статья
Бюхер, Э.Дж., Хансен, Э.Л., Ярвуд, Э.А. (1966). Активация фиколла белка, необходимого для созревания свободноживущих нематода. Proc. Soc. Exp. Биол. Med. 121 , 390–393. Абстрактный
Бернелл, А.М., Хаутхуф, К., О’Хэнлон, К., и Ванфлетерен, Дж. Р. (2005). Альтернативный метаболизм во время более поздней стадии нематода Caenorhabditis elegans .Exp. Геронтол. 40 , 850–856. Абстрактный Статья
Читвуд, Д.Дж., Датки, В.Р. (1991). Метаболизм растительных стеролов нематодами. Липиды 26 , 619–627. Абстрактный Статья
Коул, Р.Дж., Дутки С. (1969). Требование стерола в Turbatrix aceti и Panagrellus redivivus . J. Nematol. 1 , 72–75.
Колонна, У.Дж., и Макфадден, Б. (1975). Изоцитратлиаза паразитических и свободноживущих нематод. Arch. Биохим. Биофиз. 170 , 608–619. Абстрактный Статья
Купер, А.Ф. младший и Ван Ганди, С. Д. (1970). Метаболизм гликогена и нейтральных липидов у Aphelenchus avenae и Caenorhabditis sp. в аэробных, микроаэробных и анаэробных средах. J. Nematol. 2 , 305–315.
Купер, А.Ф. мл. И Ван Ганди, С.Д. (1971). Производство и использование этанола Aphelenchus avenae и Caenorhabditis sp.J. Nematol. 3 , 205–214.
Де Кайпер, К., и Ванфлетерен, Дж. Р. (1982). Потребление кислорода при развитии и старении нематоды Caenorhabditis elegans . Комп. Биохим. Physiol. 73A , 283–289. Статья
Диллин А., Сюй А.L., Arantes-Oliveira, N., Lehrer-Graiwer, J., Hsin, H., Fraser, A.G., Kamath, R.S., Ahringer, J., and Kenyon, С. (2002). Скорость поведения и старения определяется функцией митохондрий во время развития. Наука 298 , 2398–2401. Абстрактный Статья
Догерти, E.C., Хансен, E.L., Николас, W.L., Mollett, J.A., and Yarwood, E.А. (1959). Аксеническое культивирование Caenorhabditis briggsae (Nematoda: Rhabditidae) на средах определенного химического состава без добавок и с добавками. Анна. N.Y. Acad. Sci. 77 , 176–217. Статья
Догерти, E.C., и Хансен, E.L. (1957). Потребность в фолиевой кислоте и ее антагонизм со стороны аминоптерина у нематоды Caenorhabditis briggsae (Rhabditidae).Анат. Рек. 128 , 541–542.
Духон, С.А., Джонсон, Т. (1995). Движение как показатель жизнеспособности: сравнение дикого типа и мутанта age-1 Caenorhabditis elegans . J. Gerontol. Биол. Sci. Med. Sci. 50 , B254–261. Абстрактный
Дузенбери, Д.Б., Андерсон Г.Л., Андерсон Э.А. (1978). Термическая акклиматизация более обширна по поведенческим параметрам, чем по потреблению кислорода нематодой Caenorhabditis elegans . J. Exp. Zool. 206 , 191–198. Статья
Эпштейн, Дж., Химмельхох, С., и Гершон, Д. (1972). Исследования старения нематод. III. Электронно-микроскопические исследования на возрастное клеточное повреждение.Мех. Aging Dev. 1 , 245–255.
Юбанк, Дж. Дж., Барнс, Т. М., Лаковски, Б., Люсье, М., Бусси, Х., и Хекими, С. (1997). Структурно-функциональная консервация из Caenorhabditis elegans гена синхронизации clk-1 . Наука 275 , 980–983. Абстрактный Статья
Фабиан, Т.Дж., Джонсон Т. (1995). Общее содержание РНК, рРНК и поли (А) + РНК во время старения у Caenorhabditis elegans . Мех. Aging Dev. 83 , 155–170. Абстрактный Статья
Фей, Й.Дж., Лю, Дж. К., Иноуэ, К., Чжуан, Л., Мияке, Л., Мияучи, С., и Ганапати, В. (2004). Актуальность NAC -2 , переносчика цитрата, связанного с Na + , для продолжительности жизни, размера тела и содержания жира в Caenorhabditis elegans .Биохимический журнал 379 , 191–198. Абстрактный Статья
Фелкай, С., Юбанк, Дж. Дж., Лемье, Дж., Лаббе, Ж.-К., Браун, Г. Г., и Хекими, С. (1999). CLK-1 контролирует дыхание, поведение и старение нематоды Caenorhabditis elegans . EMBO J. 18 , 1783–1792. Абстрактный Статья
Фэн, Дж., Бюссьер Ф. и Хекими С. (2001). Транспорт электронов в митохондриях является ключевым фактором продолжительности жизни у Caenorhabditis elegans . Dev. Ячейка 1 , 633–644. Абстрактный Статья
Фёлль Р.Л., Плейерс А., Левандовски Г.Дж., Вермтер К., Хегеманн В. и Пауль Р.Дж. (1999). Анаэробиоз при нематоде Caenorhabditis elegans .Комп. Биохим. Physiol. 124B , 269–280. Абстрактный
Джемс, Д., Макэлви, Дж. Дж. (2005). Детоксикация широкого спектра действия: основной процесс обеспечения долголетия, регулируемый инсулином / IGF-1 сигнализация? Мех. Aging Dev. 126 , 381–387. Абстрактный Статья
Гнайгер, Э., Мендес, Г., Хэнд, С.С. (2000). Высокая эффективность фосфорилирования и угнетение несвязанного дыхания у митохондрии при гипоксии. Proc. Natl. Акад. Sci. USA 97 , 11080–11085. Абстрактный Статья
Грир, Э.Л., Довлатшахи, Д., Банко, М.Р., Виллен, Дж., Хоанг, К., Бланшар, Д., Гайги С.П., Брюне, А. (2007).AMPK-FOXO Путь опосредует долголетие, индуцированное новым методом ограничения диеты у C. elegans . Curr. Биол. 17 , 1646–1656. Абстрактный Статья
Голден, Дж. У., и Риддл, Д. Л. (1984а). Личинка дауэра Caenorhabditis elegans : влияние феромона, пищи и температуры на развитие.Dev. Биол. 102 , 368–378. Абстрактный Статья
Голден, Дж. У., и Риддл, Д. Л. (1984b). Индуцированный феромоном переключатель развития у Caenorhabditis elegans : чувствительные к температуре мутанты обнаруживают температурно-зависимый процесс дикого типа. Proc. Natl. Акад. Sci. USA 81 , 819–823. Абстрактный Статья
Град, Л.И., Лемир Б. (2004). Мутации митохондриального комплекса I в Caenorhabditis elegans вызывают дефицит цитохрома c оксидазы, окислительный стресс и витаминно-зависимый лактоацидоз. Гм. Мол. Genet. 13 , 303–314. Абстрактный Статья
Град Л.И., Сэйлз Л.С., Лемир Б.Д. (2005). Введение в лечение дополнительного пути окисления лактата. лактоацидоза и митохондриальной дисфункции у Caenorhabditis elegans .Proc. Natl. Акад. Sci. USA 102 , 18367–18372. Абстрактный Статья
Хаджери В.А., Трехо Дж. И Падилья, Пенсильвания. (2005). Характеристика субъядерных изменений у эмбрионов Caenorhabditis elegans , подвергшихся кратковременной, промежуточной и длительной остановке клеточного цикла, вызванной аноксией. BMC Cell Biol. 6 , 47. Абстрактный Статья
Халашек-Винер, Дж., Хаттра, Дж.С., Маккей, С., Пузырев, А., Стотт, Дж. М., Янг, Г.С., Холт, Р.А., Джонс, С.Дж.М., Марра, М.А., Брукс-Уилсон, A.R., et al. (2005). Анализ долгоживущих мутантов C. elegans daf-2 с использованием серийного анализа экспрессии генов. Genome Res. 15 , 603–615. Абстрактный Статья
Холливелл, Б., и Гаттеридж, J.M.C. (1999). Свободные радикалы в биологии и медицине (Oxford: Oxford University Press).
Ганновер, Дж. А., Форсайт, М. Е., Хеннесси, П. Т., Бродиган, Т. М., Лав, Д. К., Эшвелл, Г., и Краузе, М. (2005). Модель инсулинорезистентности Caenorhabditis elegans : измененное хранение макронутриентов и образование дауэра при нокауте OGT-1. Proc. Natl. Акад. Sci. USA 102 , 11266–11271.Абстрактный Статья
Хансен М., Чандра А., Митик Л.Л., Онкен Б., Дрисколл М. и Кеньон К. (2008). Роль аутофагии в расширении продолжительности жизни путем ограничения питания у C. elegans . PLoS Genetics 4 , e24. Абстрактный Статья
Хансен, М., Сюй, А.Л., Диллин, А., Кеньон, К. (2005). Новые гены, связанные с эндокринной, метаболической и диетической регуляцией продолжительность жизни по данным скрининга геномной РНКи Caenorhabditis elegans . PLoS Genetics 1 , 119–128. Абстрактный Статья
Хартман П.С., Исии Н., Кайзер Э., Морган П.Г., Седенский М.М. (2001). Митохондриальные мутации по-разному влияют на старение, изменчивость и чувствительность к анестетикам у Caenorhabditis elegans .Мех. Aging Dev. 122 , 1187–1201. Абстрактный Статья
Hedges, S.B. (2002). Происхождение и эволюция модельных организмов. Обзоры природы 3 , 838–849. Абстрактный Статья
Хеджес, С.Б. и Кумар, С. (2003). Геномные часы и эволюционные шкалы времени. Тенденции в генетике 19 , 200–206. Абстрактный Статья
Херндон, Л.А., Шмайсснер, П.Дж., Дударонек, Дж. М., Браун, П. А., Листнер, К. М., Сакано, Ю., Паупар, М. К., Холл, Д. Х. и Дрисколл, М. (2002). Стохастические и генетические факторы влияют на тканеспецифическое снижение старения C.elegans . Природа 419 , 808–814. Абстрактный Статья
Хиб В.Ф. и Ротштейн М. (1968). Потребность в стероле для размножения свободноживущей нематоды. Наука 160 , 778–779. Абстрактный
Хиб, В.Ф., Stockstad, E.L.R., и Rothstein, M. (1970). Необходимость гема для воспроизводства свободноживущей нематоды. Наука 168 , 143–144. Абстрактный
Холт, С.Дж., Риддл, Д.Л. (2003). SAGE изучает C. elegans углеводный обмен: доказательства анаэробного сдвига у долгоживущей личинки дауэра. Мех. Aging Dev. 124 , 779–800.Абстрактный Статья
Хонда, Ю., и Хонда, С. (1999). Генная сеть daf-2 , отвечающая за долголетие, регулирует устойчивость к окислительному стрессу и экспрессию гена Mn-супероксиддисмутазы у Caenorhabditis elegans . FASEB J. 13 , 1385–1393. Абстрактный
Хугевиджс, Д., Гойенс, Э., Девильд, С., Виерстраете, А., Моенс, Л., Виноградов, С., и Ванфлетерен, Дж. Р. (2007). Широкое разнообразие в структуре и профилях экспрессии среди членов семейства глобиновых белков Caenorhabditis elegans . BMC Genomics 8 , 356 Абстрактный Статья
Houthoofd, K., Braeckman, B.P., Johnson, T.Э., Ванфлетерен Дж. Р. (2003). Продление жизни с помощью диетических ограничений не зависит сигнального пути Ins / IGF-1 в Caenorhabditis elegans . Exp. Геронтол. 38 , 947–954. Абстрактный Статья
Houthoofd, K., Braeckman, B.P., Lenaerts, I., Brys, K., De Vreese, A., Van Eygen, S., и Vanfleteren, J.Р. (2002a). Axenic Рост регулирует удельную скорость метаболизма, устойчивость к стрессу и увеличивает продолжительность жизни у Caenorhabditis elegans . Exp. Геронтол. 37 , 1371–1378. Абстрактный Статья
Хаутхуфд, К., Бракман, Б.П., Ленертс, И., Брис, К., Де Вриз, А., Ван Эйген, С., и Ванфлетерен, Дж. Р. (2002b).Старение происходит обратное, и метаболизм возвращается к молодому уровню у выздоравливающих личинок дауэра C. elegans . Exp. Геронтол. 37 , 1015–21. Абстрактный Статья
Хаутхуфд, К., Бракман, Б.П., Ленертс, И., Брис, К., Де Вриз, А., Ван Эйген, С., и Ванфлетерен, Дж. Р. (2002c). Без снижения скорости метаболизма в пище ограничено Caenorhabditis elegans .Exp. Геронтол. 37 , 1359–1369. Абстрактный Статья
Хаутхуфд К., Бракман Б.П., Ленертс И., Брис К., Маттейссенс Ф., Де Вриз А., Ван Эйген С. и Ванфлетерен Дж. (2005). Мутации пути DAF-2 и ограничение пищи при старении Caenorhabditis elegans по-разному влияют на метаболизм. Neurobiol.Старение 26 , 689–696. Абстрактный Статья
Хаутхофд К., Фидальго М.А., Хугевиджс Д., Бракман Б.П., Ленертс И., Брис К., Маттейссенс Ф., Де Вризе А., Ван Эйген, С., Муньос, М.Дж., и Ванфлетерен, Дж. Р. (2005). Метаболизм, физиология и защита от стресса у трех стареющих мутантов Ins / IGF-1 нематоды Caenorhabditis elegans .Ячейка старения 4 , 87–95. Абстрактный Статья
Houthoofd, K., Gems, D., Johnson, T.E., and Vanfleteren, J.R. (2007). Ограничение питания нематоды Caenorhabditis elegans . Междисциплинарный. Верхний. Геронтол. 35 , 98–114. Абстрактный Статья
Джонсон, Т.Э., и Маккаффри Г. (1985). Запрограммированное старение или катастрофа из-за ошибок? Исследование двумерным полиакриламидом гель-электрофорез. Мех. Aging Dev. 30 , 285–297. Абстрактный Статья
Цзя, К., и Левин, Б. (2007). Аутофагия необходима для опосредованного диетическим ограничением продления жизни у C. elegans . elegans .Аутофагия 3 , 597–599. Абстрактный
Йонассен, Т., Дэвис, Д.Э., Ларсен, П.Л., Кларк, К.Ф. (2003). Репродуктивная пригодность и содержание хинонов у мутантов Caenorhabditis elegans clk-1 , получавших изоформы кофермента Q различной длины. J. Biol. Chem. 278 , 51735–51742. Абстрактный Статья
Джонс, С.Дж., Риддл, Д.Л., Пузырев, А.Т., Велкулеску, В.Е., Хиллиер, Л., Эдди, С.Р., Стриклин, С.Л., Бейли, Д.Л., Уотерстон, Р., и Марра, М.А. (2001). Изменения в экспрессии генов, связанные с задержкой развития и продолжительностью жизни у Caenorhabditis elegans . Genome Res. 11 , 1346–1352. Абстрактный Статья
Кайзер, Э.Б., Седенский М.М., Морган П.Г., Хоппель К. (2004). Окислительное фосфорилирование митохондрий нарушено в долгоживущем мутанте clk-1 . J. Biol. Chem. 279 , 54479–54486. Абстрактный Статья
Кеньон, К. (2005). Пластичность старения: идеи долгоживущих мутантов. Ячейка 120 , 449–460.Абстрактный Статья
Хан, Ф.Р., и Макфадден, Б.А. (1982). Caenorhabditis elegans : распад изоцитратлиазы во время личиночного развития. Exp. Паразитол. 54 , 47–54. Абстрактный Статья
Класс, М., и Хирш, Д. (1976). Нестареющий вариант развития Caenorhabditis elegans . Природа 260 , 523–525. Абстрактный Статья
Класс, М.Р. (1977). Старение нематоды Caenorhabditis elegans : основные биологические факторы и факторы окружающей среды, влияющие на продолжительность жизни. Мех. Aging Dev. 6 , 413–429. Абстрактный Статья
Кондо, М., Сено-Мацуда, Н., Янасэ, С., Исии, Т., Хартман, П.С., и Исии, Н. (2005). Влияние окислительного стресса на транслокацию DAF-16 у чувствительных к кислороду мутантов, mev-1 и gas-1 из Caenorhabditis elegans . Мех. Aging Dev. 126 , 637–641. Абстрактный Статья
Коршунов, С.С., Скулачев В.П., Старков А.А. (1997). Высокий протонный потенциал приводит в действие механизм производства активные формы кислорода в митохондриях. Письма FEBS 416 , 15–18. Абстрактный Статья
Кюнл, Дж., Бобик, Т., Проктер, Дж. Б., Бурмейстер, К., Хеппнер, Дж., Уайльд, И., Люерсен, К., Торда, А. Э., Вальтер, Р. Д., и Либау, Э.(2005). Функциональный анализ метилмалонил-КоА эпимеразы из Caenorhabditis elegans . FEBS J. 272 , 1465–1477. Абстрактный Статья
Лаковски Б. и Хекими С. (1998). Генетика ограничения калорийности у Caenorhabditis elegans . Proc. Natl. Акад. Sci. USA 95 , 13091–13096. Абстрактный Статья
Ламберт, А.Дж. И Бранд, доктор медицины (2007). Исследования митохондрий и старения, 2006–2007 гг. Ячейка старения 6 , 415–593 Абстрактный Статья
Ламитина, С.Т., Стрэндж, К. (2005). Транскрипционные мишени сигнального пути инсулина DAF-16 защищают C. elegans от экстремального гипертонического стресса. Являюсь. J. Physiol. Cell Physiol. 288 , C467-C474.Абстрактный Статья
Ламитина, С.Т., Моррисон, Р., Моекель, Г.М., и Стрэндж, К. (2004). Адаптация нематоды Caenorhabditis elegans к экстремальному осмотическому стрессу. Являюсь. J. Physiol. Cell Physiol. 286 , C785-C791. Абстрактный Статья
Ламитина, С.Т., Хуанг, К.Г., Стрэндж, К. (2006). Полногеномный скрининг РНКи определяет повреждение белка как регулятор экспрессия осмозащитных генов. Proc. Natl. Акад. Sci. USA 103 , 12173–12178. Абстрактный Статья
Ларсен, П. (1993). Старение и устойчивость к окислительному повреждению у Caenorhabditis elegans . Proc.Natl. Акад. Sci. USA 90 , 8905–8909. Абстрактный Статья
Ли, Д.Л., и Аткинсон, Х.Дж. (1976). Физиология нематод . 2 nd edition (Нью-Йорк: MacMillan).
Ли, E.Y., Шим, Y.H., Читвуд, Д.Дж., Хван, С.Б., Ли, Дж., Пайк, Ю.К. (2005). Трансгенный ген Caenorhabditis elegans , продуцирующий холестерин, живет дольше из-за недавно приобретенной повышенной стрессоустойчивости. Биохим. Биофиз. Res. Commun. 328 , 929–936. Абстрактный Статья
Ли С.С., Ли Р.Й.Н., Фрейзер А.Г., Камат Р.С., Аринджер Дж. И Рувкун Г. (2003). Систематический скрининг РНКи определяет критическая роль митохондрий в C.elegans долговечность. Nature Genetics 33 , 40–48. Абстрактный Статья
Лю А. и Ротштейн М. (1976). Биохимия нематод — XV. Изменения ферментов, связанные с экскрецией глицерина у Caenorhabditis briggsae . Комп. Биохим. Physiol. 54B , 233–238. Абстрактный
Лю, Ф., Тэтчер, Дж. Д., Баррал, Дж. М., и Эпштейн, Х. Ф. (1995). Бифункциональный белок глиоксилатного цикла Caenorhabditis elegans : белок кишечника и мышц, регулируемый развитием. Dev. Биол. 169 , 399–414. Абстрактный Статья
Лю Ф., Тэтчер Дж. Д. и Эпштейн Х. Ф. (1997). Индукция экспрессии глиоксилатного цикла у Caenorhabditis elegans : реакция голодания на протяжении личиночного развития.Биохимия 36 , 255–260. Абстрактный Статья
Лу, Н. К., Гетч, К. М. (1993). Потребность в углеводах Caenorhabditis elegans и окончательная разработка химически определенной среды. Nematologica 39 , 303–311.
Лу, Н.C., Hieb, W.F., Stokstad, E.L.R. (1974). Накопление формимино-L-глутаминовой кислоты у свободноживущей нематоды Caenorhabditis briggsae в связи с дефицитом фолиевой кислоты. Proc. Soc. Exp. Биол. Med. 145 , 67–69. Абстрактный
Лу, Н.С., Хиб, В.Ф., и Стокстад, Е.Л.Р. (1976). Влияние витамина B 12 и фолиевой кислоты на биосинтез метионина из гомоцистеина у нематоды Caenorhabditis briggsae .Proc. Soc. Exp. Биол. Med. 151 , 701–706. Абстрактный
Лу, Н.С., Хугенберг Г. мл., Бриггс, Г.М., и Стокстад, Э.Л.Р. (1978). Стимулирующая рост активность нескольких липид-зависимых соединений в свободноживущей нематоде Caenorhabditis briggsae . Proc. Soc. Exp. Биол. Med. 158 , 187–191. Абстрактный
Лу, Н.К., Ньютон, К., Стокстад, E.L.R. (1977). Потребность в стеринах и различных предшественниках стеролов у свободноживущих нематоды. Nematologica 23 , 57–60.
Лунд, Дж., Тедеско, П., Дюк, К., Ван, Дж., Ким, С.К., и Джонсон, Т. (2002). Транскрипционный профиль старения C. elegans . Curr. Биол. 12 , 1566–1573. Абстрактный Статья
Сеть, P.Э., и МакГи, Дж. Д. (1999). Биохимия C. elegans . В C. elegans — практический подход, И.А. Надежда, изд. (Оксфорд, Великобритания: Oxford University Press), стр. 227–244.
Мэтьюз, К.К., и Ван Холд К.Э. (1996). Биохимия, второе издание (Менло-Парк, Калифорния: издательство «Бенджамин / Каммингс»).
Макэлви, Дж., Бубб К. и Томас Дж. Х. (2003). Выходы транскрипции вилкообразного белка DAF-16 Caenorhabditis elegans . Ячейка старения 2 , 111–121. Абстрактный Статья
Макэлви, Дж. Дж., Шустер, Э., Блан, Э., Томас, Дж. Х. и Джемс, Д. (2004). Общая транскрипционная подпись у личинок Caenorhabditis elegans, dauer и долгоживущих мутантов daf-2 предполагает участие системы детоксикации в обеспечении долголетия.J. Biol. Chem. 279 , 44533–44543. Абстрактный Статья
Макэлви, Дж. Дж., Шустер, Э., Блан, Э., Торнтон, Дж., И Джемс, Д. (2006). Подтверждены метаболические особенности, связанные с диапаузой у долгоживущих мутантов daf-2 у нематоды Caenorhabditis elegans . Мех. Aging Dev. 127 , 458–472.Абстрактный Статья
Мак-Кинли, М.П., Трелиз, Р.М. (1978a). Сосуществование изоцитратлиазы и НАДФ-изоцитратдегидрогеназы в митохондриях Turbatrix aceti . Биохим. Биофиз. Res. Comm. 81 , 434–438. Абстрактный Статья
Мак-Кинли, М.П., Трелиз Р. (1978b). Ферменты глиоксилатного цикла и каталаза в митохондриях, фракционированных дигитонином, в Turbatrix aceti . Протоплазма 94 , 249–261. Статья
Менденхолл А.Р., ЛаРю Б. и Падилья П.А. (2006). Глицеральдегид-3-фосфатдегидрогеназа опосредует аноксический ответ и выживаемость в Caenorhabditis elegans .Генетика 174 , 1173–1187. Абстрактный Статья
Миядера, Х., Амино, Х., Хираиши, А., Така, Х., Мураяма, К., Миёси, Х., Сакамото, К., Исии, Н., Хекими, С., и Кита, К. (2001). Изменен биосинтез хинона у долгоживущих мутантов clk-1 Caenorhabditis elegans . J. Biol. Chem. 276 , 7713–7716.Абстрактный Статья
Мухопадхьяй, А., О, С.В., и Тиссенбаум, Х.А. (2006). Пути заражения к DAF-16 / FOXO и от него. Exp. Геронтол. 41 , 928–934. Абстрактный Статья
Мерфитт, Р.Р., Фогель, К., и Санади, Д. (1976). Характеристика митохондрий свободноживущей нематоды, Caenorhabditis elegans . Комп. Биохим. Physiol. 53B , 423–430. Абстрактный Статья
Николас, У.Л., Догерти, Э.С., Хансен, Э.Л., Холм-Хансен, О., и Моисей, В. (1960). Включение 14 C из ацетата натрия-2- 14 C в аминокислоты обитающей в почве нематоды Caenorhabditis briggsae .J. Exp. Биол. 37 , 435–443.
Николас У.Л., Хансен Э. и Догерти Э. К. (1962). Витамины группы В, необходимые для Caenorhabditis briggsae (Rhabditidae). Nematologica 8 , 129–135.
Николас, W.L., и Jantunen, R. (1963). Потребность в биотине для Caenorhabditis briggsae (Rhabditidae).Nematologica 9 , 332–336.
Николлс, Д. (2002). Функция и дисфункция митохондрий в клетке: ее значение для старения и связанных со старением заболеваний. Int. J. Biochem. Cell Biol. 34 , 1372–1381. Абстрактный Статья
Нистуль, Т.Г., Рот, М. (2004). Прерывистая анимация, вызванная угарным газом, защищает от гипоксического повреждения у Caenorhabditis elegans . Proc. Natl. Акад. Sci. USA 101 , 9133–9136. Абстрактный Статья
О’Риордан, В.Б., Бернелл, А.М. (1989). Промежуточный метаболизм у личинки дауэра нематоды Caenorhabditis elegans — 1.Гликолиз, глюконеогенез, окислительное фосфорилирование и цикл трикарбоновых кислот. Комп. Биохим. Physiol. 92B , 233–238. Статья
О’Риордан, В.Б., Бернелл, А.М. (1990). Промежуточный метаболизм у личинки дауэра нематоды Caenorhabditis elegans — II. Глиоксилатный цикл и окисление жирных кислот. Комп. Биохим. Physiol. 95 В, 125–130.
Огг, С., Паради, С., Готтлиб, С., Паттерсон, Г.И., Ли, Л., Тиссенбаум, Х.А., и Рувкун, Г. (1997). Транскрипция головы вилки Фактор DAF-16 трансдуцирует инсулиноподобные сигналы метаболизма и долголетия у C. elegans . Природа 389 , 994–999. Абстрактный Статья
Падилья, П.А., Нистул, Т.Г., Загер, Р.А., Джонсон, А.С., и Рот, М.Б. (2002). Дефосфорилирование регулируемого клеточного цикла белков коррелирует с вызванной аноксией приостановкой анимации у Caenorhabditis elegans . Мол. Биол. Ячейка 13 , 1473–1483. Абстрактный Статья
Панагидес, Дж., И Ротштейн, М. (1973). Синтез рибита в свободноживущей нематоде, Turbatrix aceti .II. Биосинтетические пути. Int. J. Biochem. 4 , 407–414. Статья
Пановски, С.Х., Вольф, С., Агиланиу, Х., Дюрье, Дж., И Диллин, А. (2007). PHA-4 / Foxa обеспечивает продолжительность жизни C. elegans, вызванную ограничением диеты. Природа 447 , 536–537. Абстрактный Статья
Патель, Т.Р., Макфадден Б.А. (1977). Частицы изоцитратлиазы и малатсинтазы в Caenorhabditis elegans . Arch. Биохим. Биофиз. 183 , 24–30. Абстрактный Статья
Пауль Р.Дж., Гохла Дж., Фёлль Р. и Шнекенбургер Х. (2000). Метаболические адаптации к изменениям окружающей среды у Caenorhabditis elegans . Комп. Биохим.Physiol. 127B , 469–479. Абстрактный
Промислоу Д.Э.Л. и Хазелкорн Т.С. (2002). Повозрастные показатели метаболизма и смертности представителей рода Drosophila . Ячейка старения 1 , 66–74. Абстрактный Статья
Рао, А.U., Carta, L.K., Lesuisse, E., and Hamza, I. (2005). Отсутствие синтеза гема у свободноживущих эукариот. Proc. Natl. Акад. Sci. USA 102 , 4270–4275. Абстрактный Статья
Rea, S.L. (2005). Метаболизм у мутантов Caenorhabditis elegans Mit. Exp. Геронтол. 40 , 841–849. Абстрактный Статья
Ротштейн, М.(1963). Биохимия нематод. III. Продукты экскреции. Комп. Биохим. Physiol. 9Б , 51–59. Абстрактный Статья
Ротштейн, М. (1965). Биохимия нематод. V. Промежуточный метаболизм и взаимопревращения аминокислот в Caenorhabditis briggsae . Комп. Биохим. Physiol. 14B , 541–552. Абстрактный Статья
Ротштейн, М.(1968). Биохимия нематод. IX. Отсутствие биосинтеза стеролов у свободноживущих нематод. Комп. Биохим. Physiol. 27B , 309–317. Абстрактный Статья
Ротштейн, М. (1969). Биохимия нематод. X. Экскреция глицерина у свободноживущих нематод. Комп. Биохим. Physiol. 30B , 641–648. Абстрактный Статья
Ротштейн, М.(1970). Биохимия нематод. XI. Биосинтез жирных кислот Caenorhabditis briggsae и Panagrellus redivivus . Int. J. Biochem. 1 , 422–428. Статья
Ротштейн, М., и Майо, Х. (1964a). Биохимия нематод. IV. Об изоцитратлиазе в Caenorhabditis briggsae . Arch. Биохим. Биофиз. 108 , 134–142.Абстрактный Статья
Ротштейн, М., и Майо, Х. (1964b). Синтез глицина и изоцитратлиаза у нематоды Caenorhabditis briggsae . Биохим. Биофиз. Res. Commun. 14 , 43–53. Абстрактный Статья
Ротштейн, М., и Мэйо, Х. (1965). Биохимия нематод. VII. Присутствие изоцитратлиазы в Panagrellus redivivus , Turbatrix aceti и Rhabditis anomala . Комп. Биохим. Physiol. 16 , 361–365. Абстрактный Статья
Ротштейн, М., и Майо, Х. (1966). Биохимия нематод. VIII. Малатсинтетаза. Комп. Биохим.Physiol. 17B , 1181–1188.
Ротштейн, М., Томлинсон, Г.А. (1961). Биосинтез аминокислот нематодой Caenorhabditis briggsae . Биохим. Биофиз. Acta 49 , 625–627. Абстрактный
Ротштейн, М., Томлинсон, Г. (1962).Биохимия нематод. II. Биосинтез аминокислот. Биохим. Биофиз. Acta 63 , 471–480. Абстрактный Статья
Рубин, Х., Трелиз, Р. (1976). Субклеточная локализация ферментов глиоксилатного цикла у личинок Ascaris suum . J. Cell Biol. 70 , 374–383. Абстрактный
Рузанов, П., Риддл, Д.Л., Марра, М.А., Маккей, С.Дж., Джонс, С.М. (2007). Гены, которые могут модулировать продолжительность жизни у C. elegans как у личинок dauer, так и у долгоживущих особей daf-2 . Exp. Геронтол. 42 , 825–839. Абстрактный Статья
Сейр, Ф.В., Хансен, Э.Л., Ярвуд, Э.А. (1963). Биохимические аспекты питания Caenorhabditis briggsae .Exp. Паразитол. 13 , 98–107. Абстрактный
Шульц, Т.Дж., Зарсе, К., Войт, А., Урбан, Н., Биррингер, М., и Ристоу, М. (2007). Ограничение глюкозы увеличивает продолжительность жизни Caenorhabditis elegans за счет стимулирования митохондриального дыхания и увеличения окислительного стресса. Cell Metab. 6 , 280–293. Абстрактный Статья
Скотт, Б.А., Авидан М.С., Краудер К. (2002). Регулирование гипоксической смерти у C. elegans с помощью DAF-2, гомолога рецепторов инсулина / IGF. Наука 296 , 2388–2391. Абстрактный Статья
Седенский М.М., Морган П.Г. (2006). Митохондриальное дыхание и активные формы кислорода в C. elegans . Exp. Геронтол. 41 , 957–967.Абстрактный Статья
Сено-Мацуда, Н., Ясуда, К., Цуда, М., Окубо, Т., Йошимура, С., Накадзава, Х., Хартман, П.С. и Исии, Н. (2001). А дефект в большой субъединице цитохрома b в комплексе II вызывает как избыточную продукцию супероксид-аниона, так и аномальный энергетический метаболизм в Caenorhabditis elegans .J. Biol. Chem. 276 , 41553–41558. Абстрактный Статья
Шен К., Пауэлл-Коффман Дж. А. (2003). Генетический анализ передачи сигналов гипоксии и ответа у C. elegans . . Анна. Акад. Sci. 995 , 191–199. Абстрактный
Шояма, Т., Одзаки, Т., Исии, Н., Йокота, С., и Суда, Х. (2007). Основной принцип продолжительности жизни нематоды C. elegans . Мех. Aging Dev. 128 , 529–539. Абстрактный Статья
Сохал, Р.С., и Вайндрух, Р. (1996). Окислительный стресс, ограничение калорийности и старение. Наука 273 , 59–63. Абстрактный
Штейн, Л.Д., Бао, З., Блазиар, Д., Блюменталь, Т., Брент, М.Р., Чен, Н., Чинвалла, А., Кларк, Л., Кли, К., Коглан, А., и другие. (2003). Последовательность генома Caenorhabditis briggsae : платформа для сравнительной геномики. PLoS Biology 1 , 166–192. Абстрактный Статья
Шевчик, Н.Дж., Козак, Э., и Конли, К.А. (2003). Химически определенная среда и Caenorhabditis elegans . BMC Biotechnology 3 , 19. Абстрактный Статья
Такамия, С., Мацуи, Т., Така, Х., Мураяма, К., Мацуда, М., и Аоки, Т. (1999). Свободноживущие нематоды Caenorhabditis elegans обладают в своих митохондриях дополнительным родохиноном, важным компонентом эукариотической системы фумаратредуктазы.Arch. Биохим. Биофиз. 371 , 284–9. Абстрактный Статья
Тауб, Дж., Лау, Дж. Ф., Ма, К., Хан, Дж. Х., Хок, Р., Ротблатт, Дж., И Чалфи, М. (1999). Необходима цитозольная каталаза для увеличения продолжительности взрослой жизни у мутантов C. elegans daf-C и clk-1 . Природа 399 , 162–166.Абстрактный Статья
Таден, Дж.Дж., и Шмуклер Рейс, Р.Дж. (2000). Аммиак, дыхание и продолжительность жизни нематод: сведения о регуляции метаболизма продолжительности жизни от временного масштабирования. Варенье. Старение доц. 23 , 75–84.
Тиленс, А.Г.М., Ротте, К., Ван Хеллемонд, Дж. Дж., И Мартин, В. (2002). Митохондрии, какими мы их не знаем. Тенденции Биохим. Sci. 27 , 564–572. Абстрактный
Толберт, Н. (1981). Метаболические пути в пероксисомах и глиоксисомах. Анна. Rev. Biochem. 50 , 133–157.
Цанг, В.Ю., Сэйлз, Л.С., Град, Л.И., Пилигрим, Д.Б., Лемир, Б.Д. (2001). Дефицит митохондриальной дыхательной цепи у Ceanorhabditis elegans приводит к задержке развития и увеличению продолжительности жизни. J. Biol. Chem. 276 , 32240–32246. Абстрактный Статья
Урета-Видал, А., Эттвиллер, Л., и Бирни, Э. (2003).Сравнительная геномика: полногеномный анализ у многоклеточных эукариот. Nature Reviews Genetics 4 , 251–262. Абстрактный Статья
Ван Вурхис, W.A. (2002). Влияние скорости метаболизма на продолжительность жизни нематоды Caenorhabditis elegans . Ячейка старения 1 , 91–101. Абстрактный
Ван Вурхис, В.А. и Уорд С. (1999). Генетические условия и условия окружающей среды, увеличивающие продолжительность жизни у Caenorhabditis elegans , снижают скорость метаболизма. Proc. Natl. Акад. Sci. USA 96 , 11399–11403. Абстрактный
Ван Вурхис, В.А., и Уорд, С. (2000). Широкая толерантность к кислороду у нематоды Caenorhabditis elegans . J. Exp. Биол. 203 , 2467–2478.Абстрактный
Ванфлетерен, Дж. Р. (1974). Фактор роста нематод. Природа 248 , 255–257. Абстрактный
Ванфлетерен, Дж. Р. (1980). Нематоды как модели питания. В книге «Нематоды как биологические модели» (том 2) Б.М. Цукерманн, изд. (Нью-Йорк: Academic Press), стр.47–79.
Ванфлетерен, Дж. Р. (1993). Окислительный стресс и старение у Caenorhabditis elegans . Биохим. J. 292 , 605–608. Абстрактный
Ванфлетерен, Дж. Р., и Де Вриз, А. (1996). Скорость аэробного метаболизма и потенциал производства супероксида у нематод Caenorhabditis elegans. J. Exp. Zool. 274 , 93–100. Абстрактный
Вастрик, И., Д’Эустахио, П., Шмидт, Э., Джоши-Топ, Г., Гопинат, Г., Крофт, Д., де Боно, Б., Гиллеспи, М., Джассал, Б., Lewis, S., et al. (2007). Reactome: база знаний о биологических путях и процессах. Genome Biol. 8 , R39. Абстрактный Статья
Веллаи, Т., Такач-Веллаи, К., Чжан, Ю., Ковач, А.Л., Орос, Л., и Мюллер, Ф. (2003). Генетика: влияние TOR-киназы по продолжительности жизни в C. elegans . Природа, 426 , 620. Абстрактный Статья
Вентура, Н., Ри, С.Л. (2007). Caenorhabditis elegans митохондриальных мутантов в качестве исследовательского инструмента для изучения нейродегенеративных заболеваний человека, связанных с митохондриальной дисфункцией.Biotechnol. J. 2 , 584–595. Абстрактный Статья
Вентура, Н., Ри, С.Л., Тести, Р. (2006). Долгоживущие митохондриальные мутанты C. elegans как модель митохондриально-ассоциированных заболеваний человека. Exp. Геронтол. 41 , 974–991. Абстрактный Статья
Уодсворт, У.Г., Риддл Д. (1989). Регуляция развития энергетического обмена у Caenorhabditis elegans . Dev. Биол. 132 , 167–173. Абстрактный
Уокер, Г., Хаутхуф, К., Ванфлетерен, Дж. Р., и Джемс, Д. (2005). Ограничение диеты в C. elegans : от эффектов скорости жизни до путей восприятия питательных веществ. Мех. Aging Dev. 126 , 929–937.Абстрактный Статья
Ван Дж., Ким С.К. (2003). Глобальный анализ экспрессии гена dauer у Caenorhabditis elegans . Разработка 130 , 1621–1634. Абстрактный
Уиллер, Дж. М., и Томас, Дж. Х. (2006).Идентификация нового семейства генов, участвующих в реакции на осмотический стресс у Caenorhabditis elegans . Генетика 174 , 1327–1336. Абстрактный Статья
Уиллет, Д.Д., Дауни, В.Л. (1974). Биосинтез стеролов у свободноживущей нематоды Panagrellus redivivus . Биохим. J. 138 , 233–237. Абстрактный
Вольф, С., и Диллин, А. (2006). Трифект старения в Caenorhabditis elegans . Exp. Геронтол. 41 , 894–903. Абстрактный Статья
Вонг, А., Бутис, П., и Хекими, С. (1995). Мутации в гене clk-1 Caenorhabditis elegans влияют на время развития и поведения. Генетика 139 , 1247–1259.Абстрактный
Райт, Д.Дж. (1975a). Удаление азотистых соединений с помощью Panagrellus redivivus , Goodey, 1945 (Nematoda: Cephalobidae). Комп. Биохим. Physiol. 52B , 247–253. Абстрактный
Райт, Д.Дж. (1975b). Исследования катаболизма азота у Panagrellus redivivus , Goodey, 1945 (Nematoda: Cephalobidae).Комп. Биохим. Physiol. 52B , 255–260. Абстрактный
Ю., Х., Ларсен, П. (2001). DAF-16-зависимые и независимые мишени экспрессии DAF-2 инсулино-рецепторного пути в Caenorhabditis elegans включают FKBP. J. Mol. Биол. 314 , 1017–1028. Абстрактный Статья
.